
The Effect of Supplementation Using a Mixture of Fish Oil and Linseed on the Level of Immunomodulatory Components in Bovine Colostrum

Abstract
:1. Introduction
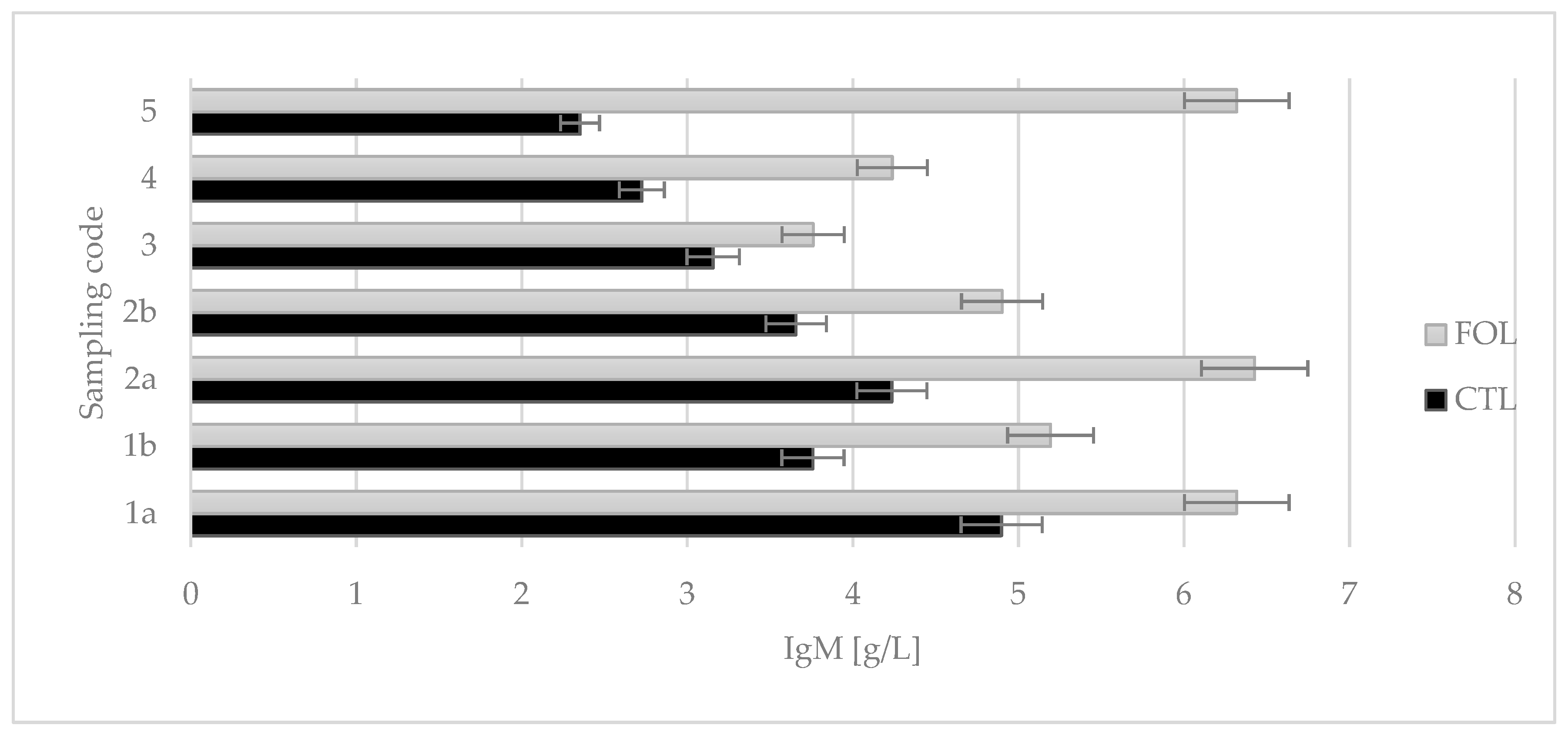
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Site and Study Animals
4.2. Chemical Analyses of Colostrum
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Błaszkowska, M.; Twardoń, J. Koncentracja IgG1, IgG2 i IgM w siarze krów oraz w surowicy cieląt pobranej w różnym czasie po urodzeniu. Med. Weter. 2005, 61, 1308–1311. [Google Scholar]
- Pejsak, Z.; Truszczyński, M. Uodparnianie noworodków i młodych zwierząt. Życie Wet 2007, 3, 183–186. [Google Scholar]
- McGrath, B.; Fox, P.; McSweeney, P.; Kelly, A. Composition and properties of bovine colostrum: A review. Dairy Sci. Technol. 2016, 96, 133–158. [Google Scholar] [CrossRef] [Green Version]
- Hammon, H.; Zanker, I.; Blum, J. Delayed colostrum feeding affects IGF-I and insulin plasma concentrations in neonatal calves. J. Dairy Sci. 2000, 83, 85–92. [Google Scholar] [CrossRef]
- Foley, J.; Otterby, D. Availability, storage, treatment, composition, and feeding value of surplus colostrum: A review. J. Dairy Sci. 1978, 61, 1033–1060. [Google Scholar] [CrossRef]
- Barrington, G.; McFadden, T.; Huyler, M.; Besser, T. Regulation of colostrogenesis in cattle. Livest. Prod. Sci. 2001, 70, 95–104. [Google Scholar] [CrossRef]
- Baumrucker, C.; Burkett, A.; Magliaro-Macrina, A.; Dechow, C. Colostrogenesis: Mass transfer of immunoglobulin G1 into colostrum. J. Dairy Sci. 2010, 93, 3031–3038. [Google Scholar] [CrossRef] [Green Version]
- Coleman, L.; Hickson, R.; Amoore, J.; Laven, R.; Back, P. Colostral immunoglobulin G as a predictor for serum immunoglobulin G concentration in dairy calves. Proc. New Zealand Soc. Anim. Prod. 2015, 75, 3–8. [Google Scholar]
- Puppel, K.; Gołębiewski, M.; Grodkowski, G.; Slósarz, J.; Kunowska-Slósarz, M.; Solarczyk, P.; Łukasiewicz, M.; Balcerak, M.; Przysucha, T. Composition and factors affecting quality of bovine colostrum: A review. Animals 2019, 9, 1070. [Google Scholar] [CrossRef] [Green Version]
- Siqueiros-Cendón, T.; Arévalo-Gallegos, S.; Iglesias-Figueroa, B.F.; García-Montoya, I.A.; Salazar-Martínez, J.; Rascón-Cruz, Q. Immunomodulatory effects of lactoferrin. Acta Pharmacol. Sin. 2014, 35, 557–566. [Google Scholar] [CrossRef]
- Król, J.; Litwińczuk, Z.; Brodziak, A.; Karasińska, A. Zawartość immunoglobulin G w mleku w zależności od rasy i wieku krów oraz fazy laktacji. Med. Weter. 2014, 70, 237–241. [Google Scholar]
- Larson, B.; Heary, H., Jr.; Devery, J. Immunoglobulin production and transport by the mammary gland. J. Dairy Sci. 1980, 63, 665–671. [Google Scholar] [CrossRef]
- Levieux, D.; Ollier, A. Bovine immunoglobulin G, β-lactoglobulin, α-lactalbumin and serum albumin in colostrum and milk during the early post partum period. J. Dairy Res. 1999, 66, 421–430. [Google Scholar] [CrossRef]
- Georgiev, I.P. Differences in chemical composition between cow colostrum and milk. Bulg. J. Vet. Med. 2008, 11, 3–12. [Google Scholar]
- Hernández-Ledesma, B.; Recio, I.; Amigo, L. β-Lactoglobulin as source of bioactive peptides. Amino Acids 2008, 35, 257–265. [Google Scholar] [CrossRef]
- Tripathi, V.; Vashishtha, B. Bioactive compounds of colostrum and its application. Food Rev. Int. 2006, 22, 225–244. [Google Scholar] [CrossRef]
- Gajda-Morszewski, P.; Śpiewak, K. Laktoferyna–Białko Multipotencjalne. In Zeszyty Naukowe Towarzystwa Doktorantów Uniwersytetu Jagiellońskiego; Nauki Ścisłe: Krakow, Poland, 2015; pp. 177–188. [Google Scholar]
- Ward, P.; Paz, E.; Conneely, O. Lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2540–2548. [Google Scholar] [CrossRef]
- Małaczewska, J.; Rotkiewicz, Z. Laktoferyna-białko multipotencjalne. Med. Weter. 2007, 63, 136–139. [Google Scholar]
- Aimutis, W.R. Bioactive properties of milk proteins with particular focus on anticariogenesis. J. Nutr. 2004, 134, 989S–995S. [Google Scholar]
- Andrzejczak, O. Lizozym–aktualne trendy w nauce i przemyśle. Przegląd Wybr. Pr. Z Zakr. Enzymol. 2016, 59–71. [Google Scholar]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Marciniak-Lukasiak, K. Rola i znaczenie kwasów tłuszczowych omega-3. Żywność Nauka Technol. Jakość 2011, 6, 24–35. [Google Scholar]
- Wojciech, K. Rola wielonienasyconych kwasów tłuszczowych omega-3 w przebiegu zapaleń. Med. Weter 2015, 71, 608–614. [Google Scholar]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Michaelsen, K.F.; Dewey, K.G.; Perez-Exposito, A.B.; Nurhasan, M.; Lauritzen, L.; Roos, N. Food sources and intake of n-6 and n-3 fatty acids in low-income countries with emphasis on infants, young children (6–24 months), and pregnant and lactating women. Matern. Child Nutr. 2011, 7, 124–140. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Banu, J.; Rahman, M.; Causey, J.; Fernandes, G. Biological effects of conjugated linoleic acids in health and disease. J. Nutr. Biochem. 2006, 17, 789–810. [Google Scholar] [CrossRef]
- Ma, A.; Koka, R.; Burkett, P. Diverse functions of IL-2, IL-15, and IL-7 in lymphoid homeostasis. Annu. Rev. Immunol. 2006, 24, 657–679. [Google Scholar] [CrossRef]
- Young, H.; Bream, J. IFN-γ: Recent advances in understanding regulation of expression, biological functions, and clinical applications. In Interferon: The 50th Anniversary; Springer: Berlin/Heidelberg, Germany, 2007; pp. 97–117. [Google Scholar]
- Ramírez-Santana, C.; Castellote, C.; Castell, M.; Moltó-Puigmartí, C.; Rivero, M.; Pérez-Cano, F.J.; Franch, À. Enhancement of antibody synthesis in rats by feeding cis-9, trans-11 conjugated linoleic acid during early life. J. Nutr. Biochem. 2011, 22, 495–501. [Google Scholar] [CrossRef]
- Hussen, J.; Dänicke, S.; Schuberth, H. The effect of a long term dietary supplementation with conjugated linoleic acid (CLA) on the composition of bovine peripheral blood mononuclear cells (PBMC) and the concentration of IgG isotypes in blood and milk. Proc. Soc. Nutr. Physiol. 2011, 20, 85. [Google Scholar]
- Calder, P.C. Polyunsaturated fatty acids, inflammation, and immunity. Lipids 2001, 36, 1007–1024. [Google Scholar] [CrossRef]
- Harizi, H.; Corcuff, J.-B.; Gualde, N. Arachidonic-acid-derived eicosanoids: Roles in biology and immunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Przybylska, J.; Albera, E.; Kankofer, M. Antioxidants in bovine colostrum. Reprod. Domest. 2007, 42, 402–409. [Google Scholar] [CrossRef]
- Guz, J.; Dziaman, T.; Szpila, A. Czy witaminy antyoksydacyjne mają wpływ na proces karcynogenezy? Do antioxidant vitamins influence carcinogenesis? Postepy Hig. Med. Dosw. 2007, 61, 185–198. [Google Scholar]
- Debier, C.; Pottier, J.; Goffe, C.; Larondelle, Y. Present knowledge and unexpected behaviours of vitamins A and E in colostrum and milk. Livest. Prod. Sci. 2005, 98, 135–147. [Google Scholar] [CrossRef]
- Puppel, K.; Gołębiewski, M.; Konopka, K.; Kunowska-Slósarz, M.; Slósarz, J.; Grodkowski, G.; Przysucha, T.; Balcerak, M.; Madras-Majewska, B.; Sakowski, T. Relationship between the quality of colostrum and the formation of microflora in the digestive tract of calves. Animals 2020, 10, 1293. [Google Scholar] [CrossRef]
- Quigley Iii, J.; Martin, K.; Dowlen, H.; Wallis, L.; Lamar, K. Immunoglobulin concentration, specific gravity, and nitrogen fractions of colostrum from Jersey cattle. J. Dairy Sci. 1994, 77, 264–269. [Google Scholar] [CrossRef]
- Sobczuk-Szul, M.; Wielgosz-Groth, Z.; Wronski, M.; Rzemieniewski, A. Changes in the bioactive protein concentrations in the bovine colostrum of Jersey and Polish Holstein–Friesian cows. Turk. J. Vet. Anim. Sci. 2013, 37, 43–49. [Google Scholar] [CrossRef]
- Bar, E.; Tiris, I.; Sarbu, D.; Iridon, C.; Ochea, I.; Bratu, I. Full characterization of bovine colostrum, raw material for dietary supplements. His beneficial effect on the human immune system. AUCFT 2010, 14, 33–40. [Google Scholar]
- Król, J.; Litwińczuk, Z.; Brodziak, A.; Barłowska, J. Lactoferrin, lysozyme and immunoglobulin G content in milk of four breeds of cows managed under intensive production system. Pol. J. Vet. Sci. 2010, 13, 357–361. [Google Scholar]
- Gulliksen, S.; Lie, K.; Sølverød, L.; Østerås, O. Risk factors associated with colostrum quality in Norwegian dairy cows. J. Dairy Sci. 2008, 91, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Abdullahoğlu, E.; Duru, S.; Özlüer, A.; Filya, İ. Factors affecting colostrum quality and calf passive transfer levels in Holstein cattle. Anim. Sci. Pap. Rep. 2019, 37, 29–39. [Google Scholar]
- Dunn, A.; Ashfield, A.; Earley, B.; Welsh, M.; Gordon, A.; Morrison, S. Evaluation of factors associated with immunoglobulin G, fat, protein, and lactose concentrations in bovine colostrum and colostrum management practices in grassland-based dairy systems in Northern Ireland. J. Dairy Sci. 2017, 100, 2068–2079. [Google Scholar] [CrossRef] [Green Version]
- Genc, M.; Coban, O. Effect of Some Environmental Factors on Colostrum Quality and Passive Immunity in Brown Swiss and Holstein Cattle. Ph.D. Thesis, Atatürk Uni., Health Sciences Institute, Erzurum, Turkey, 2015. [Google Scholar]
- Kuczaj, M.; Janik-Dubowiecka, A.; Szulc, T.; Zachwieja, A. Wpływ rasy i genotypu krów oraz liczby komórek somatycznych na wybrane cechy siary. Med. Weter. 2006, 62, 1403–1406. [Google Scholar]
- Puppel, K.; Gołębiewski, M.; Grodkowski, G.; Solarczyk, P.; Kostusiak, P.; Klopčič, M.; Sakowski, T. Use of somatic cell count as an indicator of colostrum quality. PLoS ONE 2020, 15, e0237615. [Google Scholar] [CrossRef] [PubMed]
- Włodarczyk, R.; Budvytis, M. Właściwe żywienie krów wysoko wydajnych–jak w pełni wykorzystać ich potencjał produkcyjny. Życie Weter. 2011, 86, 771–776. [Google Scholar]
- Nowak, W.; Mikuła, R.; Kasprowicz-Potocka, M.; Ignatowicz, M.; Zachwieja, A.; Paczyńska, K.; Pecka, E. Effect of cow nutrition in the far-off period on colostrum quality and immune response of calves. Bull. Vet. Inst. Pulawy 2012, 56, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Mann, S.; Yepes, F.L.; Overton, T.; Lock, A.; Lamb, S.; Wakshlag, J.; Nydam, D. Effect of dry period dietary energy level in dairy cattle on volume, concentrations of immunoglobulin G, insulin, and fatty acid composition of colostrum. J. Dairy Sci. 2016, 99, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Jolazadeh, A.; Mohammadabadi, T.; Dehghan-Banadaky, M.; Chaji, M.; Garcia, M. Effect of supplementing calcium salts of n-3 and n-6 fatty acid to pregnant nonlactating cows on colostrum composition, milk yield, and reproductive performance of dairy cows. Anim. Feed. Sci. Technol. 2019, 247, 127–140. [Google Scholar] [CrossRef]
- Halik, G.; Łozicki, A.; Koziorzębska, A.; Arkuszewska, E.; Puppel, K. Effect of the diets with pumpkin silage and synthetic β-carotene on the carotenoid, immunoglobulin and bioactive protein content and fatty acid composition of colostrum. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Guliński, P.; Młynek, K.; Giersz, B. Wpływ długosci okresu po wycieleniu i wieku krów na poziom immunoglobulin w siarze. Rocz. Nauk. Zootech 2006, 33, 193–200. [Google Scholar]
- Wąsowska, E.; Puppel, K. Changes in the content of immunostimulating components of colostrum obtained from dairy cows at different levels of production. J. Sci. Food Agric. 2018, 98, 5062–5068. [Google Scholar] [CrossRef]
- Cortese, V.S. Neonatal immunology. Vet. Clin. North Am. Food Anim. Pract. 2009, 25, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Stefaniak, T.; Chełmońska-Soyta, A.; Bajzert, J.; Jawor, P.; Rząsa, A.; Sitnik, O. Rozwój układu odpornościowego u przeżuwaczy w okresie pre-i postnatalnym. Med. Weter. 2012, 68, 534–539. [Google Scholar]
- Puppel, K.; Kuczyńska, B.; Nałęcz-Tarwacka, T.; Sakowski, T.; Gołębiewski, M.; Kunowska-Slósarz, M.; Budziński, A.; Grodzki, H. Effect of fish oil and linseed supplementation on the protein composition of milk from cows with different β-lactoglobulin phenotypes. J. Sci. Food Agric. 2014, 94, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Choque, B.; Catheline, D.; Rioux, V.; Legrand, P. Linoleic acid: Between doubts and certainties. Biochimie 2014, 96, 14–21. [Google Scholar] [CrossRef]
- Haghighatdoost, F.; Hariri, M. The effect of alpha-lipoic acid on inflammatory mediators: A systematic review and meta-analysis on randomized clinical trials. Eur. J. Pharmacol. 2019, 849, 115–123. [Google Scholar] [CrossRef]
- Opgenorth, J.; Sordillo, L.M.; Lock, A.L.; Gandy, J.C.; VandeHaar, M.J. Colostrum supplementation with n-3 fatty acids alters plasma polyunsaturated fatty acids and inflammatory mediators in newborn calves. J. Dairy Sci. 2020, 103, 11676–11688. [Google Scholar] [CrossRef]
- Śpitalniak-Bajerska, K.; Szumny, A.; Pogoda-Sewerniak, K.; Kupczyński, R. Effects of n-3 fatty acids on growth, antioxidant status, and immunity of preweaned dairy calves. J. Dairy Sci. 2020, 103, 2864–2876. [Google Scholar] [CrossRef]
- Mašek, T.; Krstulović, L.; Brozić, D.; Vranić, M.; Maurić, M.; Bajić, M.; Starčević, K. Cow colostrum and early milk enriched with eicosapentaenoic and docosahexaenoic fatty acid. Eur. Food Res. Technol. 2014, 238, 635–640. [Google Scholar] [CrossRef]
- Leiber, F.; Hochstrasser, R.; Wettstein, H.-R.; Kreuzer, M. Feeding transition cows with oilseeds: Effects on fatty acid composition of adipose tissue, colostrum and milk. Livest. Sci. 2011, 138, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, D.; Dell’Orto, V.; Varisco, G.; Agazzi, A.; Savoini, G. Enrichment in n− 3 fatty acids of goat’s colostrum and milk by maternal fish oil supplementation. Small Rumin. Res. 2006, 64, 22–29. [Google Scholar] [CrossRef]
- Annett, R.; Dawson, L.; Edgar, H.; Carson, A. Effects of source and level of fish oil supplementation in late pregnancy on feed intake, colostrum production and lamb output of ewes. Anim. Feed. Sci. Technol. 2009, 154, 169–182. [Google Scholar] [CrossRef]
- Santschi, D.; Wettstein, H.-R.; Leiber, F.; Witschi, A.-K.M.; Kreuzer, M. Colostrum and milk fatty acids of dairy cows as influenced by extruded linseed supplementation during the transition period. Can. J. Anim. Sci. 2009, 89, 383–392. [Google Scholar] [CrossRef]
- Puppel, K.; Kuczyńska, B.; Nałęcz-Tarwacka, T.; Grodzki, H. Influence of linseed variety on fatty acids profile in cow milk. J. Sci. Food Agric. 2013, 93, 2276–2280. [Google Scholar] [CrossRef] [PubMed]
- EN ISO 5509; Animal and Vegetable Fats and Oils- Preparation of Methyl Esters of Fatty Acids (ISO 5509:2000). Available online: https://www.iso.org/standard/11560.html (accessed on 21 January 2023).
- IBM Corp. Released IBM SPSS for Windows, Version 23.0. Armonk, N. 2022. Available online: https://www.ibm.com/support/pages/downloading-ibm-spss-statistics-25 (accessed on 21 January 2023).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feed (kg/cow/day) | Dry Cows Groups | |||
|---|---|---|---|---|
| I (First 5 Weeks) | II (Last 3 Weeks) | |||
| Dry Matter (kg) | Feed (kg) | Dry Matter (kg) | Feed (kg) | |
| Roughage: | 12.12 | 24.20 | 9.25 | 23.50 |
| Maize silage | 4.65 | 16.00 | ||
| Alfalfa silage | 3.05 | 5.55 | ||
| Grass silage | 7.65 | 19.55 | ||
| Corn grain | 0.42 | 0.87 | ||
| Straw | 3.26 | 3.69 | 1.20 | 1.30 |
| Concentrates: | 0.65 | 0.65 | 2.48 | 2.80 |
| Fodder chalk | 0.05 | 0.05 | ||
| Prophos Trans | 0.15 | 0.15 | 0.15 | 0.15 |
| Rape meal | 0.45 | 0.50 | 0.53 | 0.60 |
| Soya meal | 0.92 | 1.20 | ||
| Grain meal | 0.86 | 1.10 | ||
| Fatty Acid | Fish Oil [g/kg] | Linseed [g/kg] |
|---|---|---|
| C14:0 | 47.5 | 0.4 |
| C18:0 | 2.5 | 42.7 |
| C18:1 cis-9 | 251.4 | 167.7 |
| C18:2n-6 | 42.3 | 138.9 |
| C18:3n-3 | 31.1 | 561.6 |
| C20:5n-3 | 73.2 | 0.00 |
| C22:5n-3 | 7.7 | 0.00 |
| C22:6n-3 | 134.4 | 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grodkowska, K.; Gołębiewski, M.; Slósarz, J.; Sakowski, T.; Puppel, K. The Effect of Supplementation Using a Mixture of Fish Oil and Linseed on the Level of Immunomodulatory Components in Bovine Colostrum. Molecules 2023, 28, 2154. https://doi.org/10.3390/molecules28052154
Grodkowska K, Gołębiewski M, Slósarz J, Sakowski T, Puppel K. The Effect of Supplementation Using a Mixture of Fish Oil and Linseed on the Level of Immunomodulatory Components in Bovine Colostrum. Molecules. 2023; 28(5):2154. https://doi.org/10.3390/molecules28052154
Chicago/Turabian StyleGrodkowska, Kinga, Marcin Gołębiewski, Jan Slósarz, Tomasz Sakowski, and Kamila Puppel. 2023. "The Effect of Supplementation Using a Mixture of Fish Oil and Linseed on the Level of Immunomodulatory Components in Bovine Colostrum" Molecules 28, no. 5: 2154. https://doi.org/10.3390/molecules28052154
APA StyleGrodkowska, K., Gołębiewski, M., Slósarz, J., Sakowski, T., & Puppel, K. (2023). The Effect of Supplementation Using a Mixture of Fish Oil and Linseed on the Level of Immunomodulatory Components in Bovine Colostrum. Molecules, 28(5), 2154. https://doi.org/10.3390/molecules28052154