


In Silico Exploration of Microtubule Agent Griseofulvin and Its Derivatives Interactions with Different Human β-Tubulin Isotypes

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Virtual Screening and Molecular Docking Studies
2.2. Molecular Dynamic (MD) Simulations
Molecular Mechanics Poisson-Boltzmann Surface Area (MM-PBSA)
2.3. Tubulin Isotype Expression in Cancer
3. Computational Methodology
3.1. Ligand and Receptor Preparation
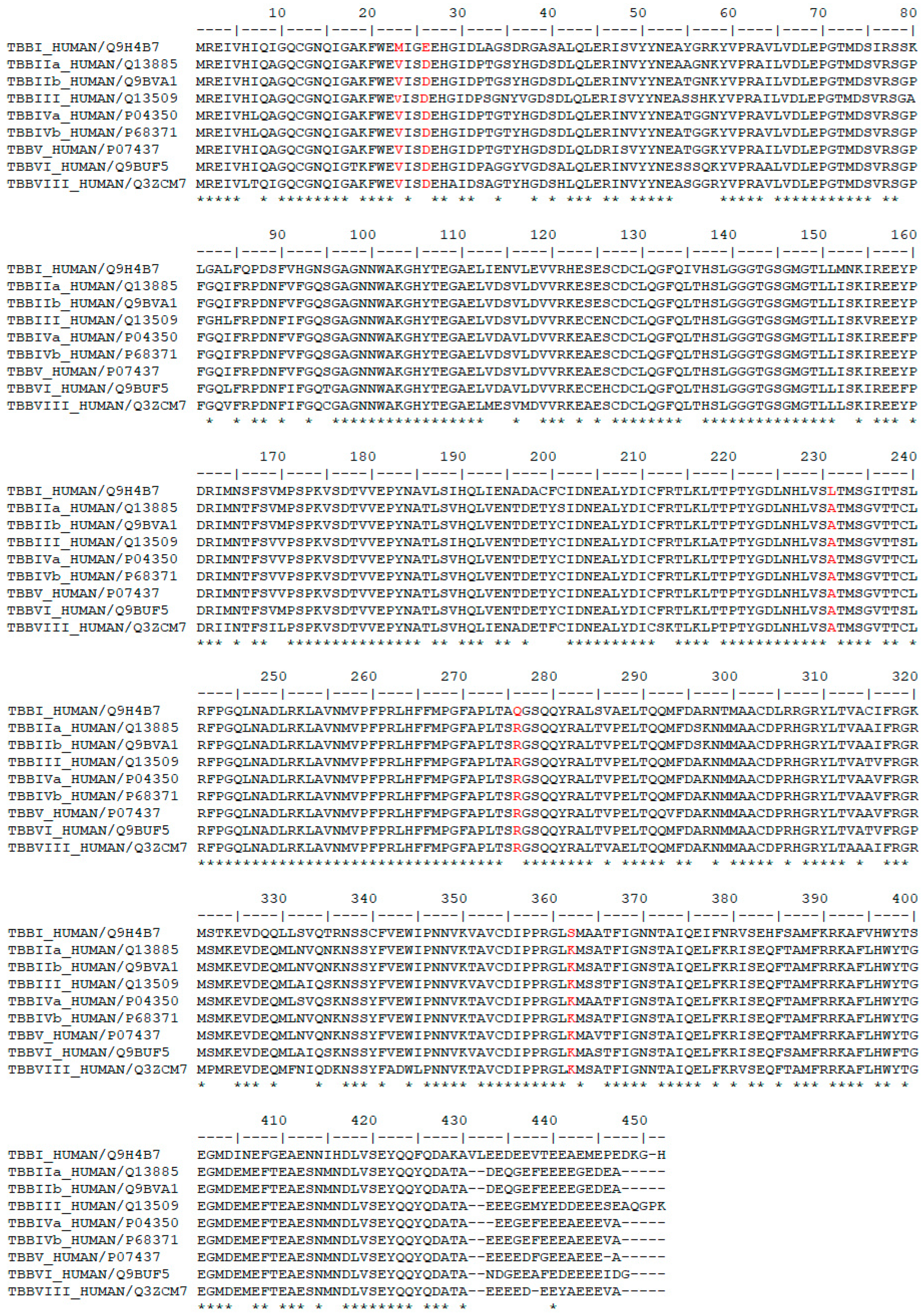
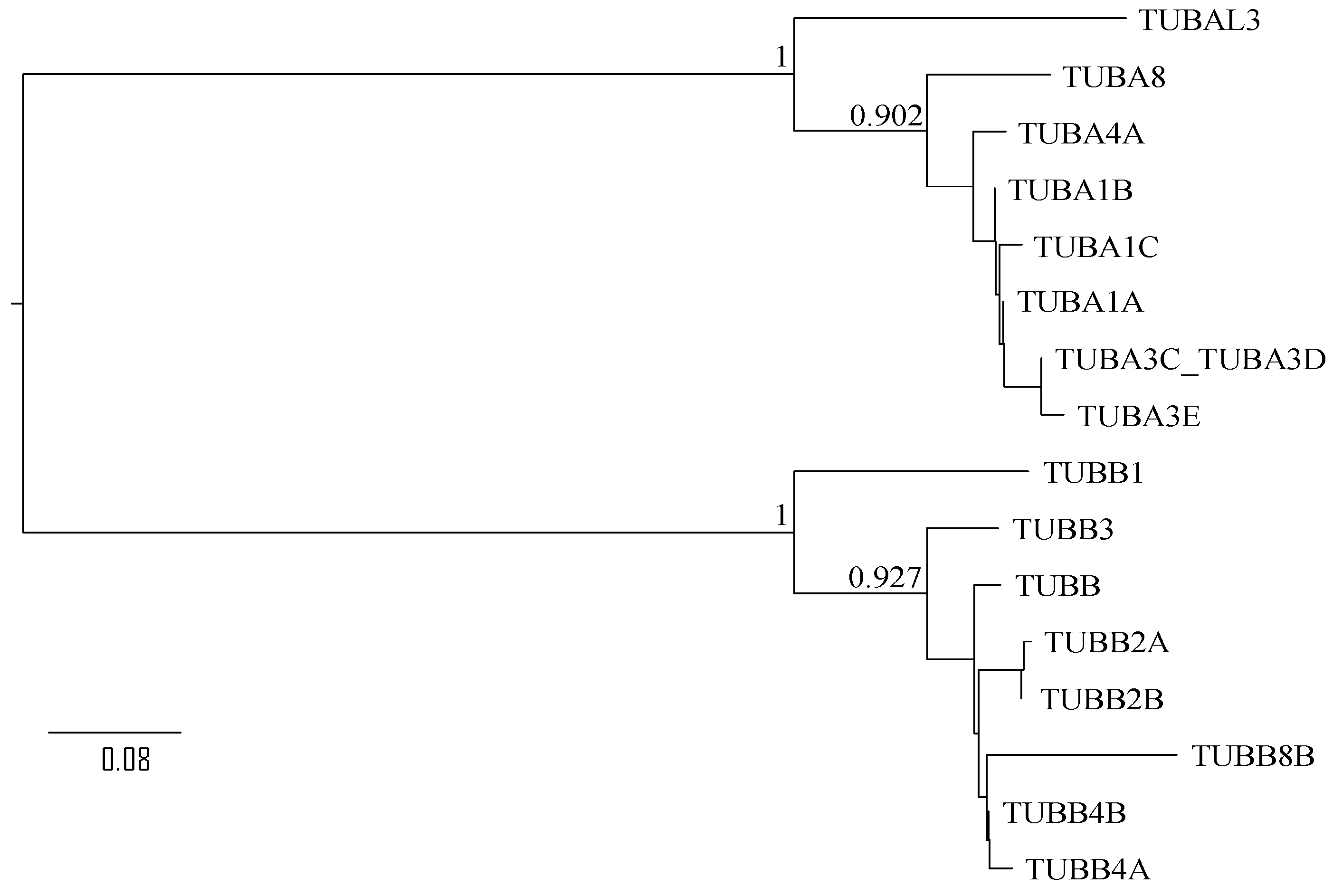
3.2. Sequence Analysis of β-Tubulin Isotypes
3.3. Virtual Screening and In Silico Molecular Docking
3.4. Molecular Dynamics Simulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oxford, A.E.; Raistrick, H.; Simonart, P. Studies in the Biochemistry of Micro-Organisms: Griseofulvin, C17h17o6cl, a Metabolic Product of Penicillium griseo-fulvum Dierckx. Biochem. J. 1939, 33, 240. [Google Scholar] [CrossRef] [PubMed]
- Lambert, D.R.; Siegle, R.J.; Camisa, C. Griseofulvin and Ketoconazole in the Treatment of Dermatophyte Infections. Int. J. Dermatol. 1989, 28, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Cacho, R.; Tang, Y. Identification of the Viridicatumtoxin and Griseofulvin Gene Clusters from Penicillium aethiopicum. Chem. Biol. 2010, 17, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.B.; Mun, H.Y.; Nguyen, T.T.T.; Kim, J.-C.; Stone, J.K. Abieticola koreana Gen. Et Sp. Nov., a Griseofulvin-Producing Endophytic Xylariaceous ascomycete from Korea. Mycotaxon 2016, 131, 749–764. [Google Scholar] [CrossRef]
- Ribeiro, A.I.; Costa, E.S.; Thomasi, S.S.; Brandão, D.F.R.; Vieira, P.C.; Fernandes, J.B.; Rossi Forim, M.; Ferreira, A.G.; Florentino Pascholati, S.; Pascholati Gusmão, L.F. Biological and Chemical Control of Sclerotinia Sclerotiorum Using Stachybotrys Levispora and Its Secondary Metabolite Griseofulvin. J. Agric. Food Chem. 2018, 66, 7627–7632. [Google Scholar] [CrossRef] [PubMed]
- Mead, M.E.; Raja, H.A.; Steenwyk, J.L.; Knowles, S.L.; Oberlies, N.H.; Rokas, A. Draft Genome Sequence of the Griseofulvin-Producing Fungus Xylaria flabelliformis Strain G536. Microbiol. Resour. Announc. 2019, 8, e00890-19. [Google Scholar] [CrossRef] [Green Version]
- Aris, P.; Yan, L.; Wei, Y.; Chang, Y.; Shi, B.; Xia, X. Conservation of Griseofulvin Genes in the Gsf Gene Cluster among Fungal Genomes. G3 2021, 12, jkab399. [Google Scholar] [CrossRef]
- Rebacz, B.; Larsen, T.O.; Clausen, M.H.; Rønnest, M.H.; Löffler, H.; Ho, A.D.; Krämer, A. Identification of Griseofulvin as an Inhibitor of Centrosomal Clustering in a Phenotype-Based Screen. Cancer Res. 2007, 67, 6342–6350. [Google Scholar] [CrossRef] [Green Version]
- Rathinasamy, K.; Jindal, B.; Asthana, J.; Singh, P.; Balaji, P.V.; Panda, D. Griseofulvin Stabilizes Microtubule Dynamics, Activates P53 and Inhibits the Proliferation of Mcf-7 Cells Synergistically with Vinblastine. BMC Cancer 2010, 10, 213. [Google Scholar] [CrossRef] [Green Version]
- Aris, P.; Wei, Y.; Mohamadzadeh, M.; Xia, X. Griseofulvin: An Updated Overview of Old and Current Knowledge. Molecules 2022, 27, 7034. [Google Scholar] [CrossRef]
- Knasmüller, S.; Parzefall, W.; Helma, C.; Kassie, F.; Ecker, S.; Schulte-Hermann, R. Toxic Effects of Griseofulvin: Disease Models, Mechanisms, and Risk Assessment. Crit. Rev. Toxicol. 1997, 27, 495–537. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, H.J. Hepatotoxicity: The Adverse Effects of Drugs and Other Chemicals on the Liver; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 1999. [Google Scholar]
- Moseley, R.H. Hepatotoxicity of Antimicrobials and Antifungal Agents. In Drug-Induced Liver Disease; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Jin, H.; Yamashita, A.; Maekawa, S.; Yang, P.; He, L.; Takayanagi, S.; Wakita, T.; Sakamoto, N.; Enomoto, N.; Ito, M. Griseofulvin, an Oral Antifungal Agent, Suppresses Hepatitis C Virus Replication in Vitro. Hepatol. Res. 2008, 38, 909–918. [Google Scholar]
- Aris, P.; Mohamadzadeh, M.; Wei, Y.; Xia, X. In Silico Molecular Dynamics of Griseofulvin and Its Derivatives Revealed Potential Therapeutic Applications for COVID-19. Int. J. Mol. Sci. 2022, 23, 6889. [Google Scholar] [CrossRef]
- Perez, E.A. Microtubule Inhibitors: Differentiating Tubulin-Inhibiting Agents Based on Mechanisms of Action, Clinical Activity, and Resistance. Mol. Cancer Ther. 2009, 8, 2086–2095. [Google Scholar] [CrossRef] [Green Version]
- Morris, P.G.; Fornier, M.N. Microtubule Active Agents: Beyond the Taxane Frontier. Clin. Cancer Res. 2008, 14, 7167–7172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdier-Pinard, P.; Pasquier, E.; Xiao, H.; Burd, B.; Villard, C.; Lafitte, D.; Miller, L.M.; Angeletti, R.H.; Horwitz, S.B.; Braguer, D. Tubulin Proteomics: Towards Breaking the Code. Anal. Biochem. 2009, 384, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janke, C. The Tubulin Code: Molecular Components, Readout Mechanisms, and Functions. J. Cell Biol. 2014, 206, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Nogales, E.; Wolf, S.G.; Downing, K.H. Erratum: Structure of the Aβ Tubulin Dimer by Electron Crystallography. Nature 1998, 393, 191. [Google Scholar] [CrossRef] [Green Version]
- Vilmar, A.; Garcia-Foncillas, J.; Huarriz, M.; Santoni-Rugiu, E.; Sorensen, J.B. Rt-Pcr Versus Immunohistochemistry for Correlation and Quantification of Ercc1, Brca1, Tubb3 and Rrm1 in Nsclc. Lung Cancer 2012, 75, 306–312. [Google Scholar] [CrossRef]
- Hasegawa, S.; Miyoshi, Y.; Egawa, C.; Ishitobi, M.; Taguchi, T.; Tamaki, Y.; Monden, M.; Noguchi, S. Prediction of Response to Docetaxel by Quantitative Analysis of Class I and Iii Β-Tubulin Isotype Mrna Expression in Human Breast Cancers. Clin. Cancer Res. 2003, 9, 2992–2997. [Google Scholar]
- Roque, D.M.; Bellone, S.; English, D.P.; Buza, N.; Cocco, E.; Gasparrini, S.; Bortolomai, I.; Ratner, E.; Silasi, D.-A.; Azodi, M.; et al. Tubulin-Β-Iii Overexpression by Uterine Serous Carcinomas Is a Marker for Poor Overall Survival after Platinum/Taxane Chemotherapy and Sensitivity to Epothilones. Cancer 2013, 119, 2582–2592. [Google Scholar] [CrossRef] [Green Version]
- Derry, W.B.; Wilson, L.; Khan, I.A.; Ludueña, R.F.; Jordan, M.A. Taxol Differentially Modulates the Dynamics of Microtubules Assembled from Unfractionated and Purified Β-Tubulin Isotypes. Biochemistry 1997, 36, 3554–3562. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Luduena, R.F. Kinetics of Colchicine Binding to Purified Beta-Tubulin Isotypes from Bovine Brain. J. Biol. Chem. 1992, 267, 13335–13339. [Google Scholar] [CrossRef]
- Xu, K.; Schwarz, P.M.; Ludueña, R.F. Interaction of Nocodazole with Tubulin Isotypes. Drug Dev. Res. 2002, 55, 91–96. [Google Scholar] [CrossRef]
- Crounse, R.G. Effective Use of Griseofulvin. Arch. Dermatol. 1963, 87, 176–178. [Google Scholar] [CrossRef]
- Liéby-Muller, F.; Le Baliner, Q.H.; Grisoni, S.; Fournier, E.; Guilbaud, N.; Marion, F. Synthesis and Activities towards Resistant Cancer Cells of Sulfone and Sulfoxide Griseofulvin Derivatives. Bioorganic Med. Chem. Lett. 2015, 25, 2078–2081. [Google Scholar] [CrossRef] [PubMed]
- Rønnest, M.H.; Rebacz, B.; Markworth, L.; Terp, A.H.; Larsen, T.O.; Krämer, A.; Clausen, M.H. Synthesis and Structure−Activity Relationship of Griseofulvin Analogues as Inhibitors of Centrosomal Clustering in Cancer Cells. J. Med. Chem. 2009, 52, 3342–3347. [Google Scholar] [CrossRef] [PubMed]
- Kellenberger, E.; Rodrigo, J.; Muller, P.; Rognan, D. Comparative Evaluation of Eight Docking Tools for Docking and Virtual Screening Accuracy. Proteins: Struct. Funct. Bioinform. 2004, 57, 225–242. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. Autodock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, I.; Hirono, S.; Liu, Q.; Nakagome, I.; Matsushita, Y. Simple Method of Calculating Octanol/Water Partition Coefficient. Chem. Pharm. Bull. 1992, 40, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Devalapally, H.; Chakilam, A.; Amiji, M.M. Role of Nanotechnology in Pharmaceutical Product Development. J. Pharm. Sci. 2007, 96, 2547–2565. [Google Scholar] [CrossRef] [PubMed]
- Rønnest, M.H.; Raab, M.S.; Anderhub, S.; Boesen, S.; Krämer, A.; Larsen, T.O.; Clausen, M.H. Disparate Sar Data of Griseofulvin Analogues for the Dermatophytes Trichophyton Mentagrophytes, T. Rubrum, and Mda-Mb-231 Cancer Cells. J. Med. Chem. 2012, 55, 652–660. [Google Scholar] [CrossRef] [Green Version]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W.; et al. Calculating Structures and Free Energies of Complex Molecules: Combining Molecular Mechanics and Continuum Models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef]
- Kavallaris, M.; Kuo, D.Y.; Burkhart, C.A.; Regl, D.L.; Norris, M.; Haber, M.; Horwitz, S.B. Taxol-Resistant Epithelial Ovarian Tumors Are Associated with Altered Expression of Specific Beta-Tubulin Isotypes. J. Clin. Investig. 1997, 100, 1282–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiser, L.; Aggarwal, A.; Young, R.; Frankfurter, A.; Spano, A.; Correia, J.J.; Lobert, S. Comparison of Β-Tubulin Mrna and Protein Levels in 12 Human Cancer Cell Lines. Cell Motil. Cytoskelet. 2006, 63, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a Knowledge-Based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, Y.; Oda, Y.; Basaki, Y.; Kobayashi, H.; Wake, N.; Kuwano, M.; Tsuneyoshi, M. Expression of Beta-Tubulin Isotypes in Human Primary Ovarian Carcinoma. Gynecol. Oncol. 2007, 105, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Bernard-Marty, C.; Treilleux, I.; Dumontet, C.; Cardoso, F.; Fellous, A.; Gancberg, D.; Bissery, M.-C.; Paesmans, M.; Larsimont, D.; Piccart, M.J.; et al. Microtubule-Associated Parameters as Predictive Markers of Docetaxel Activity in Advanced Breast Cancer Patients: Results of a Pilot Study. Clin. Breast Cancer 2002, 3, 341–345. [Google Scholar] [CrossRef]
- Levallet, G.; Bergot, E.; Antoine, M.; Creveuil, C.; Santos, A.O.; Beau-Faller, M.; De Fraipont, F.; Brambilla, E.; Levallet, J.; Morin, F. High Tubb3 Expression, an Independent Prognostic Marker in Patients with Early Non–Small Cell Lung Cancer Treated by Preoperative Chemotherapy, Is Regulated by K-Ras Signaling Pathwayk-Ras and Tubb3 in Early Nsclc. Mol. Cancer Ther. 2012, 11, 1203–1213. [Google Scholar] [CrossRef] [Green Version]
- Aoki, D.; Oda, Y.; Hattori, S.; Taguchi, K.-I.; Ohishi, Y.; Basaki, Y.; Oie, S.; Suzuki, N.; Kono, S.; Tsuneyoshi, M.; et al. Overexpression of Class Iii Β-Tubulin Predicts Good Response to Taxane-Based Chemotherapy in Ovarian Clear Cell Adenocarcinoma. Clin. Cancer Res. 2009, 15, 1473–1480. [Google Scholar] [CrossRef] [Green Version]
- Tsourlakis, M.C.; Weigand, P.; Grupp, K.; Kluth, M.; Steurer, S.; Schlomm, T.; Graefen, M.; Huland, H.; Salomon, G.; Steuber, T. Βiii-Tubulin Overexpression Is an Independent Predictor of Prostate Cancer Progression Tightly Linked to Erg Fusion Status and Pten Deletion. Am. J. Pathol. 2014, 184, 609–617. [Google Scholar] [CrossRef]
- Christoph, D.C.; Kasper, S.; Gauler, T.C.; Loesch, C.; Engelhard, M.; Theegarten, D.; Poettgen, C.; Hepp, R.; Peglow, A.; Loewendick, H. Βv-Tubulin Expression Is Associated with Outcome Following Taxane-Based Chemotherapy in Non-Small Cell Lung Cancer. Br. J. Cancer 2012, 107, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eswar, N.; Eramian, D.; Webb, B.; Shen, M.Y.; Sali, A. Protein Structure Modeling with Modeller. In Structural Proteomics: High-throughput Methods; Humana Press: Totowa, NJ, USA, 2008; pp. 145–159. [Google Scholar]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. Procheck: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with Alphafold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Asimenos, G.; Toh, H.; Posada, D. Bioinformatics for DNA Sequence Analysis. Methods Mol. Biol. 2009, 537, 39–64. [Google Scholar]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with Pyrx. In Chemical Biology; Springer: Berlin, Germany, 2015; pp. 243–250. [Google Scholar]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and Autodocktools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Bauer, P.; Hess, B.; Lindahl, E. Gromacs 2022 Source Code. 2022. Available online: https://zenodo.org/record/6103835#.ZAHm33bMJPY (accessed on 10 January 2023).
- Sousa da Silva, A.W.; Vranken, W.F. Acpype-Antechamber Python Parser Interface. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Kumbhar, B.V.; Borogaon, A.; Panda, D.; Kunwar, A. Exploring the Origin of Differential Binding Affinities of Human Tubulin Isotypes Aβii, Aβiii and Aβiv for Dama-Colchicine Using Homology Modelling, Molecular Docking and Molecular Dynamics Simulations. PLoS ONE 2016, 11, e0156048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumbhar, B.V.; Panda, D.; Kunwar, A. Interaction of Microtubule Depolymerizing Agent Indanocine with Different Human Aβ Tubulin Isotypes. PLoS ONE 2018, 13, e0194934. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PubChem IDs | Structures | Binding Energies (kcal/mol) | MlogP |
|---|---|---|---|
| 441140 (griseofulvin) |  | −6.6 | 0.71 |
| CID 25171849 (G1) |  | −9.1 | 2.43 |
| CID 46844082 (G2) |  | −9.0 | 2.98 |
| CID 73331205 (G3) |  | −9.1 | 2.95 |
| CID 73331488 (G4) |  | −9.3 | 2.51 |
| CID 118263246 (G5) |  | −8.8 | 1.96 |
| AlphaFold ID | Taxol | Griseofulvin | 25171849 (G1) | 46844082 (G2) | 73331205 (G3) | 73331488 (G4) | 118263246 (G5) | |
|---|---|---|---|---|---|---|---|---|
| BI | Q9H4B7 | −6.29 | −6.8 | −8.7 | −7.29 | −8.09 | −7.36 | −7.95 |
| BIIa | Q13885 | −6.95 | −7.38 | −8.89 | −9.07 | −8.91 | −8.21 | −8.31 |
| BIIb | Q9BVA1 | −7.17 | −7.1 | −8.8 | −10.3 | −8.7 | −8.1 | −8.3 |
| BIII | Q13509 | −6.43 | −7.38 | −8.72 | −8.75 | −8.96 | −8.31 | −8.41 |
| BIVa | P04350 | −8.11 | −7.24 | −9.04 | −9.88 | −8.75 | −8.03 | −8.37 |
| BIVb | P68371 | −7.71 | −7.24 | −8.65 | −9.68 | −8.84 | −7.95 | −8.23 |
| BV | P07437 | −7.6 | −6.9 | −9.32 | −9.16 | −8.67 | −7.6 | −8.72 |
| BVI | Q9BUF5 | −7.66 | −7.24 | −8.65 | −9.63 | −9.11 | −8.1 | −8.41 |
| BVIII | Q3ZCM7 | −7.25 | −7.29 | −8.8 | −9.12 | −9.04 | −8.17 | −8.31 |
 | ||||||||
| β-Tubulin Isotype | Ligands | Binding Energy (kcal/mol) | H bonds | Hydrophobic Interaction |
|---|---|---|---|---|
| βI | Taxol | −6.29 | Lys19, Gln276 | Leu215, His227 |
| Griseofulvin | −6.8 | Gln276, Gln279 | Cys211, Leu215, Leu217, His227 | |
| G1 | −8.7 | Gln279 | Met23, Leu215, Leu217, His227, Leu231, Pro358, Leu361 | |
| G2 | −7.29 | Gln279 | Met23, Leu215, Leu217, Pro272, Thr274, Gln276, Gln279, Gly360, Leu261 | |
| G3 | −8.09 | Lys19 | Lys19, Met23, Leu215, Gly223, His227, Leu228, Leu231, Phe270, Leu273, | |
| G4 | −7.36 | Lys19 | Met23, Lys19, Leu215, Gly223, His227, Leu228, Leu231, Phe270, Pro272, Leu273 | |
| G5 | −7.95 | Gln276, Gln279 | Leu215, Leu217, His227, Thr274 | |
| βIIa | Taxol | −6.95 | Gln279, Arg359, Lys362 | Pro272, Gln280, Leu284, Leu361 |
| Griseofulvin | −7.38 | Thr274 | Leu215, His227, Arg359, Arg276, Gln279 | |
| G1 | −8.89 | Thr274 | Leu215, His227, Thr274, Arg276, Leu361 | |
| G2 | −9.07 | - | Val23, Leu215, Leu361, Lys362, Arg359 | |
| G3 | −8.91 | Thr274 | Leu215, His227, Arg276, Gly360 | |
| G4 | −8.21 | Thr274 | Leu215, His227, Arg276, Gly360 | |
| G5 | −8.31 | Thr274, Gln279 | Leu215, Arg276, Thr274, Gln279 | |
| βIIb | Taxol | −7.17 | His227, Gln279 | Val23, Leu215, Ala231, Pro272, Arg276, Gln279, Arg359, Leu361 |
| Griseofulvin | −7.1 | Thr274 | Leu215, His227, Gln279, Arg276 | |
| G1 | −8.8 | Thr274 | Leu215, His227, Arg276, Leu361 | |
| G2 | −10.3 | Thr274 | Leu215, His227, Thr274, Arg276, Gln279, Arg282, Leu284, Arg359, LeU361 | |
| G3 | −8.7 | Thr274 | Leu215, His227, Thr274, Arg276, Gly360 | |
| G4 | −8.1 | Thr274 | Leu215, His227, Thr274, Arg276, Gly360 | |
| G5 | −8.3 | Thr274, Gln279 | Leu215, His227, Thr274, Arg276 | |
| βIII | Taxol | −6.43 | His227, Arg276, Gly360 | Leu215, His227, Pro272, Arg276, Gln279, Gly360, Leu361 |
| Griseofulvin | −7.38 | Thr274 | Leu215, His227, Arg276, Gln279 | |
| G1 | −8.72 | - | Leu215, Thr274, His227, Leu273, Arg276, Gly360, Leu361 | |
| G2 | −8.75 | Thr274 | Asp26, Val23, Leu215, His227, Leu361, Leu273, Arg276, Arg359 | |
| G3 | −8.96 | Thr274 | Leu215, His227, Thr274, Arg276, Gly360, Leu361 | |
| G4 | −8.31 | Thr274 | Leu215, His227, Arg276, Gln279, Gly360 | |
| G5 | −8.41 | Thr274, Gln279 | Leu215, Leu217, Arg276, His227 | |
| βIVa | Taxol | −8.11 | Lys19, Thr274, Arg359 | Leu215, Leu217, His227, Leu228, Ala231, Phe270, Leu273, Pro358, Gly360, |
| Griseofulvin | −7.24 | Thr274 | Leu215, His227, Pro272, Arg276, Gln279, Leu361 | |
| G1 | −9.04 | - | Leu215, His227, Thr274, Arg276, Leu361 | |
| G2 | −9.88 | Thr274 | Leu15, Arg276, Leu284 | |
| G3 | −8.75 | Gln279 | Val23, Leu215, Gly223, Asp224, His227, Leu228, Ala231, Pro358, Arg359, Leu361 | |
| G4 | −8.03 | - | Leu215, His227, Arg276, Gly360 | |
| G5 | −8.37 | Thr274 | Leu215, Leu217, His227, Arg276, Thr274 | |
| βIVb | Taxol | −7.71 | Lys19, Thr274, Arg359 | Leu215, His227, Leu228, Ala231, Leu273, Leu300, Pro358 |
| Griseofulvin | −7.24 | Thr274 | Leu215, His227, Arg276, Gln279 | |
| G1 | −8.65 | Thr274 | Leu215, His227, Arg276, Leu361 | |
| G2 | −9.68 | Thr274 | Val23, Leu215, His227, Pro272, Arg276, Pro358, Arg359, Gly360, Leu361 | |
| G3 | −8.84 | Gln279 | Val23, Leu215, Gly223, Asp224, His227, Ala231, Pro358, Arg359, Leu361 | |
| G4 | −7.95 | Thr274 | Leu215, Leu217, His227, Thr274, Arg276, Gly360 | |
| G5 | −8.23 | Thr274 | Leu215, Leu217, His227, Thr274, Arg276 | |
| βV | Taxol | −7.6 | His227, Arg276, Gln279, Arg359 | Val23, Leu215, His227, Ala231, Arg359 |
| Griseofulvin | −6.9 | Thr274 | Leu215, His227, Thr274, Arg276, Gln279, Arg359 | |
| G1 | −9.32 | Ser230 | Lys19, Val23, His227, Ala231, Pro272, Arg359, Leu361 | |
| G2 | −9.16 | Gln279 | Val23, Leu215, Ala231, Phe270, Leu273, Thr274, Arg276, Gln279, Arg359, Leu361, | |
| G3 | −8.67 | His227 | Lys19, Val23, Leu215, His227, Leu228, Ala231, Arg359, Leu273 | |
| G4 | −7.6 | His227, Ser230 | LyS19, Val23, Leu215, His227, Leu228, Ala231, Leu273, Arg359 | |
| G5 | −8.72 | Ser230 | Lys19, Val23, His227. Pro272, Arg259, Arg359, Leu361, | |
| βVI | Taxol | −7.66 | Lys19, Arg276, Arg359 | Val23, Asp224, His227, Ala231, Phe270, Arg359 |
| Griseofulvin | −7.24 | Thr274 | Leu215, His227, Arg276, Gln279 | |
| G1 | −8.65 | Arg276, Gln279 | Val23, Asp224, His227, Ala231, Pro358, Arg359, Leu361 | |
| G2 | −9.63 | - | Lys19, Glu22, Val23, Asp26, Leu215, Leu217, His227, Arg359, Leu273, Arg276, Gln279, Leu361 | |
| G3 | −9.11 | Gln279 | Val23, Cys211, Leu215, Gly223, Asp224, His227, Ala231, Pro358, Arg359, Leu361 | |
| G4 | −8.1 | Gln279, Arg359 | Val23, Glu27, Leu215, Asp224, His227, Leu228, Ala231, Pro358, Arg359, Leu361 | |
| G5 | −8.41 | Thr274 | Leu215, His227, Thr274, Arg276 | |
| βVIII | Taxol | −7.25 | Arg276, Arg359 | Lys19, Glu22, Val23, Asp26, His227, Phe270, Arg359 |
| Griseofulvin | −7.29 | Thr274 | Leu215, Arg276, His227, Thr274, Arg276, Arg359, | |
| G1 | −8.8 | Thr274 | Leu215, His227, Thr274, Arg276, Leu361 | |
| G2 | −9.12 | Thr274 | Leu215, His227, Arg276, Gln279, Leu284, | |
| G3 | −9.04 | Gln279 | Val23, LeU215, His227, Leu228, Ala231, Pro358, Arg359, Leu361 | |
| G4 | −8.17 | Thr274 | Leu215, His227, Thr274, Arg276, Gly360, Leu361 | |
| G5 | −8.31 | Thr274 | Leu215, His227, Thr274, Arg276 |
| Β-Tubulin Isotype | Binding Energy (kcal/mol) | 3D Interactions | 2D Interactions |
|---|---|---|---|
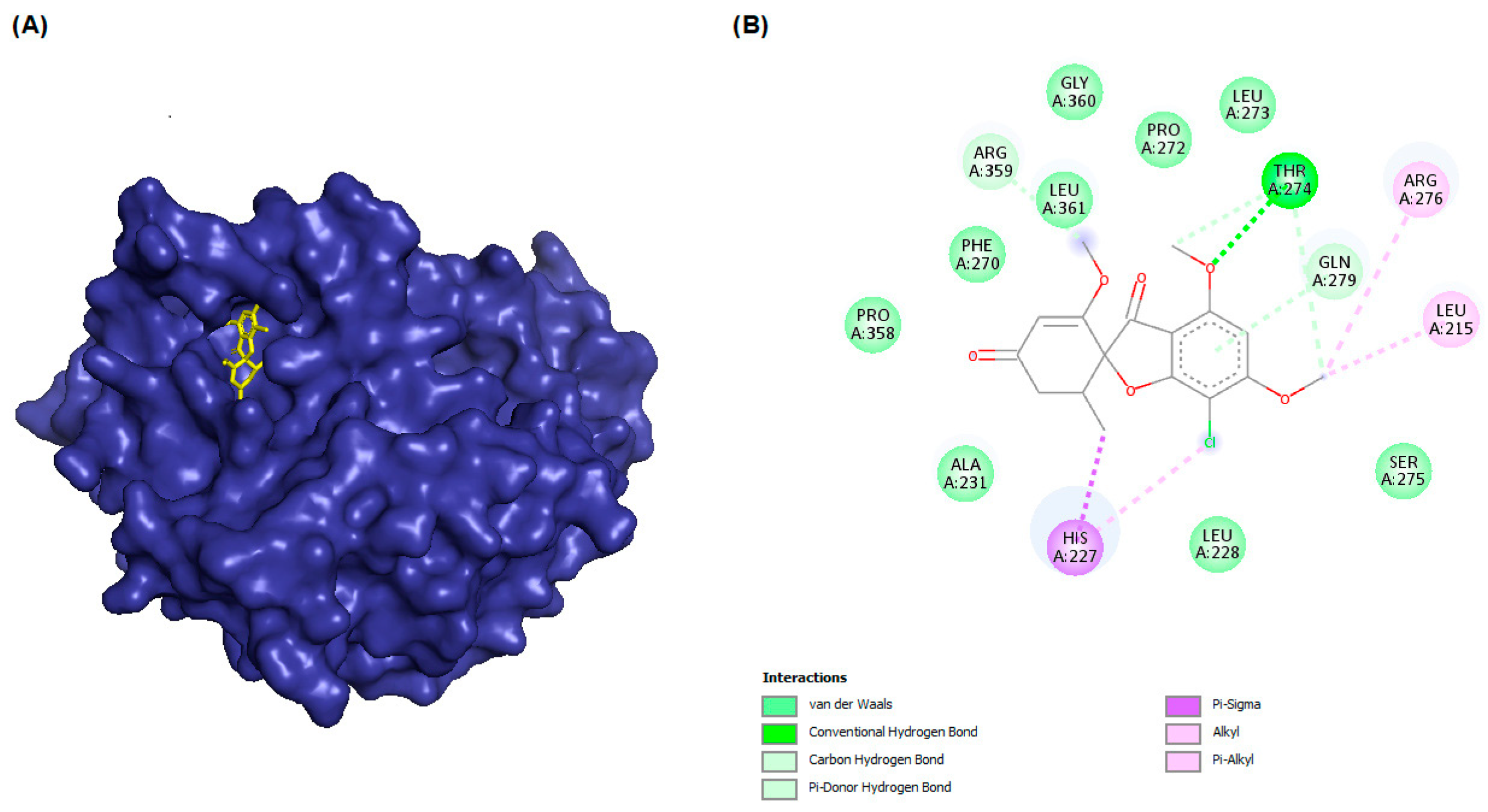
| βI | −8.7 |  |  |
| βIIa | −8.89 |  |  |
| βIIb | −8.8 |  |  |
| βIII | −8.72 |  |  |
| βIVa | −9.04 |  |  |
| βIVb | −8.65 |  |  |
| βV | −9.32 |  |  |
| βVI | −8.65 |  |  |
| βVIII | −8.8 |  |  |
| Complex | RMSD ± SD (Å) | Rg ± SD (Å) | RMSF ± SD (Å) | SASA ± SD (Å2) |
|---|---|---|---|---|
| P07437 | 0.68 ± 0.008 | 2.13 ± 0.04 | 0.096 ± 0.05 | 185.5 ± 2.26 |
| βI-G1 | 1.08 ± 0.08 | 2.15 ± 0.05 | 0.09 ± 0.05 | 188.1 ± 2.7 |
| βIIa-G1 | 0.29 ± 0.008 | 2.14 ± 0.05 | 0.096 ± 0.04 | 183.4 ± 2.2 |
| βIIb-G1 | 0.70 ± 0.02 | 2.14 ± 0.08 | 0.108 ± 0.101 | 190.3 ± 2.8 |
| βIII-G1 | 0.37 ± 0.04 | 2.15 ± 0.18 | 0.155 ± 0.201 | 200.1 ± 3.1 |
| βIVa-G1 | 0.32 ± 0.006 | 2.19 ± 0.05 | 0.093 ± 0.047 | 188.1 ± 2.5 |
| βIVb-G1 | 0.38 ± 0.022 | 2.15 ± 0.06 | 0.101 ± 0.074 | 188.7 ± 2.6 |
| βV-G1 | 0.41 ± 0.02 | 2.13 ± 0.09 | 0.11 ± 0.07 | 193.7 ± 2.7 |
| βVI-G1 | 0.43 ± 0.01 | 2.14 ± 0.04 | 0.097 ± 0.047 | 191.6 ± 2.5 |
| βVIII-G1 | 0.44 ± 0.01 | 2.13 ± 0. 04 | 0.094 ± 0.063 | 185.8 ± 2.3 |
| Complex | van der Waal Energy (kcal/mol) | Electrostatic Energy (kcal/mol) | Polar Solvation Energy (kcal/mol) | SASA Energy (kcal/mol) | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|
| βI-G1 | −40.0 ± 2.7 | −4.5 ± 1.4 | 21.1 ± 2.5 | −4.0 ± 0.3 | −34.6 ± 3.0 |
| βIIa-G1 | −41.0 ± 3.4 | −0.43 ± 0.1 | 19.9 ± 3.6 | −4.6 ± 0.3 | −25.3 ± 13.2 |
| βIIb-G1 | −40.1± 3.7 | −0.3 ± 0.1 | 18.6 ± 3.0 | −4.1 ± 0.3 | −25.9 ± 3.1 |
| βIII-G1 | −35.2 ± 7.6 | −0.7 ± 0.3 | 19.2 ± 3.7 | −3.8 ± 0.6 | −20.3 ± 6.5 |
| βIVa-G1 | −43.2± 3.1 | −4.3 ± 2.1 | 27.6 ± 4.4 | −4.4 ± 0.3 | −24.3 ± 3.0 |
| βIVb-G1 | −51.3 ± 3.0 | −3.4 ± 1.5 | 30.0 ± 3.6 | −5.1± 0.3 | −30.8 ± 3.6 |
| βV-G1 | −40.7 ± 6.1 | −0.9 ± 0.3 | 20.4 ± 4.1 | −4.2 ± 0.4 | −25.2 ± 4.4 |
| βVI-G1 | −40.5 ± 3.1 | −3.6 ± 1.4 | 23.3 ± 2.7 | −4.2 ± 0.3 | −25.0 ± 3.1 |
| βVIII-G1 | −35.9± 3.5 | −1.2 ± 0.5 | 18.3 ± 2.0 | −3.8 ± 0.3 | −22.5 ± 2.9 |
| β-Tubulin Isotype | Tissue Specificity | Single-Cell Type Expression Cluster | Cell-Line Enriched | Tumor Type | Cancer Prognostic and Effect | Reference |
|---|---|---|---|---|---|---|
| βI | Bone marrow, lymphoid tissue | Myeloid cells | HEL | Breast cancer | Poor response to docetaxel treatment | [22,38] |
| βIIa | Brain | Squamous epithelial cells, keratinization | SuSa | Breast and ovarian cancer | Prognostic marker in urothelial cancer (unfavorable) and renal cancer (favorable); poor response to taxane treatment or advanced stage disease | [38,39,40] |
| βIIb | Brain, choroid plexus | Muller glia cells, visual perception | SCLC-21H | Breast and ovarian cancer | Prognostic marker in endometrial cancer (unfavorable); poor response to taxane treatment | [38,39,40] |
| βIII | Brain, neuropeptide signaling | Pancreatic cells, digestion | SH-SY5Y | Breast, ovarian, prostate, uterine serous carcinoma cancer | Gene product is not prognostic; poor response to taxane treatment | [22,23,38,39,41,42,43] |
| βIVa | Brain, nervous system | Oligodendrocytes, myelin sheath organization | AN3-CA | Ovarian cancer | Prognostic marker in endometrial cancer (unfavorable); poor response to taxol treatment | [36,38] |
| βIVb | Ciliated cells, cilium, and cell projection | Mitochondria | MCF-7 | Ovarian cancer | Prognostic marker in thyroid cancer and endometrial cancer (favorable), and liver cancer (unfavorable) | [36,38] |
| βV | Lymphoid tissue, immune response regulation | Non-specific | WM-115 | Non-small cell lung cancer (NSCLC) | Prognostic marker in renal cancer and liver cancer (unfavorable); favorable response to taxane treatment | [38,44] |
| βVI | Fibroblasts, ECM organization | Extravillous trophoblasts | TIME | Not specified | Prognostic marker in renal cancer (unfavorable) and urothelial cancer (unfavorable) | [38] |
| βVIII | Testis | Spermatogonia and spermatocytes | MCF-7 | Not specified | Gene product is not prognostic | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aris, P.; Mohamadzadeh, M.; Kruglikov, A.; Askari Rad, M.; Xia, X. In Silico Exploration of Microtubule Agent Griseofulvin and Its Derivatives Interactions with Different Human β-Tubulin Isotypes. Molecules 2023, 28, 2384. https://doi.org/10.3390/molecules28052384
Aris P, Mohamadzadeh M, Kruglikov A, Askari Rad M, Xia X. In Silico Exploration of Microtubule Agent Griseofulvin and Its Derivatives Interactions with Different Human β-Tubulin Isotypes. Molecules. 2023; 28(5):2384. https://doi.org/10.3390/molecules28052384
Chicago/Turabian StyleAris, Parisa, Masoud Mohamadzadeh, Alibek Kruglikov, Mahbubeh Askari Rad, and Xuhua Xia. 2023. "In Silico Exploration of Microtubule Agent Griseofulvin and Its Derivatives Interactions with Different Human β-Tubulin Isotypes" Molecules 28, no. 5: 2384. https://doi.org/10.3390/molecules28052384
APA StyleAris, P., Mohamadzadeh, M., Kruglikov, A., Askari Rad, M., & Xia, X. (2023). In Silico Exploration of Microtubule Agent Griseofulvin and Its Derivatives Interactions with Different Human β-Tubulin Isotypes. Molecules, 28(5), 2384. https://doi.org/10.3390/molecules28052384