


Study on the Interaction Mechanism of Methoxy Polyethylene Glycol Maleimide with Sweet Potato β-Amylase

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of the Solutions
2.3. Enzyme Assay
2.4. Infrared Spectroscopy Measurements
2.5. Circular Dichroism (CD) Measurements
2.6. Differential Scanning Calorimetry (DSC) Measurements
2.7. Isothermal Titration Calorimetry (ITC) Measurements
2.8. UV–Vis Absorption Spectroscopy
2.9. Fluorescence Spectroscopy
2.10. Statistical Analysis
3. Results and Discussion
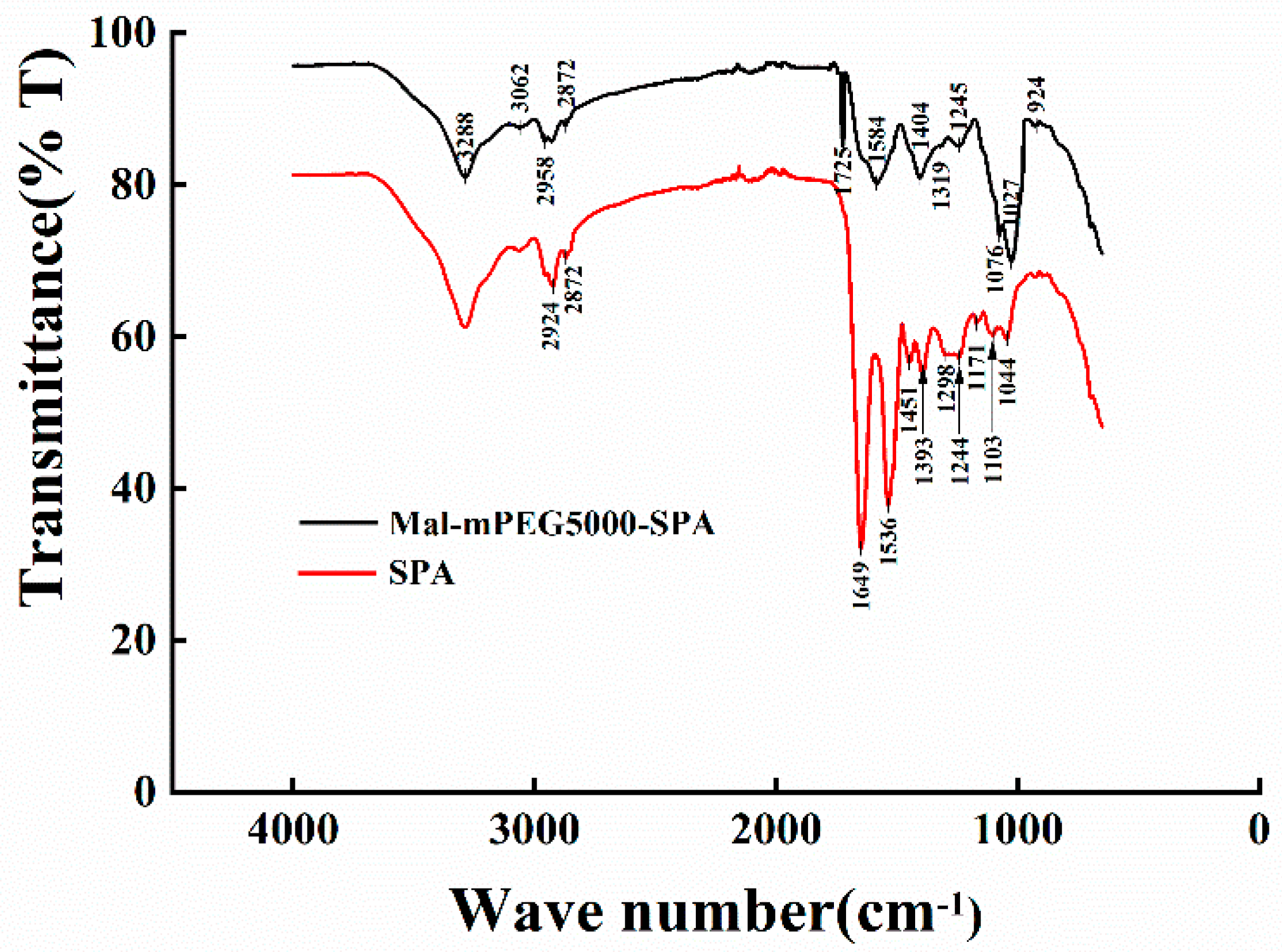
3.1. Infrared Spectra Analysis
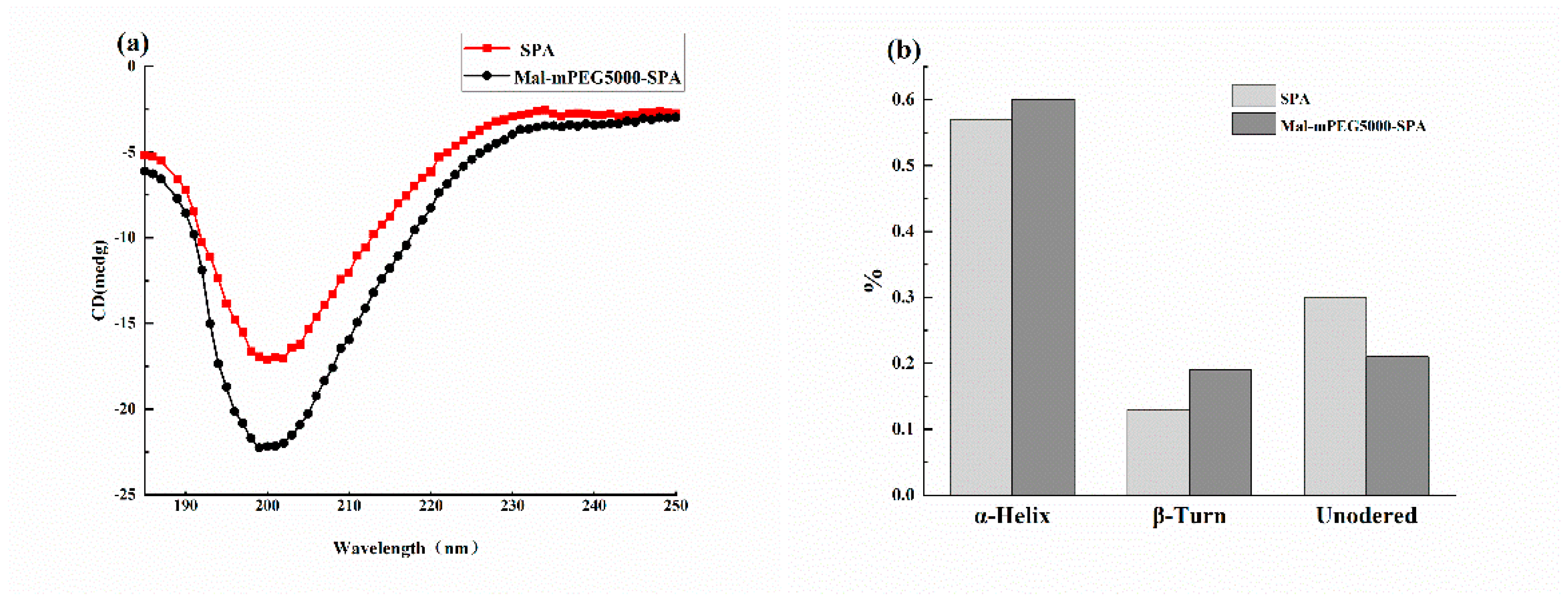
3.2. Circular Dichroism Analysis
3.3. DSC Result
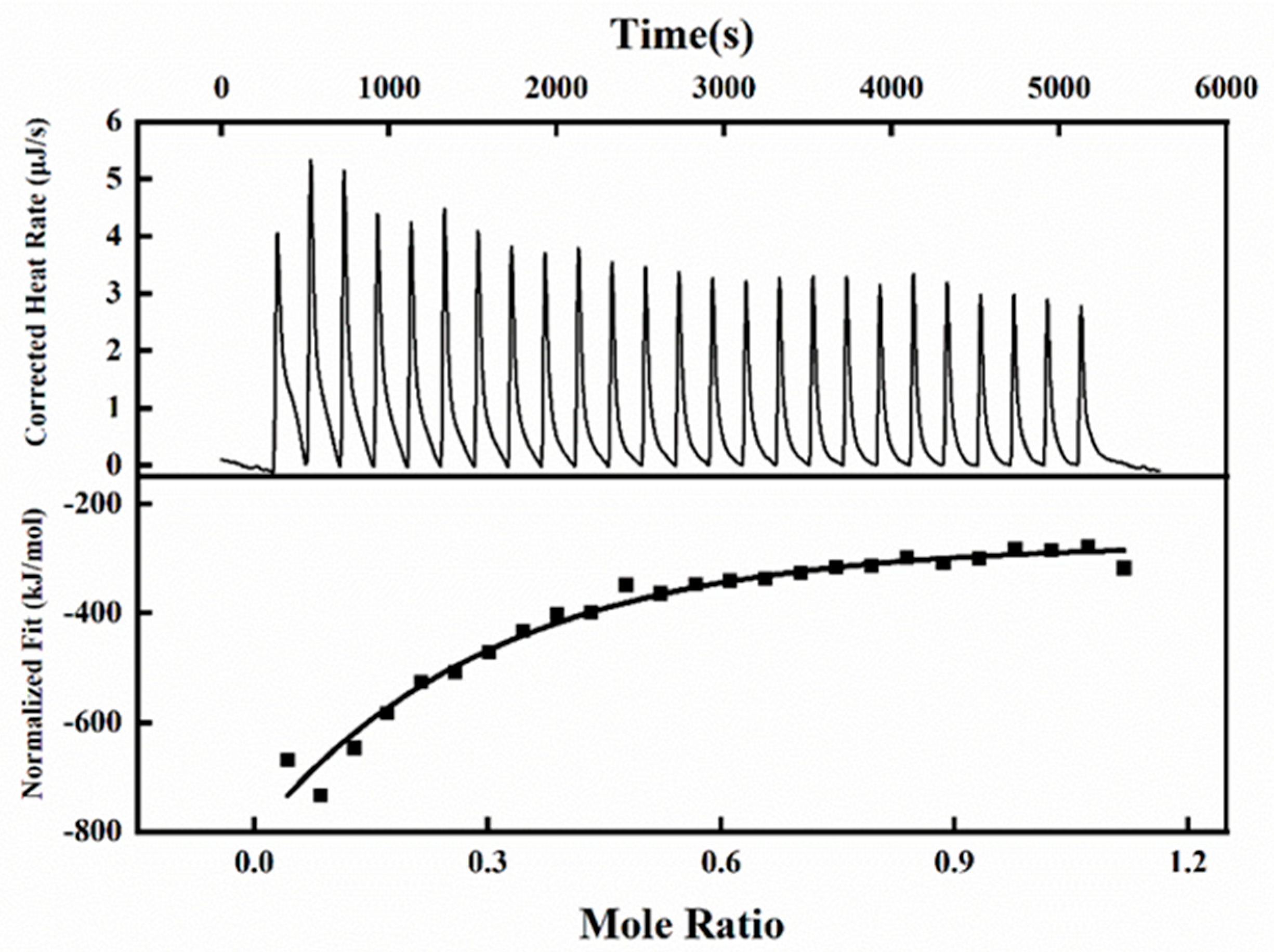
3.4. Isothermal Titration Calorimetry Investigation
3.5. UV–Vis Absorption Spectra
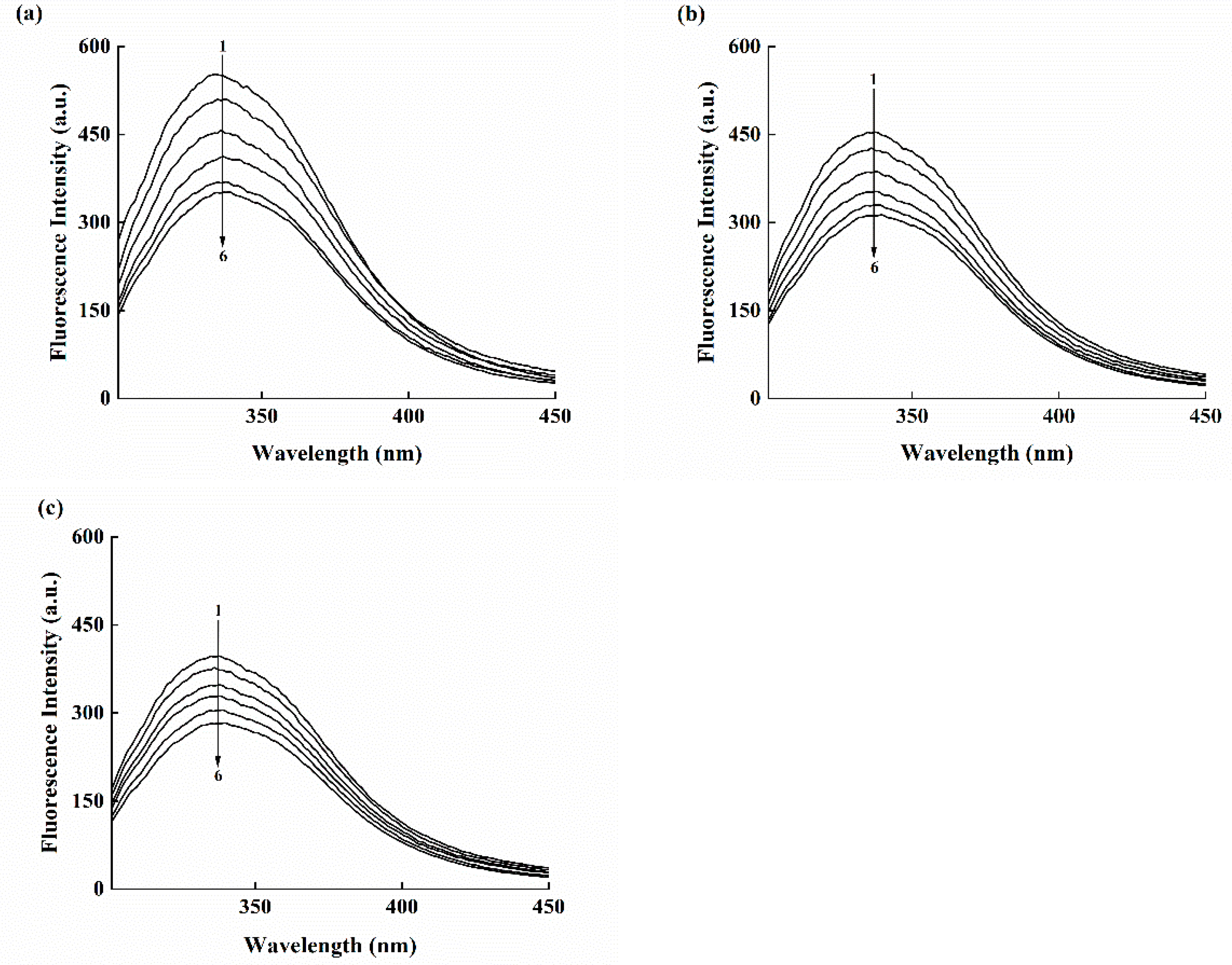
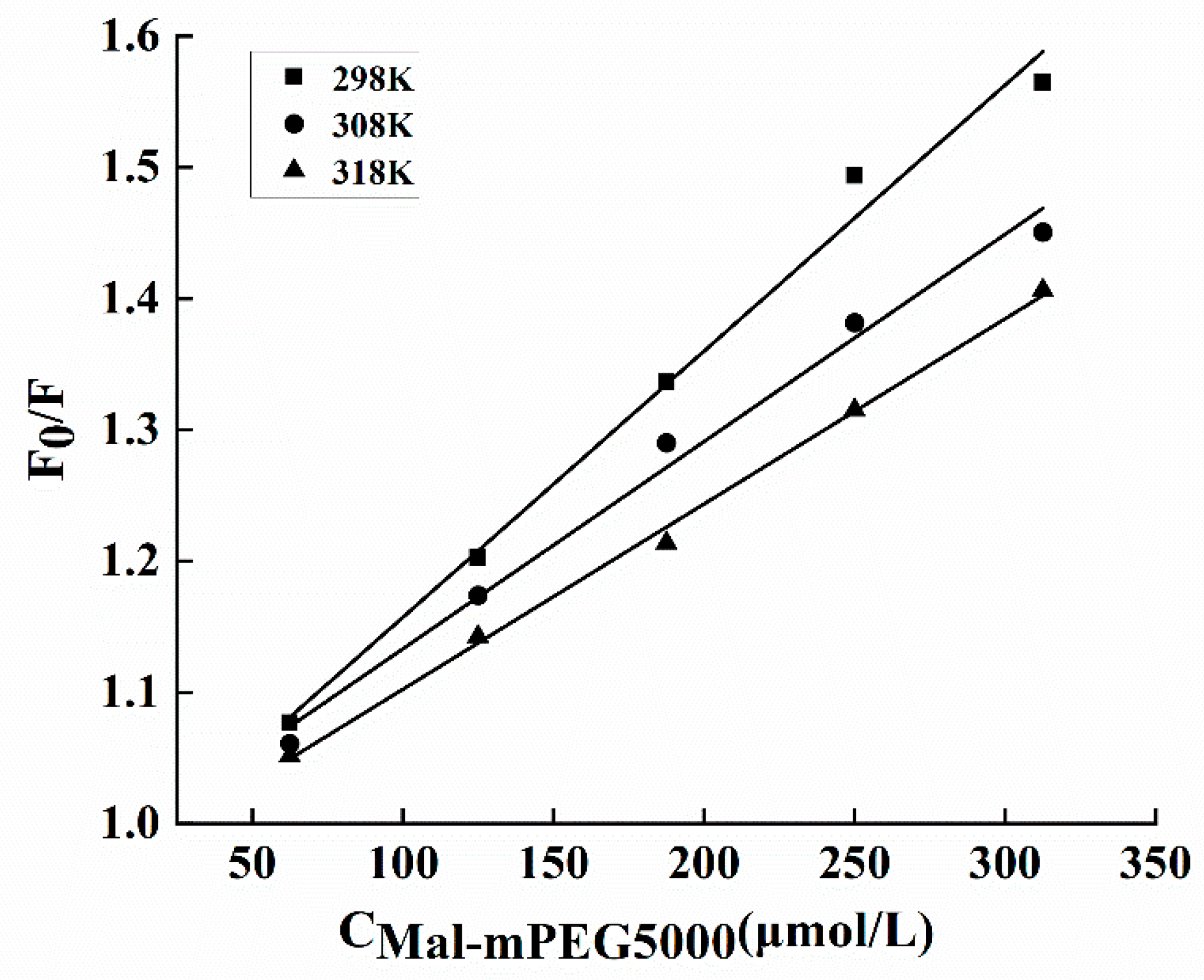
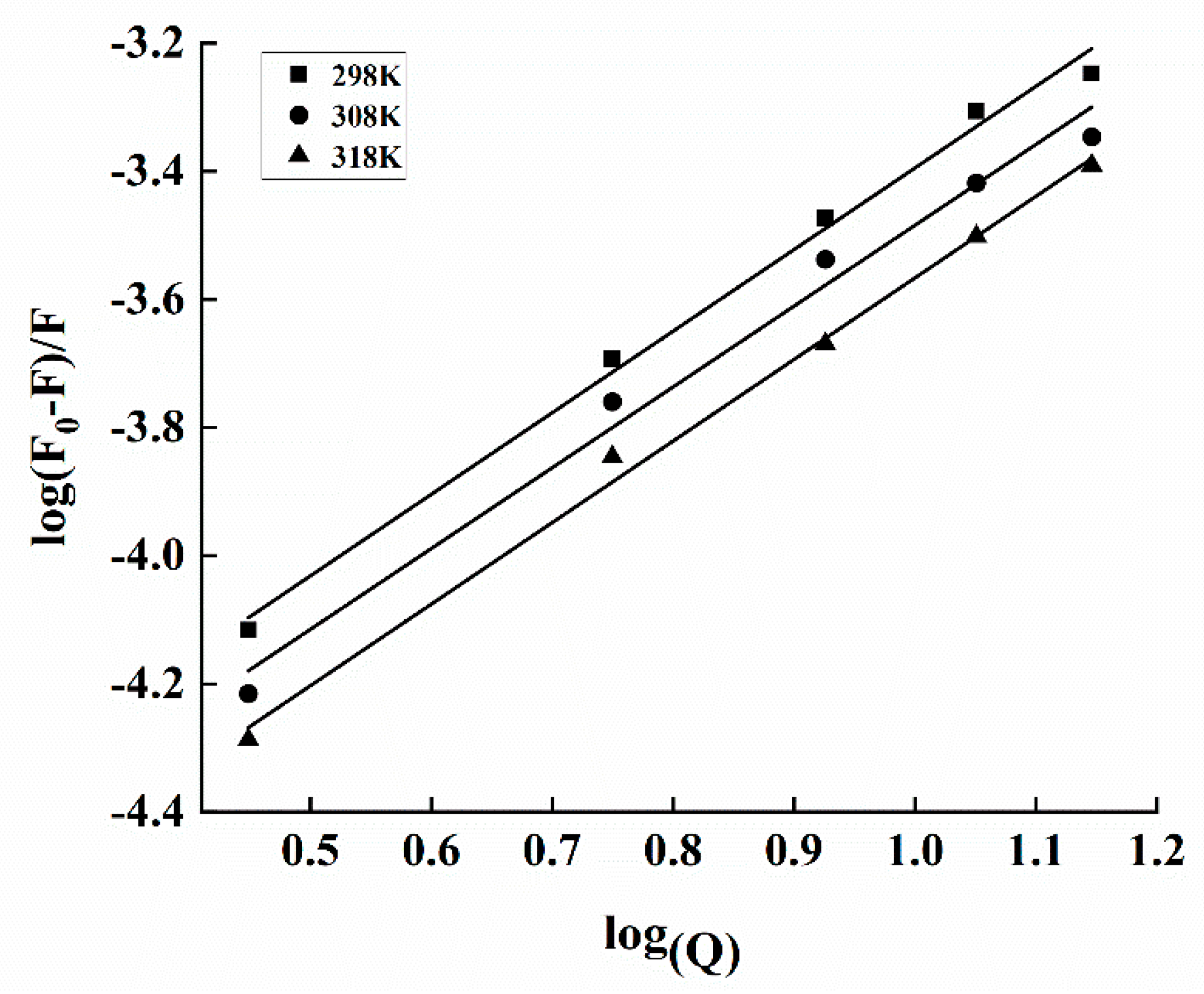
3.6. Fluorescence Measurements for Binding Constant and Site
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Gui, Y.; Zou, F.; Li, J.; Zhu, Y.; Guo, L.; Cui, B. The structural and functional properties of corn starch treated with endogenous malt amylases. Food Hydrocoll. 2021, 117, 106722. [Google Scholar] [CrossRef]
- Baker, W.L.; Smiley, K.L. Beta-amylase sulphydryl and disulphide group reactions: Additional aspects on enzyme inhibition by ascorbic acid. J. Inst. Brew. 1985, 91, 25–30. [Google Scholar] [CrossRef]
- Daba, T.; Kojima, K.; Inouye, K. Chemical modification of wheat beta-amylase by trinitrobenzenesulfonic acid, methoxypolyethylene glycol, and glutaraldehyde to improve its thermal stability and activity. Enzym. Microb. Technol. 2013, 53, 420–426. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases, α and β-sciencedirect. Methods Enzymol. 1955, 1, 149–158. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, W.; Ran, J.; Sun, J.; Jiao, L.; Feng, L.; Liu, B. Chemical modification of sweet potato β-amylase by Mal-mPEG to improve its enzymatic characteristics. Molecules 2018, 23, 2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.; Ouyang, F.; Liang, X.; Guo, X.; Dang, Z.; Zheng, L. Sorption of crude oil by enzyme-modified corn stalk vs. chemically treated corn stalk. J. Mol. Liq. 2018, 255, 324–332. [Google Scholar] [CrossRef]
- Peng, Z.; Ji, C.; Zhou, Y.; Zhao, T.; Leblanc, R.M. Polyethylene glycol (PEG) derived carbon dots: Preparation and applications. Appl. Mater. Today 2020, 20, 100677. [Google Scholar] [CrossRef]
- Shariatinia, Z.; Oroujzadeh, N. Anticancer drug delivery shuttles based on polyethylene glycol-polylactic acid nanocomposites: Molecular dynamics simulations. J. Nanostruct. 2021, 11, 347–367. [Google Scholar] [CrossRef]
- Abuchowski, A.; Van Es, T.; Palczuk, N.C.; Davis, F.F. Alteration of immunological properties of bovine serum albumin by covalent attachment of polyethylene glycol. J. Biol. Chem. 1977, 252, 3578–3581. [Google Scholar] [CrossRef]
- Huckaby, J.T.; Jacobs, T.M.; Li, Z.; Perna, R.J.; Wang, A.; Nicely, N.I.; Lai, S.K. Structure of an anti-PEG antibody reveals an open ring that captures highly flexible PEG polymers. Commun. Chem. 2020, 3, 124. [Google Scholar] [CrossRef]
- Gupta, V.; Bhavanasi, S.; Quadir, M.; Singh, K.; Ghosh, G.; Vasamreddy, K.; Ghosh, A.; Siahaan, T.J.; Banerjee, G.; Banerjee, S.K. Protein PEGylation for cancer therapy: Bench to bedside. J. Cell Commun. Signal. 2019, 13, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Brianezi, S.F.S.; Castro, K.C.; Piazza, R.D.; Melo, M.D.S.F.; Pereira, R.M.; Marques, R.F.C.; Campos, M.G.N. Preparation and characterization of chitosan/mPEG-PCL blended membranes for wound dressing and controlled gentamicin release. Mater. Res. 2018, 21, 20170951. [Google Scholar] [CrossRef]
- Chen, B.M.; Cheng, T.L.; Roffler, S.R. Polyethylene glycol immunogenicity: Theoretical, clinical, and practical aspects of anti-polyethylene glycol antibodies. ACS Nano 2021, 15, 14022–14048. [Google Scholar] [CrossRef] [PubMed]
- Serra, S. Enzymes, Biocatalysis and Chemical Biology. Molecules 2020, 25, 2354. [Google Scholar] [CrossRef]
- Sagu, S.T.; Nso, E.J.; Homann, T.; Kapseu, C.; Rawel, H.M. Extraction and purification of beta-amylase from stems of Abrus precatorius by three phase partitioning. Food Chem. 2015, 183, 144–153. [Google Scholar] [CrossRef]
- Inthamat, P.; Boonsiriwit, A.; Lee, Y.S.; Siripatrawan, U. Effects of genipin as natural crosslinker on barrier and mechanical properties of chitosan-astaxanthin film. J. Food Process. Preserv. 2022, 46, e15707. [Google Scholar] [CrossRef]
- Hu, G.; Wang, D.; Sun, L.; Su, R.; Corazzin, M.; Sun, X.; Dou, L.; Zhang, M.; Zhao, L.; Su, L.; et al. Isolation, Purification and Structure Identification of a Calcium-Binding Peptide from Sheep Bone Protein Hydrolysate. Foods 2022, 11, 2655. [Google Scholar] [CrossRef]
- Basu, A.; Kumar, G.S. Thermodynamics of the interaction of the food additive tartrazine with serum albumins: A microcalorimetric investigation. Food Chem. 2015, 175, 137–142. [Google Scholar] [CrossRef]
- He, F.; Chu, S.; Sun, N.; Li, X.; Jing, M.; Wan, J.; Tang, J.; Liu, R. Binding interactions of acrylamide with lysozyme and its underlying mechanisms based on multi-spectra, isothermal titration microcalorimetry and docking simulation. J. Mol. Liq. 2021, 337, 116460. [Google Scholar] [CrossRef]
- Pitchumani Krishnaveni, M.; Annadurai, G. Biosynthesis of nanoceria from bacillus subtilis: Characterization and antioxidant potential. Res. J. Life Sci. 2019, 5, 644. [Google Scholar] [CrossRef]
- Dai, T.; Chen, J.; McClements, D.J.; Hu, P.; Ye, X.; Liu, C.; Li, T. Protein–polyphenol interactions enhance the antioxidant capacity of phenolics: Analysis of rice glutelin–procyanidin dimer interactions. Food Funct. 2019, 10, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Lissi, E.; Abuin, E. On the evaluation of the number of binding sites in proteins from steady state fluorescence measurements. J. Fluoresc. 2011, 21, 1831–1833. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Talat, M.; Srivastava, O.N.; Kayastha, A.M. Covalent immobilization of peanut β-amylase for producing industrial nano-biocatalysts: A comparative study of kinetics, stability and reusability of the immobilized enzyme. Food Chem. 2018, 245, 488–499. [Google Scholar] [CrossRef]
- He, W.; Mu, H.; Liu, Z.; Lu, M.; Hang, F.; Chen, J.; Zeng, M.; Qin, F.; He, Z. Effect of preheat treatment of milk proteins on their interactions with cyanidin-3-O-glucoside. Food Res. Int. 2018, 107, 394–405. [Google Scholar] [CrossRef]
- Ianeselli, A.; Orioli, S.; Spagnolli, G.; Faccioli, P.; Cupellini, L.; Jurinovich, S.; Mennucci, B. Atomic detail of protein folding revealed by an ab initio reappraisal of circular dichroism. J. Am. Chem. Soc. 2018, 140, 3674–3682. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ji, H.; Bai, Y.; Jin, Z. Development of pullulanase mutants to enhance starch substrate utilization for efficient production of β-CD. Int. J. Biol. Macromol. 2021, 168, 640–648. [Google Scholar] [CrossRef]
- Ajorlou, S.; Ramezan, Y.; Bahmaei, M. Investigation on Thermal Stability of Peanut and Rice Bran Oils and Their Blends by DSC. J. Food Process. Preserv. 2021, 13, 19–32. [Google Scholar] [CrossRef]
- Huang, Z.; Ma, C.; Wu, M.; Li, X.; Lu, C.; Zhang, X.; Wu, C. Exploring the drug-lipid interaction of weak-hydrophobic drug loaded solid lipid nanoparticles by isothermal titration calorimetry. J. Nanoparticle Res. 2020, 22, 3. [Google Scholar] [CrossRef]
- Makarska-Bialokoz, M. Interactions of hemin with bovine serum albumin and human hemoglobin: A fluorescence quenching study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 193, 23–32. [Google Scholar] [CrossRef]
- Dahmane, I.; Montagner, C.; Matagne, A.; Dumbre, S.; Herdewijn, P.; Terrak, M. Peptidoglycan glycosyltransferase-ligand binding assay based on tryptophan fluorescence quenching. Biochimie 2018, 152, 1–5. [Google Scholar] [CrossRef]
- Song, F.; Wu, F.; Feng, W.; Tang, Z.; Giesy, J.P.; Guo, F.; Shi, D.; Liu, X.; Qin, N.; Xing, B.; et al. Fluorescence regional integration and differential fluorescence spectroscopy for analysis of structural characteristics and proton binding properties of fulvic acid sub-fractions. J. Environ. Sci. 2018, 74, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guan, H.; Zhang, H.; Liu, H.; Chen, Q.; Kong, B. Elucidation of interaction mechanisms between myofibrillar proteins and ethyl octanoate by SPME-GC-MS, molecular docking and dynamics simulation. LWT 2022, 154, 112787. [Google Scholar] [CrossRef]
- Liu, L.; Lan, W.; Pu, T.; Zhou, Y.; Xie, J. Combining slightly acidic electrolyzed water and slurry ice to prolong the shelf-life of mackerel (Pneumatophorus japonicus). J. Food Process. Preserv. 2021, 45, e15762. [Google Scholar] [CrossRef]
- Byadagi, K.; Meti, M.; Nandibewoor, S.; Chimatadar, S. Investigation of binding behaviour of procainamide hydrochloride with human serum albumin using synchronous, 3D fluorescence and circular dichroism. J. Pharm. Anal. 2017, 7, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Chen, J.; Li, Q.; Li, P.; Hu, P.; Liu, C.; Li, T. Investigation the interaction between procyanidin dimer and α-amylase: Spectroscopic analyses and molecular docking simulation. Int. J. Biol. Macromol. 2018, 113, 427–433. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, Y.; Xue, Z.; Gao, X.; Jia, Y.; Wang, Y.; Lu, Y.; Zhang, J.; Zhang, M.; Chen, H. Insight into the inactivation mechanism of soybean Bowman-Birk trypsin inhibitor (BBTI) induced by epigallocatechin gallate and epigallocatechin: Fluorescence, thermodynamics and docking studies. Food Chem. 2020, 303, 125380. [Google Scholar] [CrossRef]
- Prette, A.P.; Almeida, F.D.A.C.; Villa-Velez, H.A.; Telis-Romero, J. Thermodynamic properties of water sorption of jackfruit (Artocarpus heterophyllus Lam.) as a function of moisture content. Food Sci. Technol. 2013, 33, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Bi, S.; Wu, J.; Zhao, R.; Shao, D.; Song, Z. Multispectral and molecular docking investigations on the interaction of primethamine/trimethoprim with BSA/HSA. J. Biomol. Struct. Dyn. 2020, 38, 934–942. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhu, M.; Zhang, G.; Hu, X.; Pan, J. Novel insights into the interaction mechanism of 5-hydroxymethyl-2-furaldehyde with β-casein and its effects on the structure and function of β-casein. LWT 2021, 152, 112360. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, L.; Sun, C.; Zhao, D.; Tang, H. Studies on the structure-activity relationship and interaction mechanism of flavonoids and xanthine oxidase through enzyme kinetics, spectroscopy methods and molecular simulations. Food Chem. 2020, 323, 126807. [Google Scholar] [CrossRef]
- Lopes, C.A.; Roledo, C.; Reis, A.G.D. Moringa oleifera seed husks for methylene blue dye adsorption: Kinetic, equilibrium, and thermodynamic analyses. Rev. Ambiente Água 2022, 17. [Google Scholar] [CrossRef]
- Luo, L.; Chiu, L.Y.; Sugarman, A.; Gupta, P.; Rouskin, S.; Tolbert, B.S. HnRNP A1/A2 proteins assemble onto 7SK snRNA via context dependent interactions. J. Mol. Biol. 2021, 433, 166885. [Google Scholar] [CrossRef] [PubMed]
- Shaghaghi, M.; Rashtbari, S.; Vejdani, S.; Dehghan, G.; Jouyban, A.; Yekta, R. Exploring the interactions of a Tb (III)–quercetin complex with serum albumins (HSA and BSA): Spectroscopic and molecular docking studies. Luminescence 2020, 35, 512–524. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Initial Transition Temperature (°C) | Half-High Transition Temperature (°C) | Final Transition Temperature (°C) | Enthalpy (ΔH) (J/g) |
|---|---|---|---|---|
| SPA | 21.52 | 70.03 | 122.14 | 118.30 |
| Mal-mPEG5000-SPA | 23.15 | 74.22 | 127.90 | 149.71 |
| T(K) | Ksv·104/ (L·mol−1) | Kq·1012/ (L·mol−1·s−1) | KA·104/ (L·mol−1) | n | ΔHθ/ (kJ·mol−1) | ΔGθ/ (kJ·mol−1) | ΔSθ/ (J·mol−1·K−1) |
|---|---|---|---|---|---|---|---|
| 298 | 4.53 ± 0.14a | 4.53 ± 0.14a | 4.65 ± 0.16c | 1.27 ± 0.1a | 20.73 ± 0.34 | −26.63 ± 0.45a | 159.14 ± 0.81a |
| 308 | 3.53 ± 0.15b | 3.53 ± 0.15b | 5.56 ± 0.21b | 1.26 ± 0.04a | −27.98 ± 0.55b | 158.16 ± 0.83a | |
| 318 | 3.15 ± 0.13c | 3.15 ± 0.13c | 6.91 ± 0.18a | 1.21 ± 0.03b | −29.46 ± 0.26c | 157.83 ± 0.91a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Kong, Y.; Sun, H.; Zhao, R.; Jiao, L.; Zhang, W.; Liu, B. Study on the Interaction Mechanism of Methoxy Polyethylene Glycol Maleimide with Sweet Potato β-Amylase. Molecules 2023, 28, 2188. https://doi.org/10.3390/molecules28052188
Liang X, Kong Y, Sun H, Zhao R, Jiao L, Zhang W, Liu B. Study on the Interaction Mechanism of Methoxy Polyethylene Glycol Maleimide with Sweet Potato β-Amylase. Molecules. 2023; 28(5):2188. https://doi.org/10.3390/molecules28052188
Chicago/Turabian StyleLiang, Xinhong, Yaxin Kong, Huadi Sun, Ruixiang Zhao, Lingxia Jiao, Wanli Zhang, and Bing Liu. 2023. "Study on the Interaction Mechanism of Methoxy Polyethylene Glycol Maleimide with Sweet Potato β-Amylase" Molecules 28, no. 5: 2188. https://doi.org/10.3390/molecules28052188
APA StyleLiang, X., Kong, Y., Sun, H., Zhao, R., Jiao, L., Zhang, W., & Liu, B. (2023). Study on the Interaction Mechanism of Methoxy Polyethylene Glycol Maleimide with Sweet Potato β-Amylase. Molecules, 28(5), 2188. https://doi.org/10.3390/molecules28052188