
Reprotoxic Effect of Tris(2,3-Dibromopropyl) Isocyanurate (TBC) on Spermatogenic Cells In Vitro



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
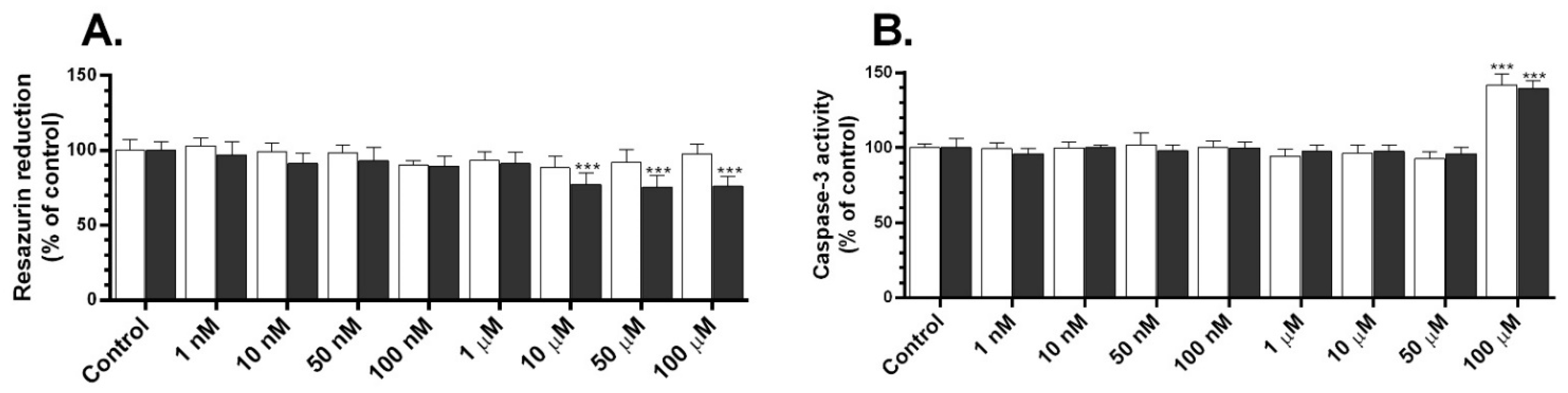
2.1. Metabolic Activity and Caspase-3 Activity of TBC
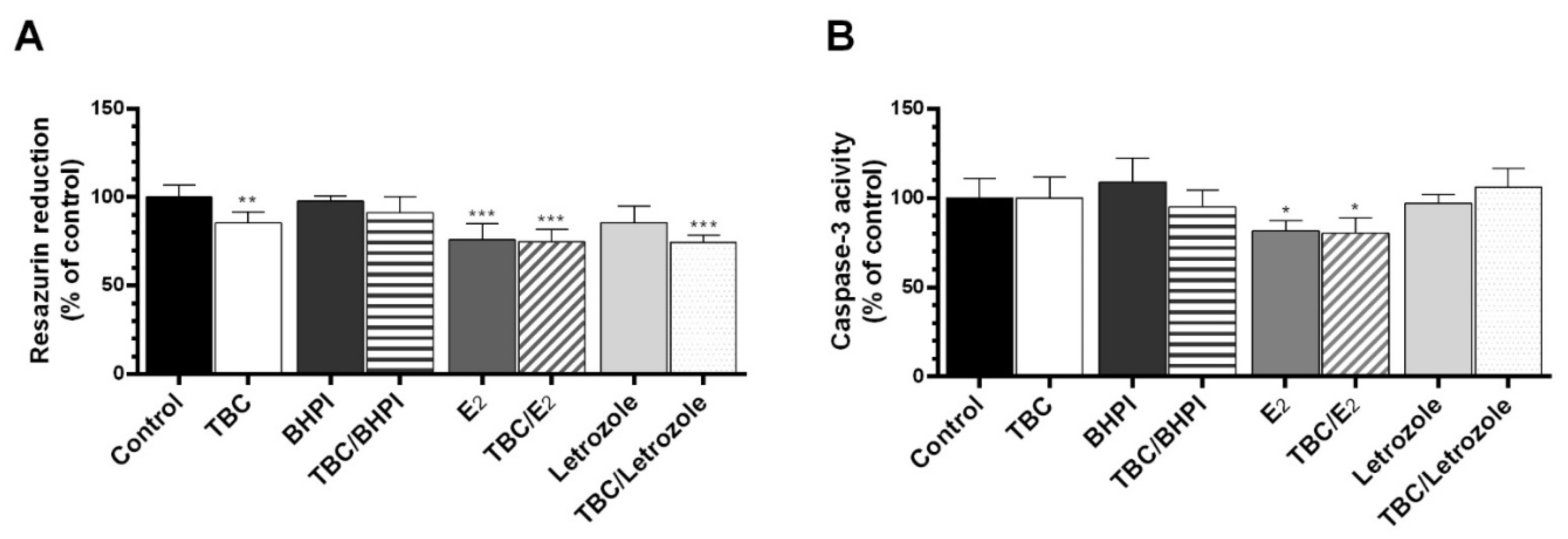
2.2. Cotreatment of the Cells with BHPI, E2, and Letrozole
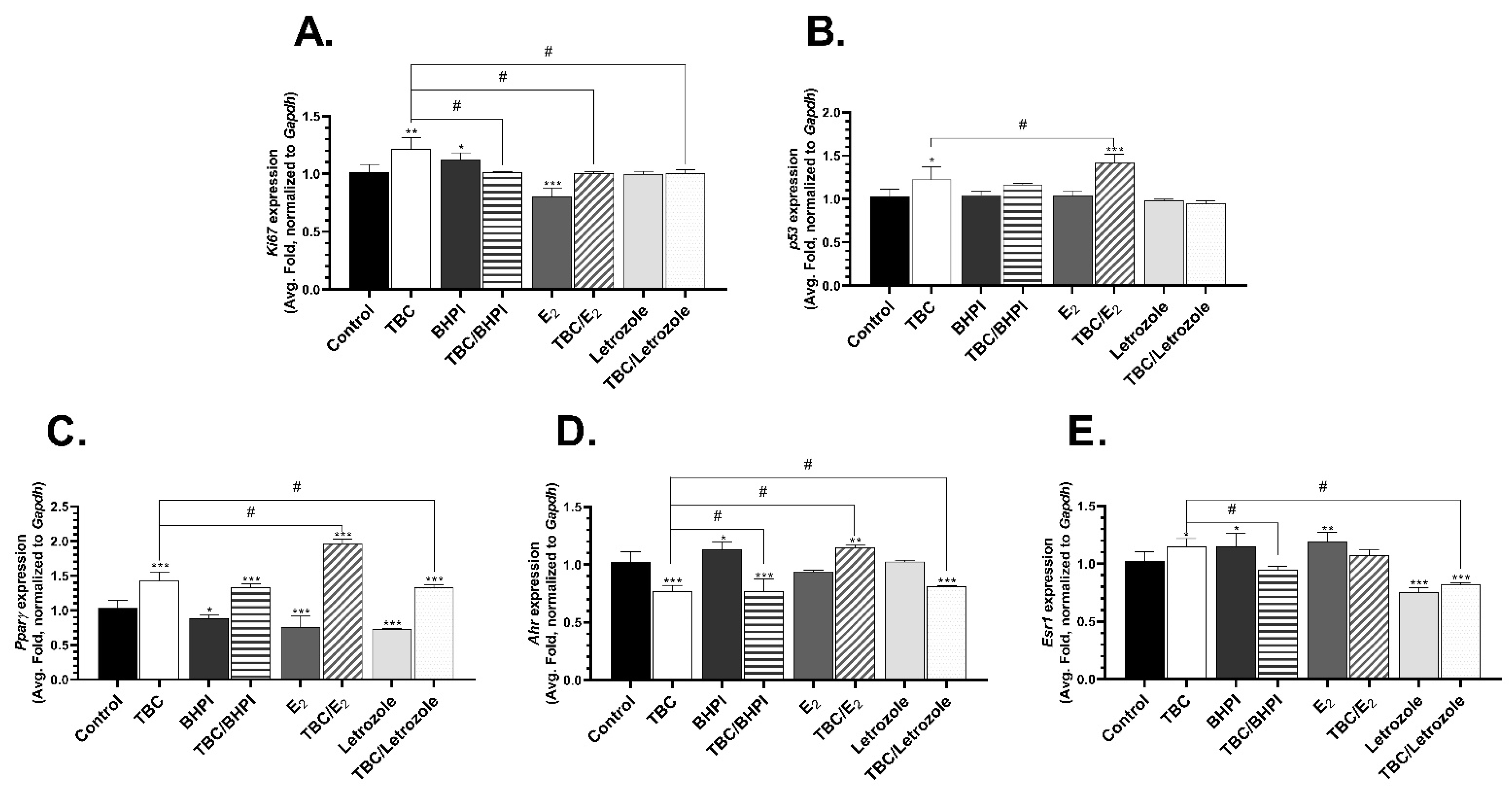
2.3. Level of mRNA Expression of Ki67, p53, Pparγ, Ahr, Esr1, and Esr2
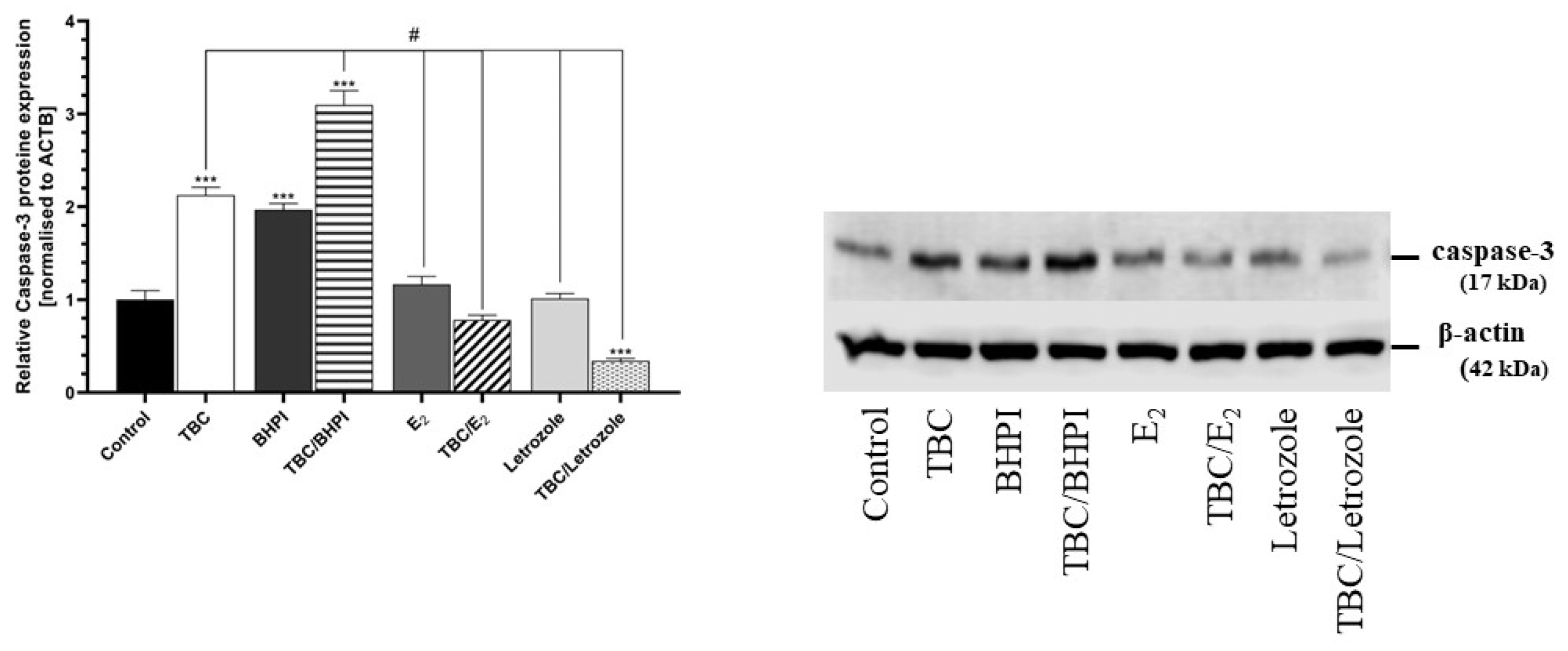
2.4. Caspase-3 Protein Expression
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture and Treatment
4.3. Resazurin Reduction Assay
4.4. Caspase-3 Activity
4.5. Western Blotting
4.6. Real-Time PCR
4.7. Statistical Analysis
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.-L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Sun, H.; Gong, T.-T.; Jiang, Y.-T.; Zhang, S.; Zhao, Y.-H.; Wu, Q.-J. Global, regional, and national prevalence and disability-adjusted life-years for infertility in 195 countries and territories, 1990–2017: Results from a global burden of disease study, 2017. Aging 2019, 11, 10952–10991. [Google Scholar] [CrossRef]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Bernhard, A.; Berntssen, M.H.G.; Lundebye, A.-K.; Røyneberg Alvheim, A.; Secher Myrmel, L.; Fjære, E.; Torstensen, B.E.; Kristiansen, K.; Madsen, L.; Brattelid, T.; et al. Marine fatty acids aggravate hepatotoxicity of α-HBCD in juvenile female BALB/c mice. Food Chem. Toxicol. 2016, 97, 411–423. [Google Scholar] [CrossRef]
- Li, X.; Pan, Y.; Wang, C.; Chen, M.; Liu, Y.; Li, J.; Zhou, Z.; Xu, J.; Liang, Y.; Song, M. Effects of tris(2,3-dibromopropyl) isocyanurate on steroidogenesis in H295R cells. Environ. Earth Sci. 2016, 75, 1339. [Google Scholar] [CrossRef]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A Systematic Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, K.J.; Tan, Z.; Zheng, C.M.; Liang, Z.; Zhao, J.Q. Identification of potential therapeutic targets for papillary thyroid carcinoma by bioinformatics analysis. Oncol. Lett. 2016, 11, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xia, H.; Wang, L.; Xia, G.; Qu, Y.; Shang, X.; Lin, S. Lignans from the Twigs of Litsea cubeba and Their Bioactivities. Molecules 2019, 24, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Wang, P.; Wei, Y.; Liu, Q.; Yang, C.; Zhao, G.; Yue, J.; Yan, X.; Zhou, Z. Characterization of Panax ginseng UDP-Glycosyltransferases Catalyzing Protopanaxatriol and Biosyntheses of Bioactive Ginsenosides F1 and Rh1 in Metabolically Engineered Yeasts. Mol. Plant 2015, 8, 1412–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johns, L.E.; Ferguson, K.K.; Soldin, O.P.; Cantonwine, D.E.; Rivera-González, L.O.; Del Toro, L.V.A.; Calafat, A.M.; Ye, X.; Alshawabkeh, A.N.; Cordero, J.F.; et al. Urinary phthalate metabolites in relation to maternal serum thyroid and sex hormone levels during pregnancy: A longitudinal analysis. Reprod. Biol. Endocrinol. 2015, 13, 4. [Google Scholar] [CrossRef] [Green Version]
- Tournaire, M.; Devouche, E.; Espié, M.; Asselain, B.; Levadou, A.; Cabau, A.; Dunbavand, A.; Grosclaude, P.; Epelboin, S. Cancer Risk in Women Exposed to Diethylstilbestrol in Utero. Therapies 2015, 70, 433–441. [Google Scholar] [CrossRef]
- Costa, E.M.F.; Spritzer, P.M.; Hohl, A.; Bachega, T.A.S.S. Efeitos dos desreguladores endócrinos no desenvolvimento do trato reprodutivo feminino. Arq. Bras. Endocrinol. Metabol. 2014, 58, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Yu, D.; Yang, L.; Sui, S.; Lv, S.; Bai, Y.; Sun, W.; Wang, Y.; Chen, L.; Sun, Z.; et al. Thyroid function and decabromodiphenyl ethane (DBDPE) exposure in Chinese adults from a DBDPE manufacturing area. Environ. Int. 2019, 133, 105179. [Google Scholar] [CrossRef]
- Eng, M.L.; Karouna-Renier, N.K.; Henry, P.F.P.; Letcher, R.J.; Schultz, S.L.; Bean, T.G.; Peters, L.E.; Palace, V.P.; Williams, T.D.; Elliott, J.E.; et al. In ovo exposure to brominated flame retardants Part II: Assessment of effects of TBBPA-BDBPE and BTBPE on hatching success, morphometric and physiological endpoints in American kestrels. Ecotoxicol. Environ. Saf. 2019, 179, 151–159. [Google Scholar] [CrossRef]
- Krivoshiev, B.V.; Dardenne, F.; Covaci, A.; Blust, R.; Husson, S.J. Assessing in-vitro estrogenic effects of currently-used flame retardants. Toxicol. In Vitro 2016, 33, 153–162. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Liang, B.; Chen, T.; Zheng, D.; Zhao, X.; Jing, L.; Zhou, X.; Sun, Z.; Shi, Z. Hepatotoxicity of decabromodiphenyl ethane (DBDPE) and decabromodiphenyl ether (BDE-209) in 28-day exposed Sprague-Dawley rats. Sci. Total Environ. 2020, 705, 135783. [Google Scholar] [CrossRef]
- Bar, M.; Szychowski, K.A. Comprehensive review of the impact of tris(2,3-dibromopropyl) isocyanurate (TBC or TDBP-TAZTO) on living organisms and the environment. Environ. Geochem. Health 2022, 44, 4203–4218. [Google Scholar] [CrossRef]
- Alzualde, A.; Behl, M.; Sipes, N.S.; Hsieh, J.-H.; Alday, A.; Tice, R.R.; Paules, R.S.; Muriana, A.; Quevedo, C. Toxicity profiling of flame retardants in zebrafish embryos using a battery of assays for developmental toxicity, neurotoxicity, cardiotoxicity and hepatotoxicity toward human relevance. Neurotoxicol. Teratol. 2018, 70, 40–50. [Google Scholar] [CrossRef]
- Honkisz, E.; Wójtowicz, A.K. The role of PPARγ in TBBPA-mediated endocrine disrupting effects in human choriocarcinoma JEG-3 cells. Mol. Cell. Biochem. 2015, 409, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Inoue, K.; Shimada, A.; Takano, H. Physiological effects of brominated flame retardants on NC/Nga mice. Immunopharmacol. Immunotoxicol. 2018, 40, 1–5. [Google Scholar] [CrossRef]
- Wojtowicz, A.K.; Szychowski, K.A.; Kajta, M. PPAR-γ Agonist GW1929 But Not Antagonist GW9662 Reduces TBBPA-Induced Neurotoxicity in Primary Neocortical Cells. Neurotox. Res. 2014, 25, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Li, X.; Zhang, W.; Wang, L.; Pan, Y.; Zhou, Z.; Chen, M.; Zhang, A.; Liang, Y.; Song, M. Anti-estrogenic activity of tris(2,3-dibromopropyl) isocyanurate through disruption of co-activator recruitment: Experimental and computational studies. Arch. Toxicol. 2018, 92, 1471–1482. [Google Scholar] [CrossRef] [PubMed]
- Hess, R.A.; Cooke, P.S. Estrogen in the male: A historical perspective†. Biol. Reprod. 2018, 99, 27–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, R.A. Estrogen in the adult male reproductive tract: A review. Reprod. Biol. Endocrinol. 2003, 1, 52. [Google Scholar] [CrossRef] [Green Version]
- Lambard, S.; Silandre, D.; Delalande, C.; Denis-Galeraud, I.; Bourguiba, S.; Carreau, S. Aromatase in testis: Expression and role in male reproduction. J. Steroid Biochem. Mol. Biol. 2005, 95, 63–69. [Google Scholar] [CrossRef]
- Yıldırım, A.B.; Kaymak, E. Aromatase, Estrogen and Male Reproduction: A Review. Gaziantep Islam Sci. Technol. Univ. 2020, 1, 100–108. [Google Scholar] [CrossRef]
- Nilsson, S.; Gustafsson, J.-Å. Estrogen Receptors: Therapies Targeted to Receptor Subtypes. Clin. Pharmacol. Ther. 2011, 89, 44–55. [Google Scholar] [CrossRef]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef] [Green Version]
- Eddy, E.M.; Washburn, T.F.; Bunch, D.O.; Goulding, E.H.; Gladen, B.C.; Lubahn, D.B.; Korach, K.S. Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinology 1996, 137, 4796–4805. [Google Scholar] [CrossRef]
- Dupont, S.; Krust, A.; Gansmuller, A.; Dierich, A.; Chambon, P.; Mark, M. Effect of single and compound knockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) on mouse reproductive phenotypes. Development 2000, 127, 4277–4291. [Google Scholar] [CrossRef]
- Warden, A.; Truitt, J.; Merriman, M.; Ponomareva, O.; Jameson, K.; Ferguson, L.B.; Mayfield, R.D.; Harris, R.A. Localization of PPAR isotypes in the adult mouse and human brain. Sci. Rep. 2016, 6, 27618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitti, M.; Di Emidio, G.; Di Carlo, M.; Carta, G.; Antonosante, A.; Artini, P.G.; Cimini, A.; Tatone, C.; Benedetti, E. Peroxisome Proliferator-Activated Receptors in Female Reproduction and Fertility. PPAR Res. 2016, 2016, 4612306. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-L.; Xian, H.; Cao, J.-C.; Zhang, C.; Zhang, Y.-H.; Chen, M.-M.; Qian, Y.; Jiang, M. Peroxisome proliferator-activated receptor gamma signaling in human sperm physiology. Asian J. Androl. 2015, 17, 942. [Google Scholar] [CrossRef]
- Bock, K.W. Aryl hydrocarbon receptor (AHR) functions in infectious and sterile inflammation and NAD+-dependent metabolic adaptation. Arch. Toxicol. 2021, 95, 3449–3458. [Google Scholar] [CrossRef]
- Barroso, A.; Mahler, J.V.; Fonseca-Castro, P.H.; Quintana, F.J. The aryl hydrocarbon receptor and the gut–brain axis. Cell. Mol. Immunol. 2021, 18, 259–268. [Google Scholar] [CrossRef]
- Ye, W.; Chen, R.; Chen, X.; Huang, B.; Lin, R.; Xie, X.; Chen, J.; Jiang, J.; Deng, Y.; Wen, J. AhR regulates the expression of human cytochrome P450 1A1 ( CYP1A1 ) by recruiting Sp1. FEBS J. 2019, 286, 4215–4231. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Hu, L.; Scrivens, P.J.; Batist, G. Transcriptional Regulation of NF-E2 p45-related Factor (NRF2) Expression by the Aryl Hydrocarbon Receptor-Xenobiotic Response Element Signaling Pathway. J. Biol. Chem. 2005, 280, 20340–20348. [Google Scholar] [CrossRef] [Green Version]
- Sommer, R.J.; Marks Sojka, K.; Pollenz, R.S.; Cooke, P.S.; Peterson, R.E. Ah Receptor and ARNT Protein and mRNA Concentrations in Rat Prostate: Effects of Stage of Development and 2,3,7,8-Tetrachlorodibenzo-p-Dioxin Treatment. Toxicol. Appl. Pharmacol. 1999, 155, 177–189. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Chen, M.; Wu, L.; Zhang, C.; Zhang, J.; Zhou, Q.; Liang, Y. Toxicity of the brominated flame retardant tris-(2,3-dibromopropyl) isocyanurate in zebrafish (Danio rerio). Chin. Sci. Bull. 2011, 56, 1548–1555. [Google Scholar] [CrossRef] [Green Version]
- Kekäläinen, J. Genetic incompatibility of the reproductive partners: An evolutionary perspective on infertility. Hum. Reprod. 2021, 36, 3028–3035. [Google Scholar] [CrossRef]
- Sołek, P.; Mytych, J.; Tabęcka-Łonczyńska, A.; Koziorowski, M. Molecular Consequences of Depression Treatment: A Potential In vitro Mechanism for Antidepressants-Induced Reprotoxic Side Effects. Int. J. Mol. Sci. 2021, 22, 11855. [Google Scholar] [CrossRef]
- Wajda, A.; Łapczuk, J.; Grabowska, M.; Pius-Sadowska, E.; Słojewski, M.; Laszczynska, M.; Urasinska, E.; Machalinski, B.; Drozdzik, M. Cell and region specificity of Aryl hydrocarbon Receptor (AhR) system in the testis and the epididymis. Reprod. Toxicol. 2017, 69, 286–296. [Google Scholar] [CrossRef]
- Qu, G.B.; Shi, J.B.; Li, Z.N.; Ruan, T.; Fu, J.J.; Wang, P.; Wang, T.; Jiang, G. Bin Detection of tris-(2, 3-dibromopropyl) isocyanurate as a neuronal toxicant in environmental samples using neuronal toxicity-directed analysis. Sci. China Chem. 2011, 54, 1651–1658. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Skóra, B.; Mańdziuk, M. Tris (2,3-Dibromopropyl) Isocyanurate (TDBP-TAZTO or TBC) Shows Different Toxicity Depending on the Degree of Differentiation of the Human Neuroblastoma (SH-SY5Y) Cell Line. Neurotox. Res. 2021, 39, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Ruan, T.; Wang, Y.; Wang, C.; Wang, P.; Fu, J.; Yin, Y.; Qu, G.; Wang, T.; Jiang, G. Identification and Evaluation of a Novel Heterocyclic Brominated Flame Retardant Tris(2,3-dibromopropyl) Isocyanurate in Environmental Matrices near a Manufacturing Plant in Southern China. Environ. Sci. Technol. 2009, 43, 3080–3086. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Hu, Z.; Wang, H.; Zhu, H.; Dong, Z.; Jiang, W.; Zhao, H.; Li, N.; Mi, W.; Wang, W.; et al. Tris-(2,3-Dibromopropyl) Isocyanurate, a New Emerging Pollutant, Impairs Cognition and Provokes Depression-Like Behaviors in Adult Rats. PLoS ONE 2015, 10, e0140281. [Google Scholar] [CrossRef] [PubMed]
- Sobecki, M.; Mrouj, K.; Camasses, A.; Parisis, N.; Nicolas, E.; Llères, D.; Gerbe, F.; Prieto, S.; Krasinska, L.; David, A.; et al. The cell proliferation antigen Ki-67 organises heterochromatin. Elife 2016, 5, e13722. [Google Scholar] [CrossRef]
- Cuylen, S.; Blaukopf, C.; Politi, A.Z.; Müller-Reichert, T.; Neumann, B.; Poser, I.; Ellenberg, J.; Hyman, A.A.; Gerlich, D.W. Ki-67 acts as a biological surfactant to disperse mitotic chromosomes. Nature 2016, 535, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Wit, N.; Buoninfante, O.A.; van den Berk, P.C.M.; Jansen, J.G.; Hogenbirk, M.A.; de Wind, N.; Jacobs, H. Roles of PCNA ubiquitination and TLS polymerases κ and η in the bypass of methyl methanesulfonate-induced DNA damage. Nucleic Acids Res. 2015, 43, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Boehm, E.M.; Powers, K.T.; Kondratick, C.M.; Spies, M.; Houtman, J.C.D.; Washington, M.T. The Proliferating Cell Nuclear Antigen (PCNA)-interacting Protein (PIP) Motif of DNA Polymerase η Mediates Its Interaction with the C-terminal Domain of Rev1. J. Biol. Chem. 2016, 291, 8735–8744. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, R.; Balla, M.; Lavranos, G.; Chalikias, M.; Kitsos, C.; Baka, S.; Kittas, C. Evaluation of immunohistochemical markers of germ cells’ proliferation in the developing rat testis: A comparative study. Tissue Cell 2008, 40, 43–50. [Google Scholar] [CrossRef]
- Steger, K. The proliferation of spermatogonia in normal and pathological human seminiferous epithelium: An immunohistochemical study using monoclonal antibodies against Ki-67 protein and proliferating cell nuclear antigen. Mol. Hum. Reprod. 1998, 4, 227–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrobel, K.-H.; Bickel, D.; Kujat, R. Immunohistochemical study of seminiferous epithelium in adult bovine testis using monoclonal antibodies against Ki-67 protein and proliferating cell nuclear antigen (PCNA). Cell Tissue Res. 1996, 283, 191–201. [Google Scholar] [CrossRef]
- Gerdes, J.; Schwab, U.; Lemke, H.; Stein, H. Production of a mouse monoclonal antibody reactive with a human nuclear antigen associated with cell proliferation. Int. J. Cancer 1983, 31, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Adeoya-Osiguwa, S.A.; Markoulaki, S.; Pocock, V.; Milligan, S.R.; Fraser, L.R. 17 -Estradiol and environmental estrogens significantly affect mammalian sperm function. Hum. Reprod. 2003, 18, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, R.; Krishna, A. Effect of Letrozole, a selective aromatase inhibitor, on testicular activities in adult mice: Both in vivo and in vitro study. Gen. Comp. Endocrinol. 2017, 241, 57–68. [Google Scholar] [CrossRef]
- Sipahutar, H.; Sourdaine, P.; Moslemi, S.; Plainfossé, B.; Séralini, G.-E. Immunolocalization of Aromatase in Stallion Leydig Cells and Seminiferous Tubules. J. Histochem. Cytochem. 2003, 51, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Benachour, N.; Moslemi, S.; Sipahutar, H.; Seralini, G. Cytotoxic effects and aromatase inhibition by xenobiotic endocrine disrupters alone and in combination☆. Toxicol. Appl. Pharmacol. 2007, 222, 129–140. [Google Scholar] [CrossRef]
- Belcher, S.M.; Cookman, C.J.; Patisaul, H.B.; Stapleton, H.M. In vitro assessment of human nuclear hormone receptor activity and cytotoxicity of the flame retardant mixture FM 550 and its triarylphosphate and brominated components. Toxicol. Lett. 2014, 228, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Gorga, A.; Rindone, G.M.; Regueira, M.; Pellizzari, E.H.; Camberos, M.C.; Cigorraga, S.B.; Riera, M.F.; Galardo, M.N.; Meroni, S.B. PPARγ activation regulates lipid droplet formation and lactate production in rat Sertoli cells. Cell Tissue Res. 2017, 369, 611–624. [Google Scholar] [CrossRef]
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef]
- Regueira, M.; Riera, M.F.; Galardo, M.N.; Pellizzari, E.H.; Cigorraga, S.B.; Meroni, S.B. Activation of PPAR α and PPAR β/δ regulates Sertoli cell metabolism. Mol. Cell. Endocrinol. 2014, 382, 271–281. [Google Scholar] [CrossRef]
- Górowska-Wójtowicz, E.; Dutka, P.; Kudrycka, M.; Pawlicki, P.; Milon, A.; Plachno, B.J.; Tworzydlo, W.; Pardyak, L.; Kaminska, A.; Hejmej, A.; et al. Regulation of steroidogenic function of mouse Leydig cells: G-coupled membrane estrogen receptor and peroxisome proliferator-activated receptor partnership. J. Physiol. Pharmacol. 2018, 69, 373–390. [Google Scholar]
- Martinez-Arguelles, D.B.; Campioli, E.; Culty, M.; Zirkin, B.R.; Papadopoulos, V. Fetal origin of endocrine dysfunction in the adult: The phthalate model. J. Steroid Biochem. Mol. Biol. 2013, 137, 5–17. [Google Scholar] [CrossRef]
- Hamers, T.; Kamstra, J.H.; Sonneveld, E.; Murk, A.J.; Kester, M.H.A.; Andersson, P.L.; Legler, J.; Brouwer, A. In vitro Profiling of the Endocrine-Disrupting Potency of Brominated Flame Retardants. Toxicol. Sci. 2006, 92, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Riu, A.; le Maire, A.; Grimaldi, M.; Audebert, M.; Hillenweck, A.; Bourguet, W.; Balaguer, P.; Zalko, D. Characterization of novel ligands of ERα, Erβ, and PPARγ: The case of halogenated bisphenol A and their conjugated metabolites. Toxicol. Sci. 2011, 122, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Riu, A.; Grimaldi, M.; le Maire, A.; Bey, G.; Phillips, K.; Boulahtouf, A.; Perdu, E.; Zalko, D.; Bourguet, W.; Balaguer, P. Peroxisome Proliferator-Activated Receptor γ Is a Target for Halogenated Analogs of Bisphenol A. Environ. Health Perspect. 2011, 119, 1227–1232. [Google Scholar] [CrossRef]
- Dunning, K.R.; Anastasi, M.R.; Zhang, V.J.; Russell, D.L.; Robker, R.L. Regulation of Fatty Acid Oxidation in Mouse Cumulus-Oocyte Complexes during Maturation and Modulation by PPAR Agonists. PLoS ONE 2014, 9, e87327. [Google Scholar] [CrossRef] [Green Version]
- Eslami, H.; Sharifi, A.M.; Rahimi, H.; Rahati, M. Protective effect of telmisartan against oxidative damage induced by high glucose in neuronal PC12 cell. Neurosci. Lett. 2014, 558, 31–36. [Google Scholar] [CrossRef]
- Zarzuelo, M.J.; López-Sepúlveda, R.; Sánchez, M.; Romero, M.; Gómez-Guzmán, M.; Ungvary, Z.; Pérez-Vizcaíno, F.; Jiménez, R.; Duarte, J. SIRT1 inhibits NADPH oxidase activation and protects endothelial function in the rat aorta: Implications for vascular aging. Biochem. Pharmacol. 2013, 85, 1288–1296. [Google Scholar] [CrossRef]
- El-Sayed, K.; Ali, D.A.; Maher, S.A.; Ghareeb, D.; Selim, S.; Albogami, S.; Fayad, E.; Kolieb, E. Prophylactic and Ameliorative Effects of PPAR-γ Agonist Pioglitazone in Improving Oxidative Stress, Germ Cell Apoptosis and Inflammation in Gentamycin-Induced Testicular Damage in Adult Male Albino Rats. Antioxidants 2022, 11, 191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, X.; Li, S.; Zhao, Z.; Li, J.; Cui, M.; Wang, E. Repression of ESR1 transcription by MYOD potentiates letrozole-resistance in ERα-positive breast cancer cells. Biochem. Biophys. Res. Commun. 2017, 492, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Harnchoowong, S.; Suchonwanit, P. PPAR- γ Agonists and Their Role in Primary Cicatricial Alopecia. PPAR Res. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wójtowicz, A.K.; Szychowski, K.A.; Wnuk, A.; Kajta, M. Dibutyl Phthalate (DBP)-Induced Apoptosis and Neurotoxicity are Mediated via the Aryl Hydrocarbon Receptor (AhR) but not by Estrogen Receptor Alpha (ERα), Estrogen Receptor Beta (ERβ), or Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) in Mouse C. Neurotox. Res. 2017, 31, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Chu, R.; van Hasselt, A.; Vlantis, A.C.; Ng, E.K.W.; Liu, S.Y.W.; Fan, M.D.; Ng, S.K.; Chan, A.B.W.; Liu, Z.; Li, X.; et al. The cross-talk between estrogen receptor and peroxisome proliferator-activated receptor gamma in thyroid cancer. Cancer 2014, 120, 142–153. [Google Scholar] [CrossRef]
- Rezvanfar, M.; Rezvanfar, M.; Ahmadi, A.; Saadi, H.S.; Baeeri, M.; Abdollahi, M. Mechanistic links between oxidative/nitrosative stress and tumor necrosis factor alpha in letrozole-induced murine polycystic ovary: Biochemical and pathological evidences for beneficial effect of pioglitazone. Hum. Exp. Toxicol. 2012, 31, 887–897. [Google Scholar] [CrossRef]
- Ohbayashi, T.; Oikawa, K.; Iwata, R.; Kameta, A.; Evine, K.; Isobe, T.; Matsuda, Y.; Mimura, J.; Fujii-Kuriyama, Y.; Kuroda, M.; et al. Dioxin induces a novel nuclear factor, DIF-3, that is implicated in spermatogenesis. FEBS Lett. 2001, 508, 341–344. [Google Scholar] [CrossRef] [Green Version]
- Schultz, R.; Suominen, J.; Vaärre, T.; Hakovirta, H.; Parvinen, M.; Toppari, J.; Pelto-Huikko, M. Expression of Aryl Hydrocarbon Receptor and Aryl Hydrocarbon Receptor Nuclear Translocator Messenger Ribonucleic Acids and Proteins in Rat and Human Testis. Endocrinology 2003, 144, 767–776. [Google Scholar] [CrossRef]
- Mitchell, K.A.; Elferink, C.J. Timing is everything: Consequences of transient and sustained AhR activity. Biochem. Pharmacol. 2009, 77, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.R.; Sandberg, R.; Lendahl, U. Notch signaling: Simplicity in design, versatility in function. Development 2011, 138, 3593–3612. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Butler, R.; Miao, Y.; Dai, Y.; Wu, W.; Su, W.; Fujii-Kuriyama, Y.; Warner, M.; Gustafsson, J.-Å. Dysregulation of Notch and ERα signaling in AhR −/− male mice. Proc. Natl. Acad. Sci. USA 2016, 113, 11883–11888. [Google Scholar] [CrossRef] [Green Version]
- Rampersad, S.N. Multiple Applications of Alamar Blue as an Indicator of Metabolic Function and Cellular Health in Cell Viability Bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Nicholson, D.W. Caspase structure, proteolytic substrates, and function during apoptotic cell death. Cell Death Differ. 1999, 6, 1028–1042. [Google Scholar] [CrossRef] [Green Version]
- Szychowski, K.A.; Wnuk, A.; Rzemieniec, J.; Kajta, M.; Leszczyńska, T.; Wójtowicz, A.K. Triclosan-Evoked Neurotoxicity Involves NMDAR Subunits with the Specific Role of GluN2A in Caspase-3-Dependent Apoptosis. Mol. Neurobiol. 2019, 56, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Skóra, B.; Szychowski, K.A. Molecular mechanism of the uptake and toxicity of EGF-LipoAgNPs in EGFR-overexpressing cancer cells. Biomed. Pharmacother. 2022, 150, 113085. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabęcka-Łonczyńska, A.; Skóra, B.; Kaleniuk, E.; Szychowski, K.A. Reprotoxic Effect of Tris(2,3-Dibromopropyl) Isocyanurate (TBC) on Spermatogenic Cells In Vitro. Molecules 2023, 28, 2337. https://doi.org/10.3390/molecules28052337
Tabęcka-Łonczyńska A, Skóra B, Kaleniuk E, Szychowski KA. Reprotoxic Effect of Tris(2,3-Dibromopropyl) Isocyanurate (TBC) on Spermatogenic Cells In Vitro. Molecules. 2023; 28(5):2337. https://doi.org/10.3390/molecules28052337
Chicago/Turabian StyleTabęcka-Łonczyńska, Anna, Bartosz Skóra, Edyta Kaleniuk, and Konrad A. Szychowski. 2023. "Reprotoxic Effect of Tris(2,3-Dibromopropyl) Isocyanurate (TBC) on Spermatogenic Cells In Vitro" Molecules 28, no. 5: 2337. https://doi.org/10.3390/molecules28052337
APA StyleTabęcka-Łonczyńska, A., Skóra, B., Kaleniuk, E., & Szychowski, K. A. (2023). Reprotoxic Effect of Tris(2,3-Dibromopropyl) Isocyanurate (TBC) on Spermatogenic Cells In Vitro. Molecules, 28(5), 2337. https://doi.org/10.3390/molecules28052337