
Characterization of Beef Coming from Different European Countries through Stable Isotope (H, C, N, and S) Ratio Analysis

 ,
,  , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Carbon Stable Isotopic Ratio
Carbon Isotopic Ratio of the Beef Fat
2.2. Nitrogen Stable Isotopic Ratio
2.3. Sulphur Stable Isotopic Ratio
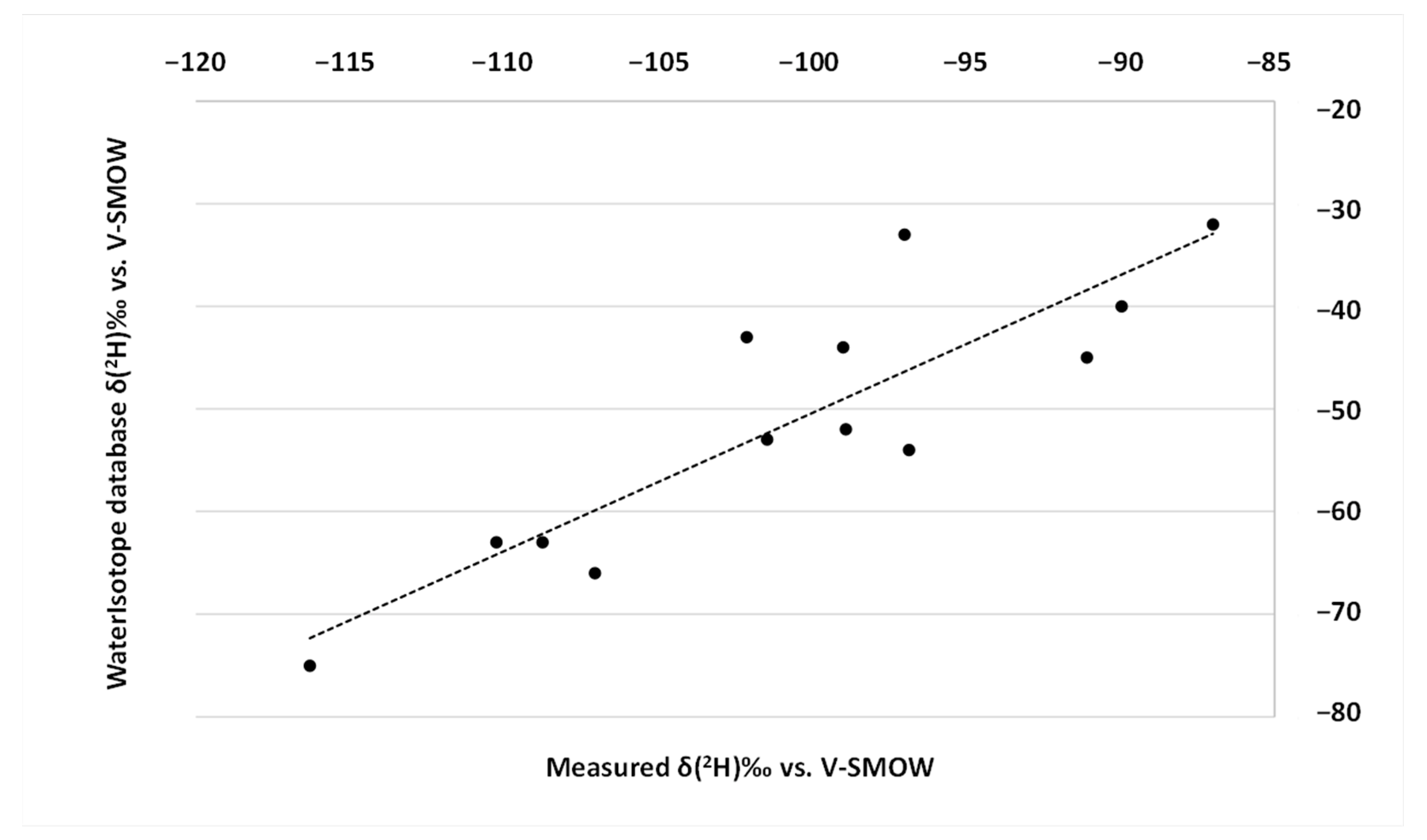
2.4. Hydrogen Stable Isotopic Ratio
2.5. Principal Component Analysis
3. Materials and Methods
3.1. Samples
3.2. Preparative and Analysis Procedures
3.3. Stable Isotope Ratio Analysis
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kendall, H.; Clark, B.; Rhymer, C.; Kuznesof, S.; Hajslova, J.; Tomaniova, M.; Brereton, P.; Frewer, L. A Systematic Review of Consumer Perceptions of Food Fraud and Authenticity: A European Perspective. Trends Food Sci. Technol. 2019, 94, 79–90. [Google Scholar] [CrossRef]
- Camin, F.; Dordevic, N.; Wehrens, R.; Neteler, M.; Delucchi, L.; Postma, G.; Buydens, L. Climatic and Geographical Dependence of the H, C and O Stable Isotope Ratios of Italian Wine. Anal. Chim. Acta 2015, 853, 384–390. [Google Scholar] [CrossRef]
- Horacek, M.; Ogrinc, N.; Magdas, D.A.; Wunderlin, D.; Sucur, S.; Maras, V.; Misurovic, A.; Eder, R.; Čuš, F.; Wyhlidal, S.; et al. Isotope Analysis (13C, 18O) of Wine From Central and Eastern Europe and Argentina, 2008 and 2009 Vintages: Differentiation of Origin, Environmental Indications, and Variations Within Countries. Front. Sustain. Food Syst. 2021, 5, 1–10. [Google Scholar] [CrossRef]
- Camin, F.; Boner, M.; Bontempo, L.; Fauhl-Hassek, C.; Kelly, S.D.; Riedl, J.; Rossmann, A. Stable Isotope Techniques for Verifying the Declared Geographical Origin of Food in Legal Cases. Trends Food Sci. Technol. 2017, 61, 176–187. [Google Scholar] [CrossRef]
- Rossmann, A. Determination of Stable Isotope Ratios in Food Analysis. Food Rev. Int. 2001, 17, 347–381. [Google Scholar] [CrossRef]
- Camin, F.; Bontempo, L.; Perini, M.; Piasentier, E. Stable Isotope Ratio Analysis for Assessing the Authenticity of Food of Animal Origin. Compr. Rev. Food Sci. Food Saf. 2016, 15, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Camin, F.; Bontempo, L.; Heinrich, K.; Horacek, M.; Kelly, S.D.; Schlicht, C.; Thomas, F.; Monahan, F.J.; Hoogewerff, J.; Rossmann, A. Multi-Element (H,C,N,S) Stable Isotope Characteristics of Lamb Meat from Different European Regions. Anal. Bioanal. Chem. 2007, 389, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perini, M.; Camin, F.; Bontempo, L.; Rossmann, A.; Piasentier, E. Multielement (H, C, N, O, S) Stable Isotope Characteristics of Lamb Meat from Different Italian Regions. Rapid Commun. Mass Spectrom. 2009, 23, 2573–2585. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.M. Poultry and Pork Muscle Defects and Meat Quality–Consequences, Causes, and Management. J. Anim. Sci. 2020, 98, skaa263. [Google Scholar] [CrossRef]
- Zhao, Y.; Tu, T.; Tang, X.; Zhao, S.; Qie, M.; Chen, A.; Yang, S. Authentication of Organic Pork and Identification of Geographical Origins of Pork in Four Regions of China by Combined Analysis of Stable Isotopes and Multi-Elements. Meat Sci. 2020, 165, 108129. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, B.; Guo, B.; Wang, D.; Yang, S. Combination of Multi-Element and Stable Isotope Analysis Improved the Traceability of Chicken from Four Provinces of China. CyTA J. Food 2016, 14, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Boner, M.; Förstel, H. Stable Isotope Variation as a Tool to Trace the Authenticity of Beef. Anal. Bioanal. Chem. 2004, 378, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Franke, B.M.; Hadorn, R.; Bosset, J.O.; Gremaud, G.; Kreuzer, M. Is Authentication of the Geographic Origin of Poultry Meat and Dried Beef Improved by Combining Multiple Trace Element and Oxygen Isotope Analysis? Meat Sci. 2008, 80, 944–947. [Google Scholar] [CrossRef]
- Nakashita, R.; Suzuki, Y.; Akamatsu, F.; Iizumi, Y.; Korenaga, T.; Chikaraishi, Y. Stable Carbon, Nitrogen, and Oxygen Isotope Analysis as a Potential Tool for Verifying Geographical Origin of Beef. Anal. Chim. Acta 2008, 617, 148–152. [Google Scholar] [CrossRef]
- Bong, Y.-S.; Shin, W.-J.; Lee, A.-R.; Kim, Y.-S.; Kim, K.; Lee, K.-S. Tracing the Geographical Origin of Beefs Being Circulated in Korean Markets Based on Stable Isotopes. Rapid Commun. Mass Spectrom. 2010, 24, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Horacek, M.; Min, J.-S. Discrimination of Korean Beef from Beef of Other Origin by Stable Isotope Measurements. Food Chem. 2010, 121, 517–520. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, H.; Zhang, B.; Xu, Z.; Chen, A.; Zhao, Y. A Rapid Sample Preparation Method for the Analysis of Stable Isotope Ratios of Beef Samples from Different Countries. Rapid Commun. Mass Spectrom. 2020, 34, e8795. [Google Scholar] [CrossRef] [PubMed]
- Heaton, K.; Kelly, S.D.; Hoogewerff, J.; Woolfe, M. Verifying the Geographical Origin of Beef: The Application of Multi-Element Isotope and Trace Element Analysis. Food Chem. 2008, 107, 506–515. [Google Scholar] [CrossRef]
- Osorio, M.T.; Moloney, A.P.; Schmidt, O.; Monahan, F.J. Beef Authentication and Retrospective Dietary Verification Using Stable Isotope Ratio Analysis of Bovine Muscle and Tail Hair. J. Agric. Food Chem. 2011, 59, 3295–3305. [Google Scholar] [CrossRef]
- Galimov, E.M. Causes of Fractionation of Isotopes. The Biological Fractionation of Isotopes; Academic Press: Cambridge, MA, USA, 1985; pp. 1–15. [Google Scholar]
- Rubenstein, D.R.; Hobson, K.A. From Birds to Butterflies: Animal Movement Patterns and Stable Isotopes. Trends Ecol. Evol. 2004, 19, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Camin, F.; Wietzerbin, K.; Cortes, A.B.; Haberhauer, G.; Lees, M.; Versini, G. Application of Multielement Stable Isotope Ratio Analysis to the Characterization of French, Italian, and Spanish Cheeses. J. Agric. Food Chem. 2004, 52, 6592–6601. [Google Scholar] [CrossRef]
- Voerkelius, S.; Lorenz, G.D.; Rummel, S.; Quétel, C.R.; Heiss, G.; Baxter, M.; Brach-Papa, C.; Deters-Itzelsberger, P.; Hoelzl, S.; Hoogewerff, J.; et al. Strontium Isotopic Signatures of Natural Mineral Waters, the Reference to a Simple Geological Map and Its Potential for Authentication of Food. Food Chem. 2010, 118, 933–940. [Google Scholar] [CrossRef]
- Camin, F.; Larcher, R.; Nicolini, G.; Bontempo, L.; Bertoldi, D.; Perini, M.; Schlicht, C.; Schellenberg, A.; Thomas, F.; Heinrich, K.; et al. Isotopic and Elemental Data for Tracing the Origin of European Olive Oils. J. Agric. Food Chem. 2010, 58, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Schellenberg, A.; Chmielus, S.; Schlicht, C.; Camin, F.; Perini, M.; Bontempo, L.; Heinrich, K.; Kelly, S.D.; Rossmann, A.; Thomas, F.; et al. Multielement Stable Isotope Ratios (H, C, N, S) of Honey from Different European Regions. Food Chem. 2010, 121, 770–777. [Google Scholar] [CrossRef]
- Asfaha, D.G.; Quétel, C.R.; Thomas, F.; Horacek, M.; Wimmer, B.; Heiss, G.; Dekant, C.; Deters-Itzelsberger, P.; Hoelzl, S.; Rummel, S.; et al. Combining Isotopic Signatures of n(87Sr)/n(86Sr) and Light Stable Elements (C, N, O, S) with Multi-Elemental Profiling for the Authentication of Provenance of European Cereal Samples. J. Cereal Sci. 2011, 53, 170–177. [Google Scholar] [CrossRef]
- Rummel, S.; Dekant, C.H.; Hölzl, S.; Kelly, S.D.; Baxter, M.; Marigheto, N.; Quetel, C.R.; Larcher, R.; Nicolini, G.; Fröschl, H.; et al. Sr Isotope Measurements in Beef-Analytical Challenge and First Results. Anal. Bioanal. Chem. 2012, 402, 2837–2848. [Google Scholar] [CrossRef]
- Molkentin, J.; Giesemann, A. Follow-up of Stable Isotope Analysis of Organic versus Conventional Milk. Anal. Bioanal. Chem. 2010, 398, 1493–1500. [Google Scholar] [CrossRef]
- Bear, R.; Rintoul, D.; Snyder, B.; Smith-Caldas, M.; Herren, C.; Horne, E. Principles of Biology; Open Access Textbooks; New Prairie Press: Manhattan, KS, USA, 2016; Available online: https://newprairiepress.org/textbooks/1 (accessed on 7 February 2023).
- Pianezze, S.; Perini, M.; Bontempo, L.; Piasentier, E.; Poma, G.; Covaci, A.; Camin, F. Stable Isotope Ratio Analysis for the Characterisation of Edible Insects. J. Insects Food Feed 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Piasentier, E.; Valusso, R.; Camin, F.; Versini, G. Stable Isotope Ratio Analysis for Authentication of Lamb Meat. Meat Sci. 2003, 64, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Deniro, M.J.; Epstein, S. Carbon Isotopic Evidence for Different Feeding Patterns in Two Hyrax Species Occupying the Same Habitat. Science 1978, 201, 906–908. [Google Scholar] [CrossRef]
- Camin, F.; Perini, M.; Bontempo, L.; Galeotti, M.; Tibaldi, E.; Piasentier, E. Stable Isotope Ratios of H, C, O, N and S for the Geographical Traceability of Italian Rainbow Trout (Oncorhynchus Mykiss). Food Chem. 2018, 267, 288–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiljunen, M.; Grey, J.; Sinisalo, T.; Harrod, C.; Immonen, H.; Jones, R.I. A Revised Model for Lipid-Normalizing δ13C Values from Aquatic Organisms, with Implications for Isotope Mixing Models. J. Appl. Ecol. 2006, 43, 1213–1222. [Google Scholar] [CrossRef]
- Smet, S.D.; De Smet, S.; Balcaen, A.; Claeys, E.; Boeckx, P.; Van Cleemput, O. Stable Carbon Isotope Analysis of Different Tissues of Beef Animals in Relation to Their Diet. Rapid Commun. Mass Spectrom. 2004, 18, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Kornexl, B.E.; Werner, T.; Roßmann, A.; Schmidt, H.-L. Measurement of Stable Isotope Abundances in Milk and Milk Ingredients—A Possible Tool for Origin Assignment and Quality Control. Z. Für Lebensm.-Forsch. A 1997, 205, 19–24. [Google Scholar] [CrossRef]
- Otero, N.; Soler, A.; Canals, À. Fertilizer Characterization: Isotopic Data (N, S, O, C, and Sr). Environ. Sci. Technol. 2004, 38, 3254–3262. [Google Scholar] [CrossRef]
- Bateman, A.S.; Kelly, S.D.; Woolfe, M. Nitrogen Isotope Composition of Organically and Conventionally Grown Crops. J. Agric. Food Chem. 2007, 55, 2664–2670. [Google Scholar] [CrossRef] [PubMed]
- Gehl, R.J.; Schmidt, J.P.; Stone, L.R.; Schlegel, A.J.; Clark, G.A. In Situ Measurements of Nitrate Leaching Implicate Poor Nitrogen and Irrigation Management on Sandy Soils. J. Environ. Qual. 2005, 34, 2243–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- (Tanz), N.K.; Rossmann, A.; Schmidt, H.-L. Potentials and Caveats with Oxygen and Sulfur Stable Isotope Analyses in Authenticity and Origin Checks of Food and Food Commodities. Food Control 2015, 48, 143–150. [Google Scholar] [CrossRef]
- Krouse, H.R.; Roy Krouse, H.; Mayer, B. Sulphur and Oxygen Isotopes in Sulphate. In Environmental Tracers in Subsurface Hydrology; Springer: Boston, MA, USA, 2000; pp. 195–231. [Google Scholar]
- Mizota, C.; Sasaki, A. Sulfur Isotope Composition of Soils and Fertilizers: Differences between Northern and Southern Hemispheres. Geoderma 1996, 71, 77–93. [Google Scholar] [CrossRef]
- Cortecci, G.; Dinelli, E.; Boschetti, T.; Arbizzani, P.; Pompilio, P.; Mussi, M. The Serchio River catchment, northern Tuscany: Geochemistry of stream waters and sediments, and isotopic composition of dissolved sulfate. Appl. Geochem. 2008, 23, 1513–1543. [Google Scholar] [CrossRef]
- Zazzo, A.; Monahan, F.J.; Moloney, A.P.; Green, S.; Schmidt, O. Sulphur Isotopes in Animal Hair Track Distance to Sea. Rapid Commun. Mass Spectrom. 2011, 25, 2371–2378. [Google Scholar] [CrossRef] [PubMed]
- Krouse, H.R. Sulphur Isotopes in Our Environment. In Terrestrial Environment A; Elsevier Scientific: Amsterdam, The Netherlands, 1980; pp. 435–471. [Google Scholar]
- Cortecci, G. Isotopic constraints on the origin of sulfate in the Bolsena, Vico and Bracciano volcanic lakes, Latium. Italy. Miner. Petrogr. Acta 1996, 39, 15–24. [Google Scholar]
- Osorio, M.T.; Moloney, A.P.; Schmidt, O.; Monahan, F.J. Multielement Isotope Analysis of Bovine Muscle for Determination of International Geographical Origin of Meat. J. Agric. Food Chem. 2011, 59, 3285–3294. [Google Scholar] [CrossRef]
- Auerswald, K.; Rossmann, A.; Schäufele, R.; Schwertl, M.; Monahan, F.J.; Schnyder, H. Does Natural Weathering Change the Stable Isotope Composition (2H, 13C, 15N, 18O and 34S) of Cattle Hair? Rapid Commun. Mass Spectrom. 2011, 25, 3741–3748. [Google Scholar] [CrossRef] [PubMed]
- Hobson, K.A.; Atwell, L.; Wassenaar, L.I. Influence of Drinking Water and Diet on the Stable-Hydrogen Isotope Ratios of Animal Tissues. Proc. Natl. Acad. Sci. USA 1999, 96, 8003–8006. [Google Scholar] [CrossRef] [Green Version]
- Sharp, Z.D.; Atudorei, V.; Panarello, H.O.; Fernández, J.; Douthitt, C. Hydrogen Isotope Systematics of Hair: Archeological and Forensic Applications. J. Archaeol. Sci. 2003, 30, 1709–1716. [Google Scholar] [CrossRef]
- Dansgaard, W. Stable Isotopes in Precipitation. Tellus 1964, 16, 436–468. [Google Scholar] [CrossRef]
- Gat, J.R.; Gonfiantini, R. (Eds.) Technical Reports Series No 210; International Atomic Energy Agency: Vienna, Austria, 1981. [Google Scholar]
- Roden, J.S.; Ehleringer, J.R. There Is No Temperature Dependence of Net Biochemical Fractionation of Hydrogen and Oxygen Isotopes in Tree-Ring Cellulose. Isot. Environ. Health Stud. 2000, 36, 303–317. [Google Scholar] [CrossRef]
- Kohn, M.J. You Are What You Eat. Science 1999, 283, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Camin, F.; Bontempo, L.; Ziller, L.; Piangiolino, C.; Morchio, G. Stable Isotope Ratios of Carbon and Hydrogen to Distinguish Olive Oil from Shark Squalene-Squalane. Rapid Commun. Mass Spectrom. 2010, 24, 1810–1816. [Google Scholar] [CrossRef]
- Coplen, T.B. Guidelines and Recommended Terms for Expression of Stable-Isotope-Ratio and Gas-Ratio Measurement Results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef]
- Granato, D.; Putnik, P.; Kovačević, D.B.; Santos, J.S.; Calado, V.; Rocha, R.S.; Da Cruz, A.G.; Jarvis, B.; Rodionova, O.Y.; Pomerantsev, A. Trends in Chemometrics: Food Authentication, Microbiology, and Effects of Processing. Compr. Rev. Food Sci. Food Saf. 2018, 17, 663–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Group | Country | Site (N. Samples) | δ(2H) (‰, Vs. V–SMOW) | δ(13C) (‰, Vs. V–PDB) | δ(15N) (‰, Vs. Air) | δ(34S) (‰, Vs. V–CDT) |
|---|---|---|---|---|---|---|
| C_LL_C4 | Greece | Chalkidiki (4) | −99.0 ± 8.0 | −20.3 ± 2.1 | 4.3 ± 1.6 | 6.1 ± 0.6 |
| Spain | Barcelona (15) | −97.0 ± 3.6 | −18.4 ± 1.1 | 6.8 ± 0.5 | 7.1 ± 0.4 | |
| Italy | Florence (20) | −91.1 ± 2.7 | −18.9 ± 1.4 | 5.2 ± 0.6 | 4.4 ± 0.9 | |
| Sicily (20) | −87.0 ± 3.5 | −20.2 ± 1.3 | 4.4 ± 0.6 | 3.3 ± 1.6 | ||
| I_LL_MIX | Trento (33) | −101.5 ± 4.3 | −22.0 ± 1.8 | 4.8 ± 0.8 | 5.6 ± 0.5 | |
| Austria | Mühlviertel (4) | −110.3 ± 5.0 | −24.1 ± 2.6 | 4.7 ± 0.4 | 3.9 ± 0.1 | |
| France | Limousin (20) | −98.9 ± 6.0 | −21.3 ± 3.2 | 5.6 ± 0.6 | 6.6 ± 0.7 | |
| Germany | Frankonia (20) | −108.8 ± 4.7 | −22.9 ± 2.8 | 6.0 ± 1.0 | 4.1 ± 1.7 | |
| Allgäu (20) | −116.3 ± 4.1 | −25.0 ± 1.8 | 6.0 ± 1.2 | 3.3 ± 1.4 | ||
| Gäuboden (20) | −107.1 ± 3.7 | −22.3 ± 3.6 | 6.2 ± 0.9 | 5.3 ± 0.5 | ||
| C_ML_C3 | UK | Cornwall (20) | −90.0 ± 1.9 | −26.0 ± 1.1 | 7.7 ± 1.0 | 8.1 ± 1.6 |
| Orkney (23) | −96.9 ± 2.5 | −26.8 ± 0.4 | 7.9 ± 0.6 | 10.0 ± 3.1 | ||
| Ireland | Bohernagore (8) | −102.1 ± 2.6 | −26.7 ± 0.3 | 6.4 ± 1.0 | 8.8 ± 1.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bontempo, L.; Perini, M.; Pianezze, S.; Horacek, M.; Roßmann, A.; Kelly, S.D.; Thomas, F.; Heinrich, K.; Schlicht, C.; Schellenberg, A.; et al. Characterization of Beef Coming from Different European Countries through Stable Isotope (H, C, N, and S) Ratio Analysis. Molecules 2023, 28, 2856. https://doi.org/10.3390/molecules28062856
Bontempo L, Perini M, Pianezze S, Horacek M, Roßmann A, Kelly SD, Thomas F, Heinrich K, Schlicht C, Schellenberg A, et al. Characterization of Beef Coming from Different European Countries through Stable Isotope (H, C, N, and S) Ratio Analysis. Molecules. 2023; 28(6):2856. https://doi.org/10.3390/molecules28062856
Chicago/Turabian StyleBontempo, Luana, Matteo Perini, Silvia Pianezze, Micha Horacek, Andreas Roßmann, Simon D. Kelly, Freddy Thomas, Katharina Heinrich, Claus Schlicht, Antje Schellenberg, and et al. 2023. "Characterization of Beef Coming from Different European Countries through Stable Isotope (H, C, N, and S) Ratio Analysis" Molecules 28, no. 6: 2856. https://doi.org/10.3390/molecules28062856
APA StyleBontempo, L., Perini, M., Pianezze, S., Horacek, M., Roßmann, A., Kelly, S. D., Thomas, F., Heinrich, K., Schlicht, C., Schellenberg, A., Hoogewerff, J., Heiss, G., Wimmer, B., & Camin, F. (2023). Characterization of Beef Coming from Different European Countries through Stable Isotope (H, C, N, and S) Ratio Analysis. Molecules, 28(6), 2856. https://doi.org/10.3390/molecules28062856