
(Pro)Renin Receptor Decoy Peptide PRO20 Protects against Oxidative Renal Damage Induced by Advanced Oxidation Protein Products

Abstract
:1. Introduction
2. Results
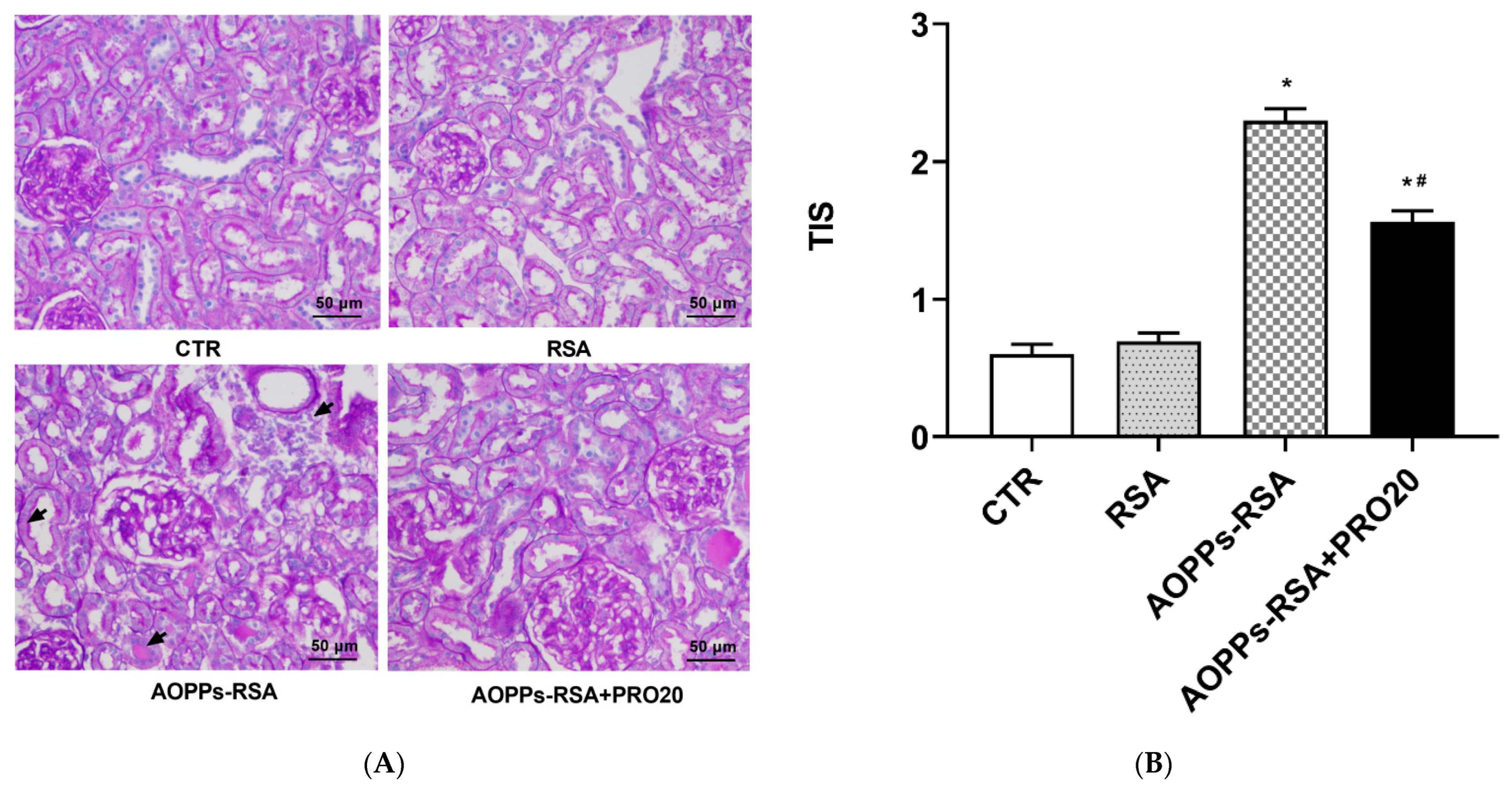
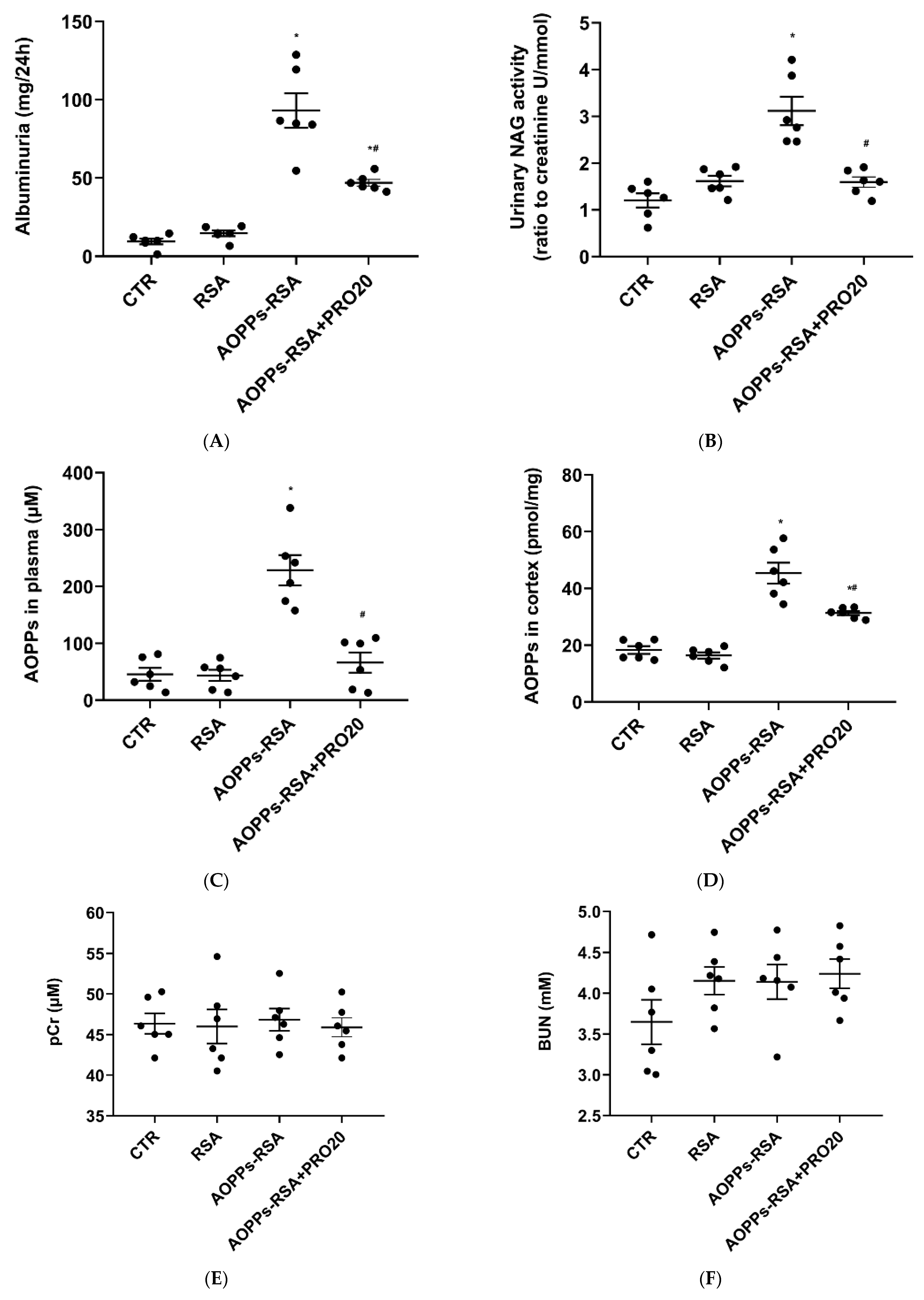
2.1. PRO20 Improved Kidney Histological Damage Caused by AOPPs-rat Serum Albumin (RSA)
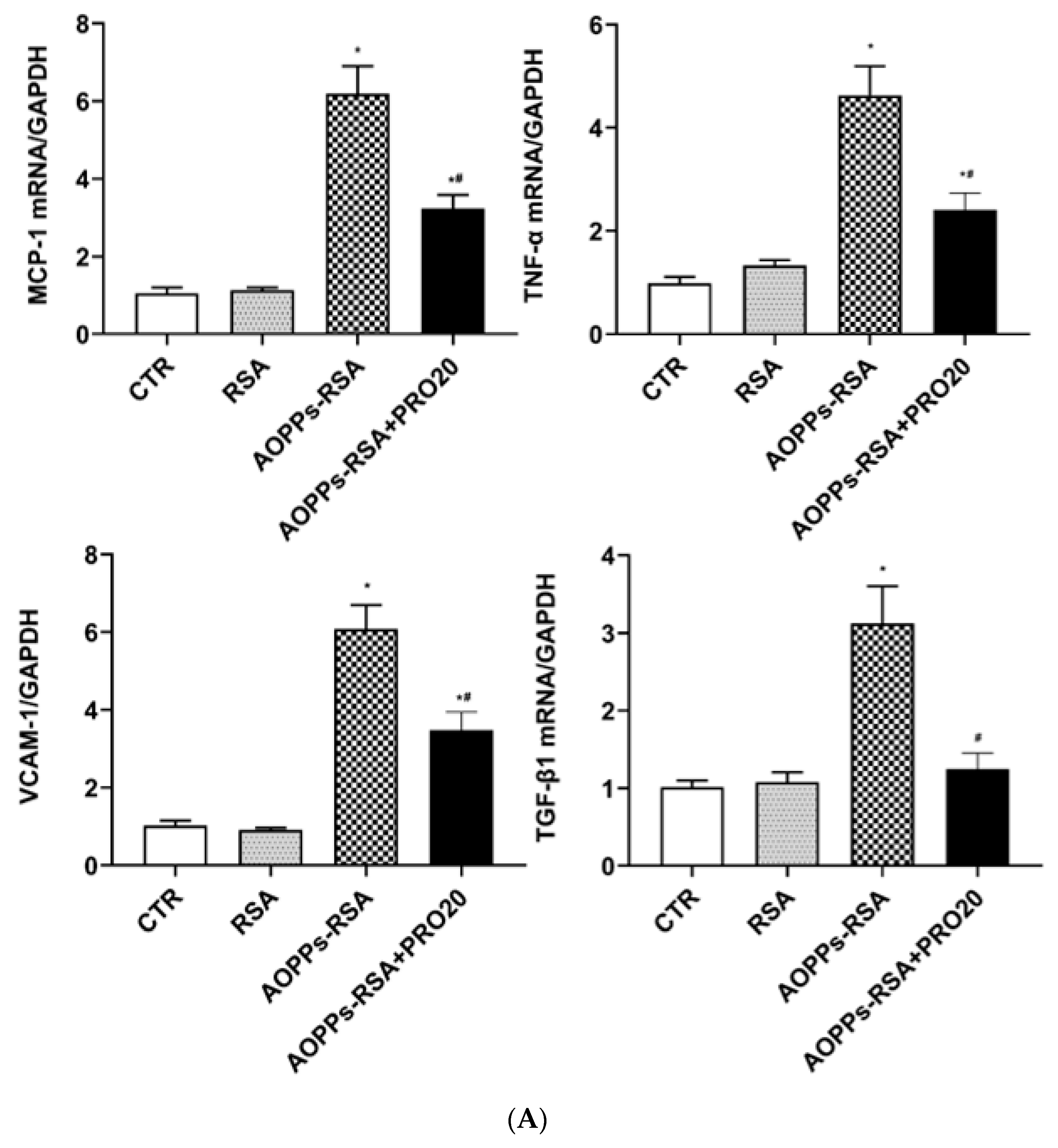
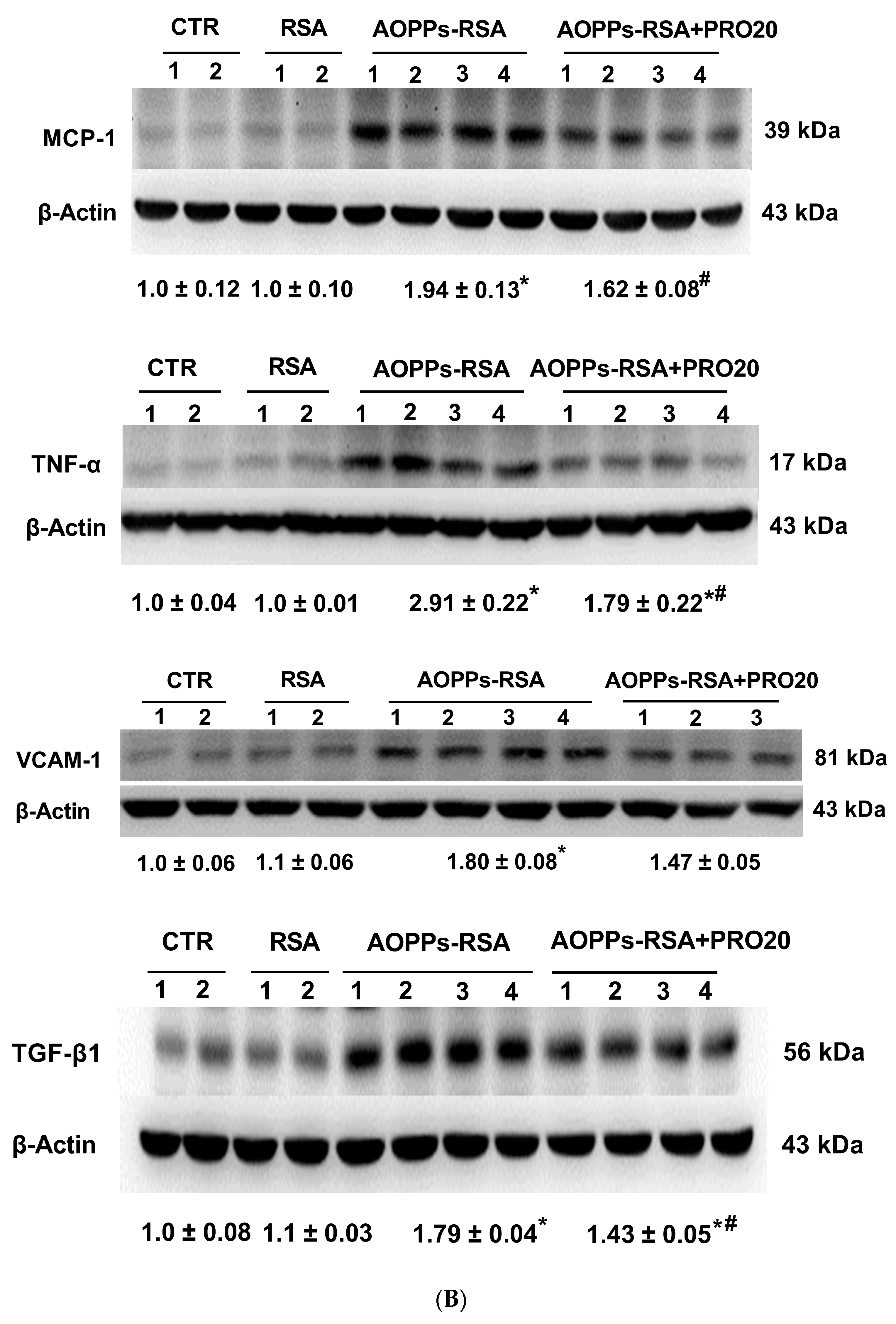
2.2. PRO20 Attenuated AOPPs-RSA-Induced Renal Inflammation and Fibrosis Markers
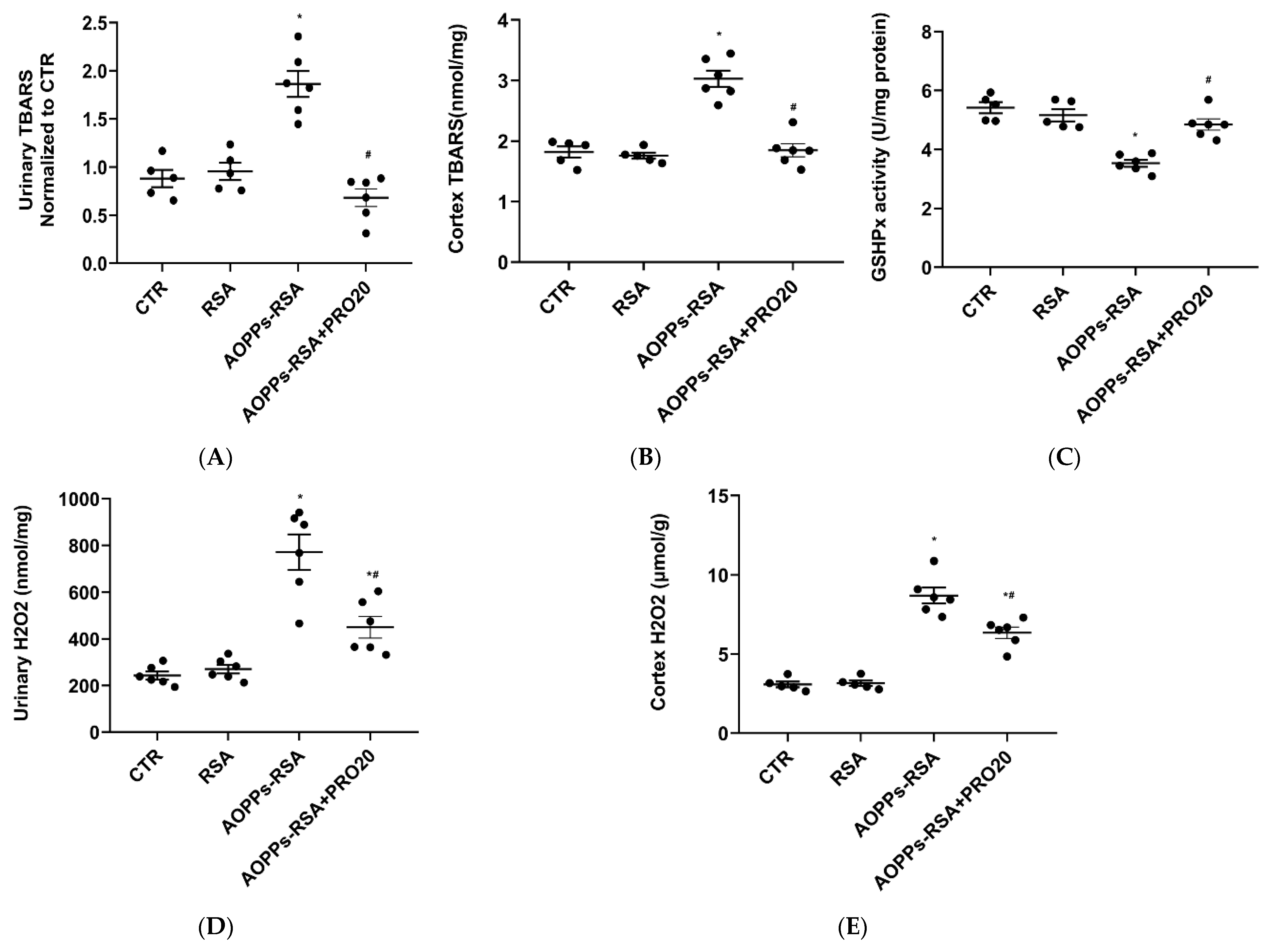
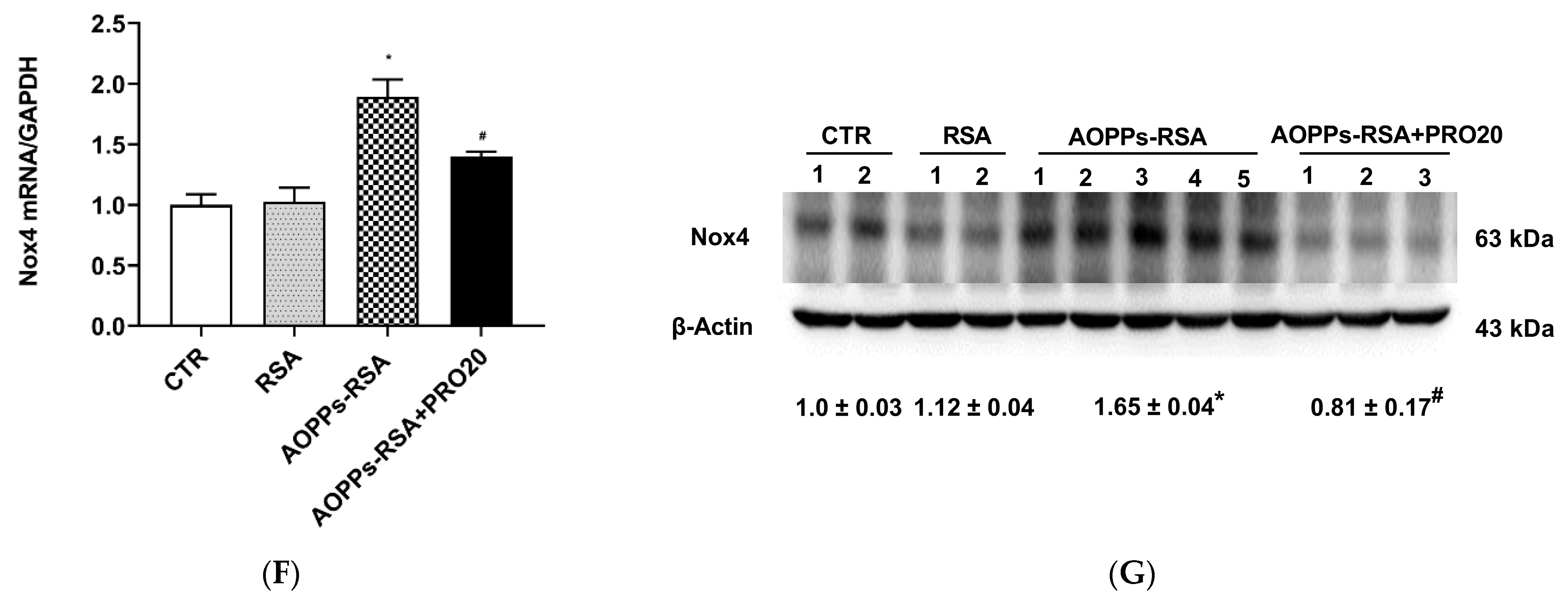
2.3. PRO20 Attenuated AOPPs-RSA-Induced Renal Oxidative Stresses
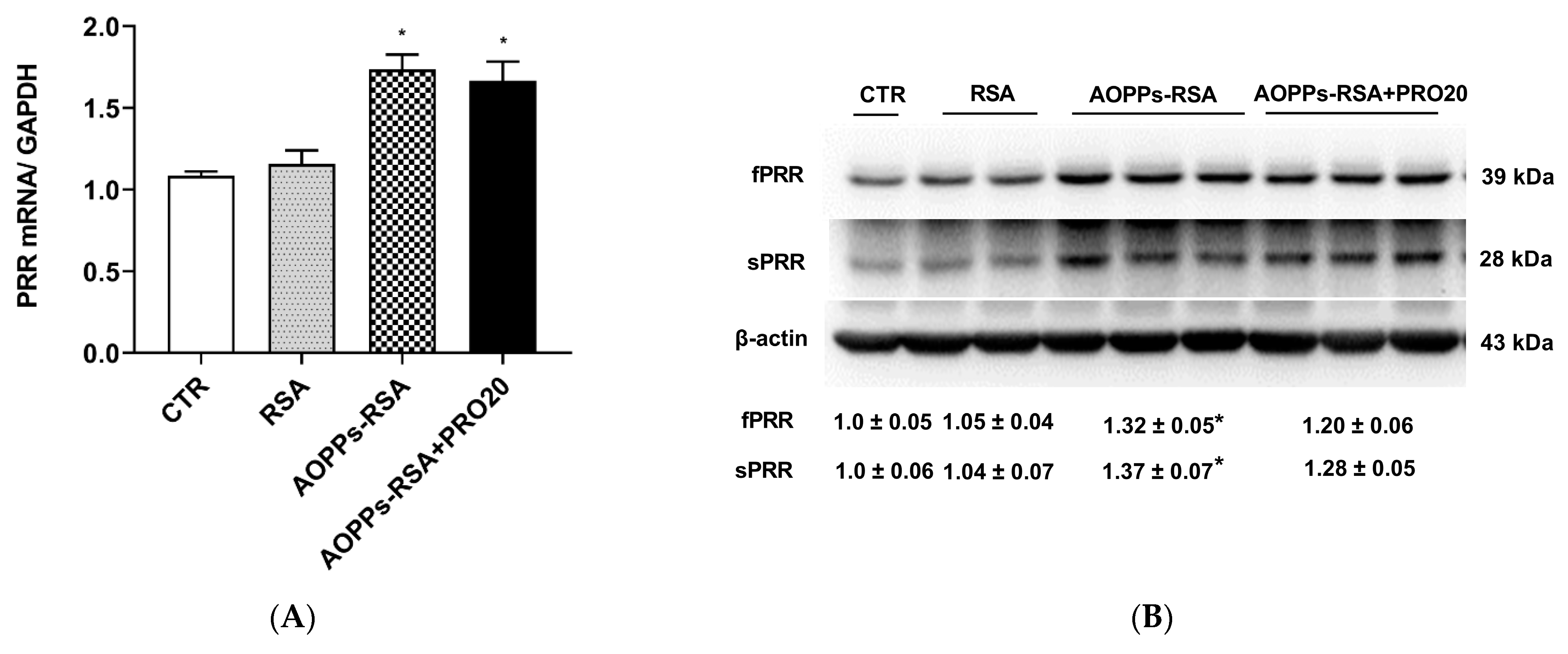
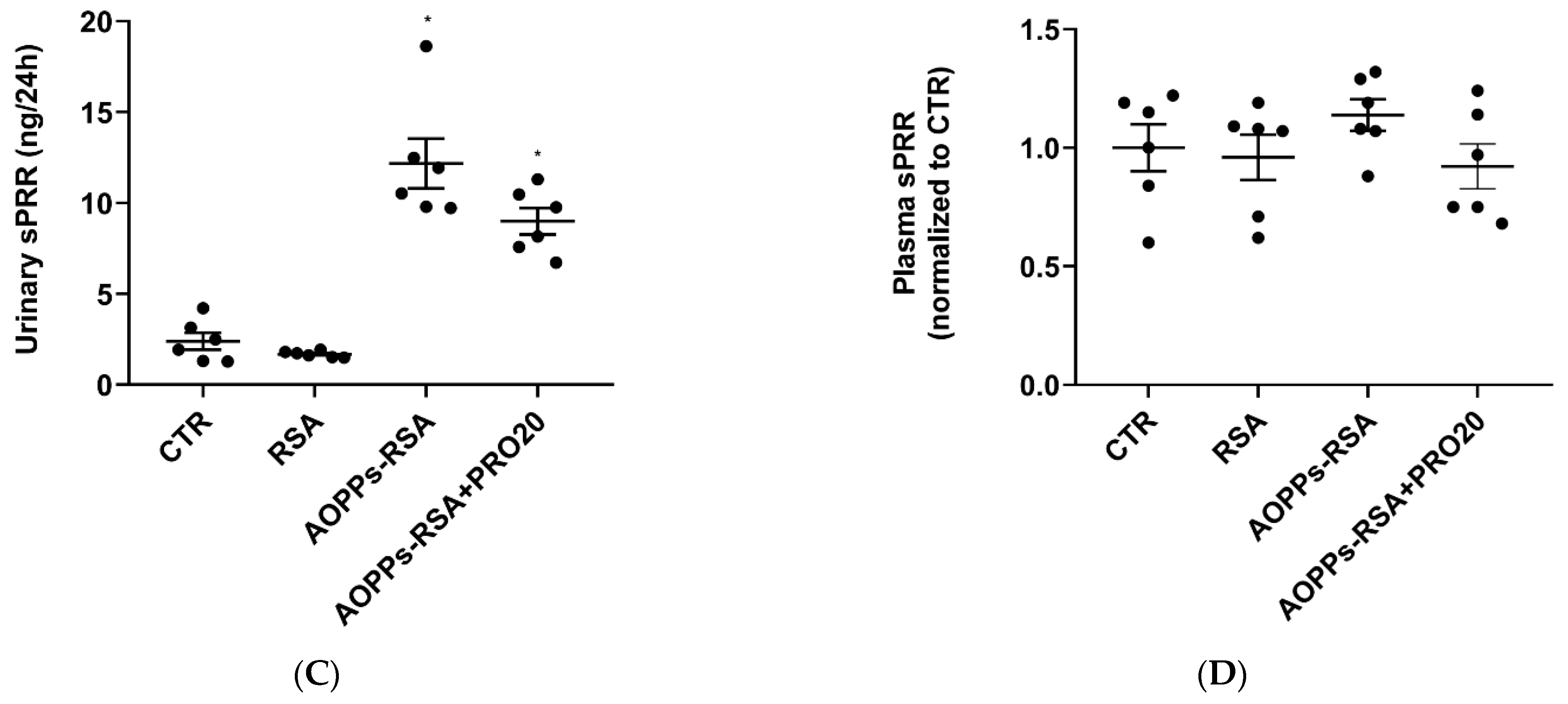
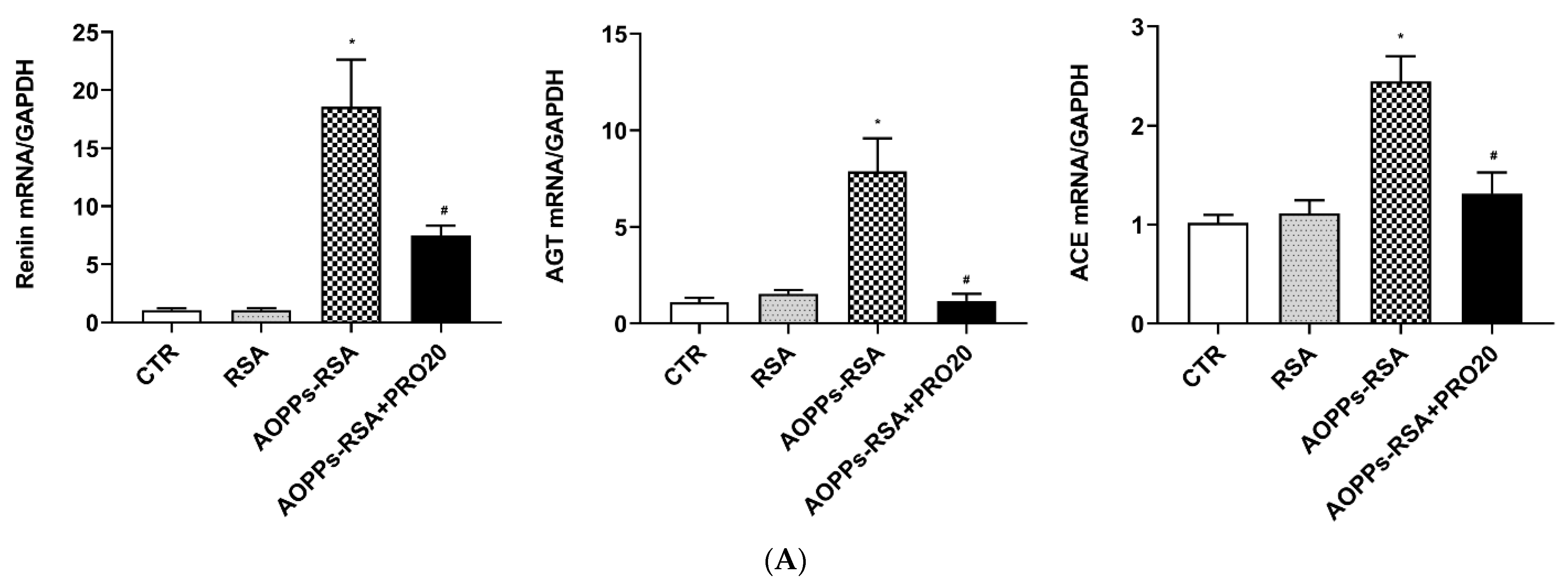
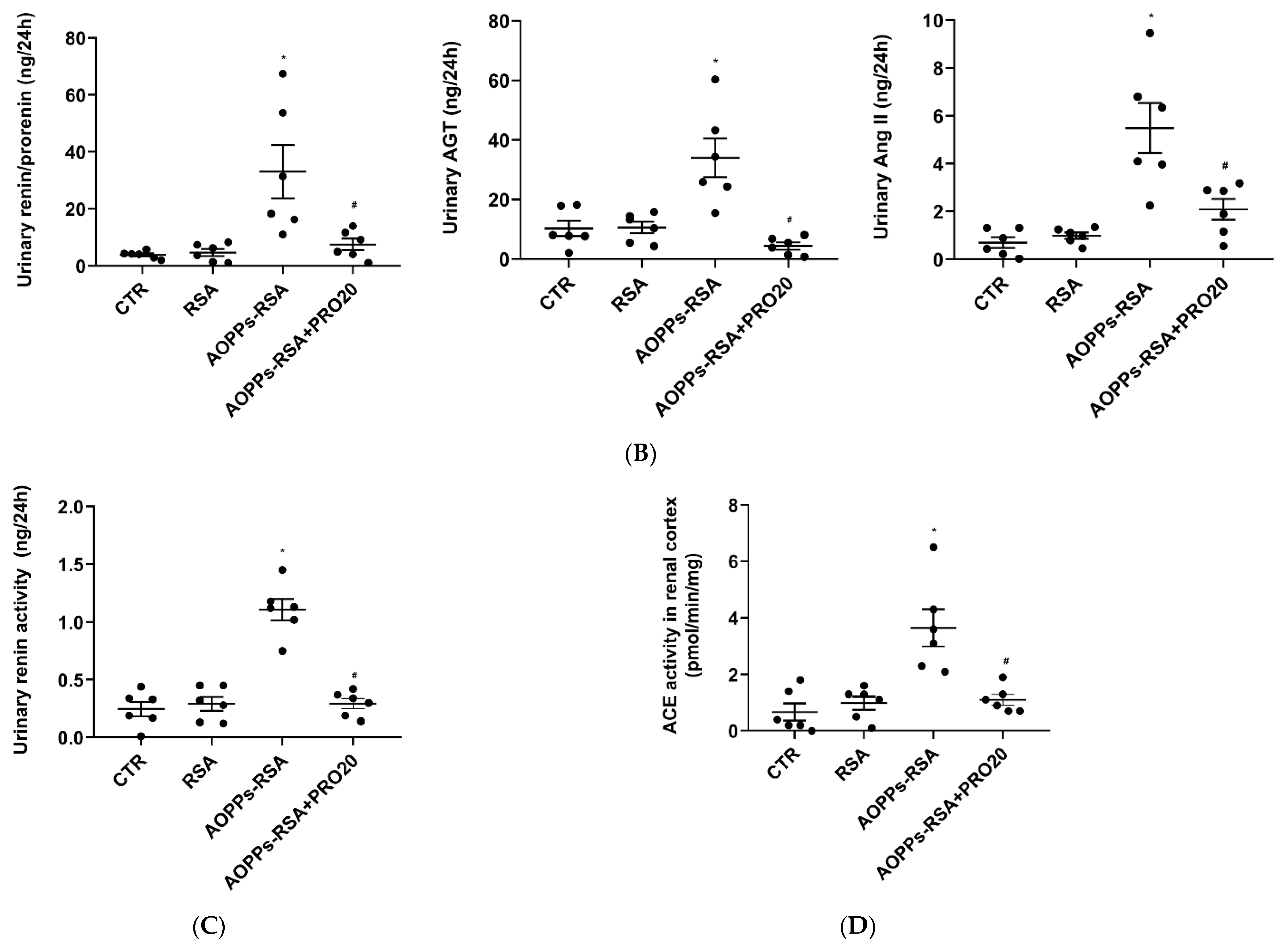
2.4. PRO20 Inhibited AOPPs-RSA-Induced Activation of Renal Local RAS
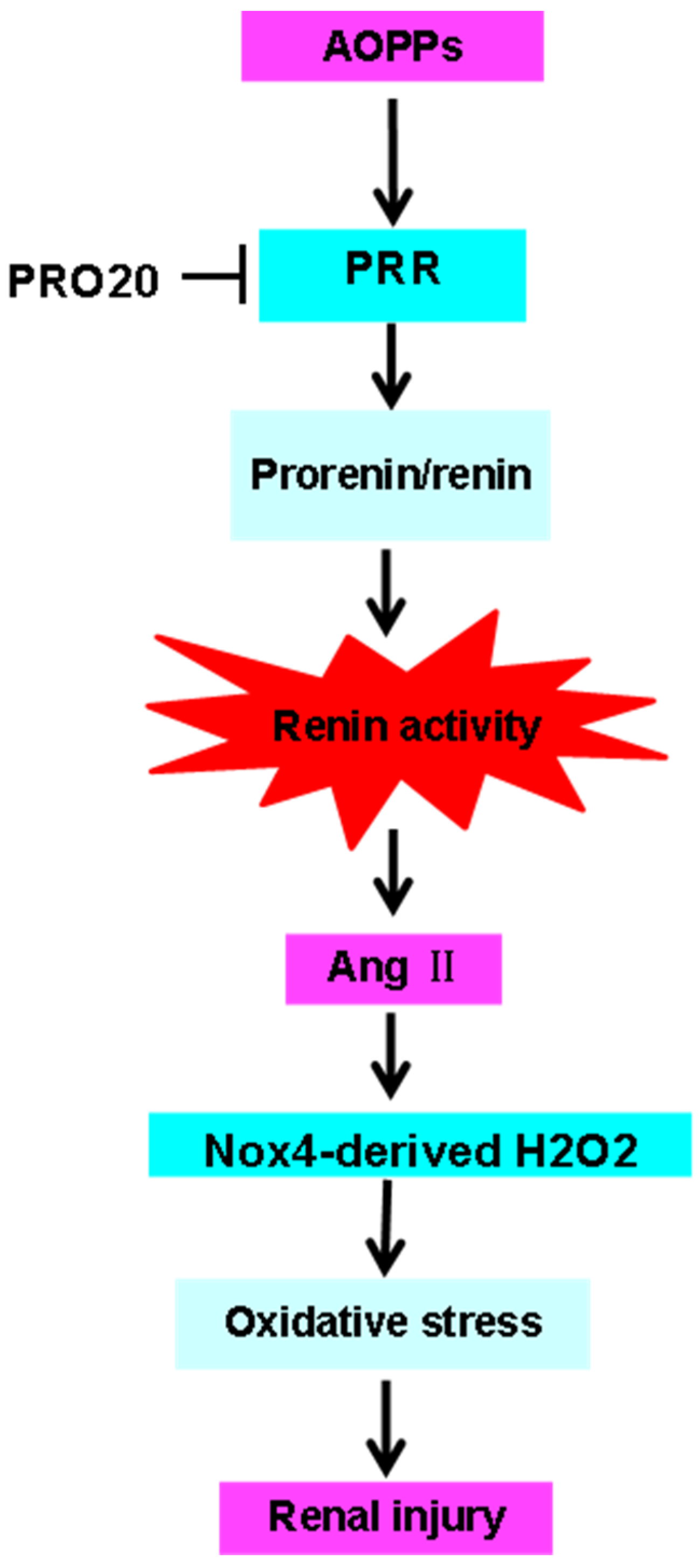
3. Discussion
4. Materials and Methods
4.1. Preparation and Measurement of AOPPs-RSA
4.2. Animals
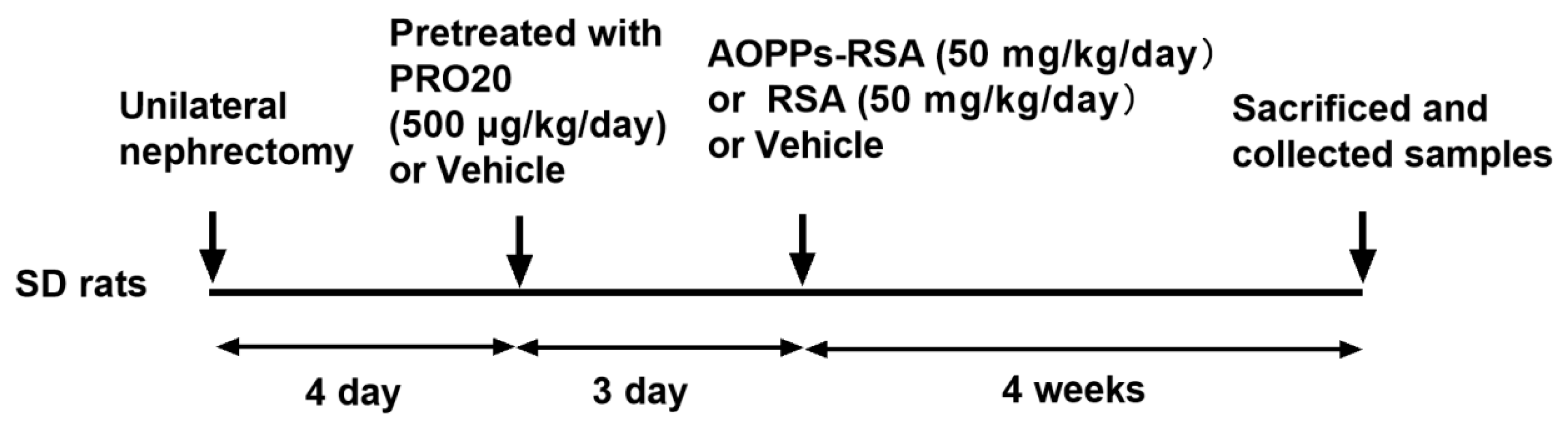
4.3. Experimental Design and Treatment Groups
4.4. Determination of Biochemical Parameters
4.5. Histopathological Assessment
4.6. Biochemical Analysis of AGT, ANG II, sPRR, ACE Activity, and Renin Activity
4.7. Measurement of TBARS Concentration, H2O2 and GSHPx Activity in Urine and Renal Cortex
4.8. Reverse Transcription Quantitative PCR (RT-qPCR)
4.9. Immunoblotting Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic kidney disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillere-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Hou, F.F.; Nie, J. AOPPs and the progression of kidney disease. Kidney Int. Suppl. 2014, 4, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Xu, J.; Zhou, Z.M.; Wang, G.B.; Hou, F.F.; Nie, J. Advanced oxidation protein products activate intrarenal renin-angiotensin system via a CD36-mediated, redox-dependent pathway. Antioxid. Redox Signal. 2013, 18, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.J.; Niu, H.X.; Hou, F.F.; Zhang, L.; Fu, N.; Nagai, R.; Lu, X.; Chen, B.H.; Shan, Y.X.; Tian, J.W.; et al. Advanced oxidation protein products activate vascular endothelial cells via a RAGE-mediated signaling pathway. Antioxid. Redox Signal. 2008, 10, 1699–1712. [Google Scholar] [CrossRef]
- Shi, X.Y.; Hou, F.F.; Niu, H.X.; Wang, G.B.; Xie, D.; Guo, Z.J.; Zhou, Z.M.; Yang, F.; Tian, J.W.; Zhang, X. Advanced oxidation protein products promote inflammation in diabetic kidney through activation of renal nicotinamide adenine dinucleotide phosphate oxidase. Endocrinology 2008, 149, 1829–1839. [Google Scholar] [CrossRef] [Green Version]
- Alagozlu, H.; Gorgul, A.; Bilgihan, A.; Tuncer, C.; Unal, S. Increased plasma levels of advanced oxidation protein products (AOPP) as a marker for oxidative stress in patients with active ulcerative colitis. Clin. Res. Hepatol. Gastroenterol. 2013, 37, 80–85. [Google Scholar] [CrossRef]
- Nguyen, G.; Delarue, F.; Burckle, C.; Bouzhir, L.; Giller, T.; Sraer, J.D. Pivotal role of the renin/prorenin receptor in angiotensin II production and cellular responses to renin. J. Clin. Investig. 2002, 109, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sullivan, M.N.; Zhang, S.; Worker, C.J.; Xiong, Z.; Speth, R.C.; Feng, Y. Intracerebroventricular infusion of the (Pro)renin receptor antagonist PRO20 attenuates deoxycorticosterone acetate-salt-induced hypertension. Hypertension 2015, 65, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lu, X.; Liu, M.; Feng, Y.; Zhou, S.F.; Yang, T. Renal medullary (pro)renin receptor contributes to angiotensin II-induced hypertension in rats via activation of the local renin-angiotensin system. BMC Med. 2015, 13, 278. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Deng, M.; Zhang, L.; Lu, A.; Su, J.; Xu, C.; Zhou, L.; Wang, L.; Ou, J.S.; Wang, W.; et al. Role of (pro)renin receptor in albumin overload-induced nephropathy in rats. Am. J. Physiol. Renal. Physiol. 2018, 315, F1759–F1768. [Google Scholar] [CrossRef]
- Luo, R.; Yang, K.; Wang, F.; Xu, C.; Yang, T. (Pro)renin receptor decoy peptide PRO20 protects against adriamycin-induced nephropathy by targeting the intrarenal renin-angiotensin system. Am. J. Physiol. Renal. Physiol. 2020, 319, F930–F940. [Google Scholar] [CrossRef]
- Nguyen, G. Renin and prorenin receptor in hypertension: What’s new? Curr. Hypertens. Rep. 2011, 13, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Xue, M.; Li, Y.; Jia, Y.J.; Zheng, Z.J.; Yang, Y.L.; Guan, M.P.; Sun, L.; Xue, Y.M. Early growth response 1 (Egr1) is a transcriptional activator of NOX4 in oxidative stress of diabetic kidney disease. J. Diabetes Res. 2018, 2018, 3405695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, X.; Wang, L.; Wang, Y.; Su, X.; Qi, Y.; Fan, Y.; Peng, Z. Intermedin inhibits unilateral ureteral obstruction-induced oxidative stress via NADPH oxidase Nox4 and cAMP-dependent mechanisms. Ren. Fail. 2017, 39, 652–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, K.; Wang, Y.; Wang, Y.; Fang, H. Advanced oxidation protein product promotes oxidative accentuation in renal epithelial cells via the soluble (Pro)renin receptor-mediated intrarenal renin-angiotensin system and Nox4-H2O2 signaling. Oxid. Med. Cell. Longev. 2021, 2021, 5710440. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, A.; Kaneshiro, Y.; Suzuki, F. Prorenin receptor blockers: Effects on cardiovascular complications of diabetes and hypertension. Expert Opin. Investig. Drugs 2006, 15, 1137–1139. [Google Scholar] [CrossRef]
- Saigo, S.; Kino, T.; Uchida, K.; Sugawara, T.; Chen, L.; Sugiyama, M.; Azushima, K.; Wakui, H.; Tamura, K.; Ishigami, T. Blood pressure elevation of tubular specific (P)RR transgenic mice and lethal tubular degeneration due to possible intracellular interactions between (P)RR and alternative renin products. Int. J. Mol. Sci. 2021, 23, 302. [Google Scholar] [CrossRef]
- Satofuka, S.; Ichihara, A.; Nagai, N.; Koto, T.; Shinoda, H.; Noda, K.; Ozawa, Y.; Inoue, M.; Tsubota, K.; Itoh, H.; et al. Role of nonproteolytically activated prorenin in pathologic, but not physiologic, retinal neovascularization. Investig. Ophthalmol. Vis. Sci. 2007, 48, 422–429. [Google Scholar] [CrossRef]
- Wang, F.; Lu, X.; Peng, K.; Du, Y.; Zhou, S.F.; Zhang, A.; Yang, T. Prostaglandin E-prostanoid4 receptor mediates angiotensin II-induced (pro)renin receptor expression in the rat renal medulla. Hypertension 2014, 64, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Lu, X.; Peng, K.; Fang, H.; Zhou, L.; Su, J.; Nau, A.; Yang, K.T.; Ichihara, A.; Lu, A.; et al. Antidiuretic action of collecting duct (Pro)renin receptor downstream of vasopressin and PGE2 receptor EP4. J. Am. Soc. Nephrol. 2016, 27, 3022–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousin, C.; Bracquart, D.; Contrepas, A.; Corvol, P.; Muller, L.; Nguyen, G. Soluble form of the (pro)renin receptor generated by intracellular cleavage by furin is secreted in plasma. Hypertension 2009, 53, 1077–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, T.; Geng, J.; Wu, Z.; Xu, L.; Liu, J.; Tian, J.; Zhou, Z.; Nie, J.; Bai, X. Advanced oxidation protein products promote lipotoxicity and tubulointerstitial fibrosis via CD36/beta-catenin pathway in diabetic nephropathy. Antioxid. Redox Signal. 2019, 31, 521–538. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Nguyen Khoa, T.; Capeillere-Blandin, C.; Nguyen, A.T.; Canteloup, S.; Dayer, J.M.; Jungers, P.; Drueke, T.; Descamps-Latscha, B. Advanced oxidation protein products as novel mediators of inflammation and monocyte activation in chronic renal failure. J. Immunol. 1998, 161, 2524–2532. [Google Scholar] [CrossRef]
- Sonawane, K.B.; Qian, J.; Hansen, R.A. Utilization patterns of antihypertensive drugs among the chronic kidney disease population in the United States: A cross-sectional analysis of the national health and nutrition examination survey. Clin. Ther. 2015, 37, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Xu, C. Physiology and pathophysiology of the intrarenal renin-angiotensin system: An update. J. Am. Soc. Nephrol. 2017, 28, 1040–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, G.; Muller, D.N. The biology of the (pro)renin receptor. J. Am. Soc. Nephrol. 2010, 21, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhou, L.; Wang, Y.; Miao, J.; Hong, X.; Hou, F.F.; Liu, Y. (Pro)renin receptor is an amplifier of Wnt/beta-catenin signaling in kidney injury and fibrosis. J. Am. Soc. Nephrol. 2017, 28, 2393–2408. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.B.; Mikhael, C.; Han, G.; de Faria, L.P.; Rody, W.J., Jr.; Holliday, L.S. Activation of (pro)renin by (pro)renin receptor in extracellular vesicles from osteoclasts. Sci. Rep. 2021, 11, 9214. [Google Scholar] [CrossRef]
- Nurun, N.A.; Uddin, N.M.; Nakagawa, T.; Iwata, H.; Ichihara, A.; Inagami, T.; Suzuki, F. Role of “handle” region of prorenin prosegment in the non-proteolytic activation of prorenin by binding to membrane anchored (pro)renin receptor. Front. Biosci. 2007, 12, 4810–4817. [Google Scholar] [CrossRef] [Green Version]
- Giese, M.J.; Speth, R.C. The ocular renin-angiotensin system: A therapeutic target for the treatment of ocular disease. Pharmacol. Ther. 2014, 142, 11–32. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Ichihara, A.; Mizuguchi, Y.; Sakoda, M.; Kurauchi-Mito, A.; Narita, T.; Kinouchi, K.; Bokuda, K.; Itoh, H. Add-on blockade of (pro)renin receptor in imidapril-treated diabetic SHRsp. Front. Biosci. (Elite Ed.) 2010, 2, 972–979. [Google Scholar] [CrossRef] [Green Version]
- Feldt, S.; Maschke, U.; Dechend, R.; Luft, F.C.; Muller, D.N. The putative (pro)renin receptor blocker HRP fails to prevent (pro)renin signaling. J. Am. Soc. Nephrol. 2008, 19, 743–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Hu, J.; Zhou, L.; Chen, Y.; Deng, M.; Liu, X.; Su, J.; Lu, A.; Fu, X.; Yang, T. (Pro)renin receptor contributes to pregnancy-induced sodium-water retention in rats via activation of intrarenal RAAS and alpha-ENaC. Am. J. Physiol. Renal Physiol. 2019, 316, F530–F538. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Fang, H.; Zhou, L.; Lu, A.; Yang, T. High potassium promotes mutual interaction between (pro)renin receptor and the local renin-angiotensin-aldosterone system in rat inner medullary collecting duct cells. Am. J. Physiol. Cell Physiol. 2016, 311, C686–C695. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Lu, A.; Lu, X.; Zhang, L.; Fang, H.; Zhou, L.; Yang, T. Activation of renal (Pro)renin receptor contributes to high fructose-induced salt sensitivity. Hypertension 2017, 69, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Lu, A.; Wang, H.; Fang, H.; Zhou, L.; Sun, P.; Yang, T. (Pro)Renin receptor regulates potassium homeostasis through a local mechanism. Am. J. Physiol. Renal Physiol. 2017, 313, F641–F656. [Google Scholar] [CrossRef]
- Kaneshiro, Y.; Ichihara, A.; Sakoda, M.; Takemitsu, T.; Nabi, A.H.; Uddin, M.N.; Nakagawa, T.; Nishiyama, A.; Suzuki, F.; Inagami, T.; et al. Slowly progressive, angiotensin II-independent glomerulosclerosis in human (pro)renin receptor-transgenic rats. J. Am. Soc. Nephrol. 2007, 18, 1789–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, G.; Caccamo, D.; Siligato, R.; Gembillo, G.; Satta, E.; Pazzano, D.; Carucci, N.; Carella, A.; Campo, G.D.; Salvo, A.; et al. Association of higher advanced oxidation protein products (AOPPs) levels in patients with diabetic and hypertensive nephropathy. Medicina 2019, 55, 675. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, J.; Namba, T.; Takabatake, Y.; Kimura, T.; Takahashi, A.; Yamamoto, T.; Minami, S.; Sakai, S.; Fujimura, R.; Kaimori, J.Y.; et al. Antioxidant role of autophagy in maintaining the integrity of glomerular capillaries. Autophagy 2018, 14, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Vendrov, A.E.; Stevenson, M.D.; Lozhkin, A.; Hayami, T.; Holland, N.A.; Yang, X.; Moss, N.; Pan, H.; Wickline, S.A.; Stockand, J.D.; et al. Renal NOXA1/NOX1 signaling regulates epithelial sodium channel and sodium retention in angiotensin II-induced hypertension. Antioxid. Redox Signal. 2022, 36, 550–566. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, Z.; Zhang, Z.; Obata, F.; Yang, X.; Zhang, X.; Huang, Y.; Mitsui, T.; Fan, J.; Takeda, M.; et al. Connexin43 contributes to inflammasome activation and lipopolysaccharide-initiated acute renal injury via modulation of intracellular oxidative status. Antioxid. Redox Signal. 2019, 31, 1194–1212. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, F.R.; Wang, J.N.; Gao, L.; Jiang, L.; Li, H.D.; Ma, Q.; Liu, X.Q.; Wei, B.; Zhou, L.; et al. Nox4 in renal diseases: An update. Free Radic. Biol. Med. 2018, 124, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Geiszt, M.; Kopp, J.B.; Varnai, P.; Leto, T.L. Identification of renox, an NAD(P)H oxidase in kidney. Proc. Natl. Acad. Sci. USA 2000, 97, 8010–8014. [Google Scholar] [CrossRef] [Green Version]
- Schader, T.; Lowe, O.; Reschke, C.; Malacarne, P.; Hahner, F.; Muller, N.; Gajos-Draus, A.; Backs, J.; Schroder, K. Oxidation of HDAC4 by Nox4-derived H2O2 maintains tube formation by endothelial cells. Redox Biol. 2020, 36, 101669. [Google Scholar] [CrossRef]
- Lu, X.; Wang, F.; Liu, M.; Yang, K.T.; Nau, A.; Kohan, D.E.; Reese, V.; Richardson, R.S.; Yang, T. Activation of ENaC in collecting duct cells by prorenin and its receptor PRR: Involvement of Nox4-derived hydrogen peroxide. Am. J. Physiol. Renal Physiol. 2016, 310, F1243–F1250. [Google Scholar] [CrossRef] [Green Version]
- Schnelle, M.; Sawyer, I.; Anilkumar, N.; Mohamed, B.A.; Richards, D.A.; Toischer, K.; Zhang, M.; Catibog, N.; Sawyer, G.; Mongue-Din, H.; et al. NADPH oxidase-4 promotes eccentric cardiac hypertrophy in response to volume overload. Cardiovasc. Res. 2021, 117, 178–187. [Google Scholar] [CrossRef]
- Obert, L.A.; Frazier, K.S. Intrarenal renin-angiotensin system involvement in the pathogenesis of chronic progressive nephropathy-bridging the informational gap between disciplines. Toxicol. Pathol. 2019, 47, 799–816. [Google Scholar] [CrossRef]
- Graciano, M.L.; de Cassia Cavaglieri, R.; Delle, H.; Dominguez, W.V.; Casarini, D.E.; Malheiros, D.M.; Noronha, I.L. Intrarenal Renin-Angiotensin system is upregulated in experimental model of progressive renal disease induced by chronic inhibition of nitric oxide synthesis. J. Am. Soc. Nephrol. 2004, 15, 1805–1815. [Google Scholar] [CrossRef] [Green Version]
- Sachse, A.; Wolf, G. Angiotensin II-induced reactive oxygen species and the kidney. J. Am. Soc. Nephrol. 2007, 18, 2439–2446. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Li, W.; Seth, D.M.; Nair, A.R.; Francis, J.; Feng, Y. (Pro)renin receptor mediates both angiotensin II-dependent and -independent oxidative stress in neuronal cells. PLoS ONE 2013, 8, e58339. [Google Scholar] [CrossRef]
- Fang, H.; Xu, C.; Lu, A.; Zou, C.J.; Xie, S.; Chen, Y.; Zhou, L.; Liu, M.; Wang, L.; Wang, W.; et al. (Pro)renin receptor mediates albumin-induced cellular responses: Role of site-1 protease-derived soluble (pro)renin receptor in renal epithelial cells. Am. J. Physiol. Cell Physiol. 2017, 313, C632–C643. [Google Scholar] [CrossRef]
- Nakagawa, T.; Suzuki-Nakagawa, C.; Watanabe, A.; Asami, E.; Matsumoto, M.; Nakano, M.; Ebihara, A.; Uddin, M.N.; Suzuki, F. Site-1 protease is required for the generation of soluble (pro)renin receptor. J. Biochem. 2017, 161, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, A.; Aizaki, Y.; Kusano, K.; Kishi, F.; Susumu, T.; Iida, S.; Ishiura, S.; Nishimura, S.; Shichiri, M.; Senbonmatsu, T. The (pro)renin receptor is cleaved by ADAM19 in the Golgi leading to its secretion into extracellular space. Hypertens. Res. 2011, 34, 599–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruciat, C.M.; Ohkawara, B.; Acebron, S.P.; Karaulanov, E.; Reinhard, C.; Ingelfinger, D.; Boutros, M.; Niehrs, C. Requirement of prorenin receptor and vacuolar H+-ATPase-mediated acidification for Wnt signaling. Science 2010, 327, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Xu, C.; Yu, J. The soluble (Pro)renin receptor in health and diseases: Foe or friend? J. Pharmacol. Exp. Ther. 2021, 378, 251–261. [Google Scholar] [CrossRef]
- Gonzalez, A.A.; Lara, L.S.; Luffman, C.; Seth, D.M.; Prieto, M.C. Soluble form of the (pro)renin receptor is augmented in the collecting duct and urine of chronic angiotensin II-dependent hypertensive rats. Hypertension 2011, 57, 859–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Wang, F.; Xu, C.; Soodvilai, S.; Peng, K.; Su, J.; Zhao, L.; Yang, K.T.; Feng, Y.; Zhou, S.F.; et al. Soluble (pro)renin receptor via beta-catenin enhances urine concentration capability as a target of liver X receptor. Proc. Natl. Acad. Sci. USA 2016, 113, E1898–E1906. [Google Scholar] [CrossRef] [Green Version]
- Gatineau, E.; Gong, M.C.; Yiannikouris, F. Soluble prorenin receptor increases blood pressure in high fat-fed male mice. Hypertension 2019, 74, 1014–1020. [Google Scholar] [CrossRef]
- Xie, S.; Su, J.; Lu, A.; Lai, Y.; Mo, S.; Pu, M.; Yang, T. Soluble (pro)renin receptor promotes the fibrotic response in renal proximal tubule epithelial cells in vitro via the Akt/beta-catenin/Snail signaling pathway. Am. J. Physiol. Renal Physiol. 2020, 319, F941–F953. [Google Scholar] [CrossRef]
- Li, H.Y.; Hou, F.F.; Zhang, X.; Chen, P.Y.; Liu, S.X.; Feng, J.X.; Liu, Z.Q.; Shan, Y.X.; Wang, G.B.; Zhou, Z.M.; et al. Advanced oxidation protein products accelerate renal fibrosis in a remnant kidney model. J. Am. Soc. Nephrol. 2007, 18, 528–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadola, L.; Noboa, O.; Marquez, M.N.; Rodriguez, M.J.; Nin, N.; Boggia, J.; Ferreiro, A.; Garcia, S.; Ortega, V.; Musto, M.L.; et al. Calcium citrate ameliorates the progression of chronic renal injury. Kidney Int. 2004, 65, 1224–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maral, J.; Puget, K.; Michelson, A.M. Comparative study of superoxide dismutase, catalase and glutathione peroxidase levels in erythrocytes of different animals. Biochem. Biophys. Res. Commun. 1977, 77, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Xue, K.; Gao, F.; Li, C.; Fang, H. Elevated reactivity of Apelin inhibited renal fibrosis induced by chronic intermittent hypoxia. Arch. Biochem. Biophys. 2021, 711, 109021. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Primer Sequence (5′–3′) | Reverse Primer Sequence (5′–3′) |
|---|---|---|
| MCP-1 | TAGCATCCACGTGCTGTCTC | CAGCCGACTCATTGGGATCA |
| TNF-α | CGTCAGCCGATTTGCCATTT | TCCCTCAGGGGTGTCCTTAG |
| VCAM-1 | GGCTCGTACACCATCCGC | CGGTTTTCGATTCACACTCGT |
| TGF-β1 | CTCAACACCTGCACAGCTCC | AGTTGGCATGGTAGCCCTTG |
| PRR | ATCCTTGAGACGAAACAAGA | AGCCAGTCATAATCCACAGT |
| REN | GATCACCATGAAGGGGGTCTCTGT | GTTCCTGAAGGGATTCTTTTGCAC |
| AGT | AGCATCCTCCTTGAACTCCA | TGATTTTTGCCCAGGATAGC |
| ACE | GAGCCATCCTTCCCTTTTTC | CCACATGTTCCCTAGCAGGT |
| Nox4 | TGTGCCGAACACTCTTGGC | ATATGCACGCCTGAGAAAATA |
| GAPDH | AGACAGCCGCATCTTCTTGT | TTCCCATTCTCAGCCTTGAC |
| Items | Details |
|---|---|
| Reaction system | FastStart Universal SYBR Green Master (ROX): 5.0 μL; forward and reverse primer (1.5 µM): 3.0 μL; cDNA: 2.0 μL. |
| Reaction conditions | 95 °C for 10 min; 40 cycles of 95 °C for 10 s and 60 °C for 30 s; followed by a melting curve. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, H.; Yang, T.; Zhou, B.; Li, X. (Pro)Renin Receptor Decoy Peptide PRO20 Protects against Oxidative Renal Damage Induced by Advanced Oxidation Protein Products. Molecules 2023, 28, 3017. https://doi.org/10.3390/molecules28073017
Fang H, Yang T, Zhou B, Li X. (Pro)Renin Receptor Decoy Peptide PRO20 Protects against Oxidative Renal Damage Induced by Advanced Oxidation Protein Products. Molecules. 2023; 28(7):3017. https://doi.org/10.3390/molecules28073017
Chicago/Turabian StyleFang, Hui, Teng Yang, Baolong Zhou, and Xinxuan Li. 2023. "(Pro)Renin Receptor Decoy Peptide PRO20 Protects against Oxidative Renal Damage Induced by Advanced Oxidation Protein Products" Molecules 28, no. 7: 3017. https://doi.org/10.3390/molecules28073017
APA StyleFang, H., Yang, T., Zhou, B., & Li, X. (2023). (Pro)Renin Receptor Decoy Peptide PRO20 Protects against Oxidative Renal Damage Induced by Advanced Oxidation Protein Products. Molecules, 28(7), 3017. https://doi.org/10.3390/molecules28073017