
In Vitro and In Silico Studies on Angiotensin I-Converting Enzyme Inhibitory Peptides Found in Hydrophobic Domains of Porcine Elastin


Abstract
:1. Introduction
2. Results
2.1. Purity of Synthetic Peptides
2.2. ACE Inhibitory Activity of Peptides Found in Tropoelastin
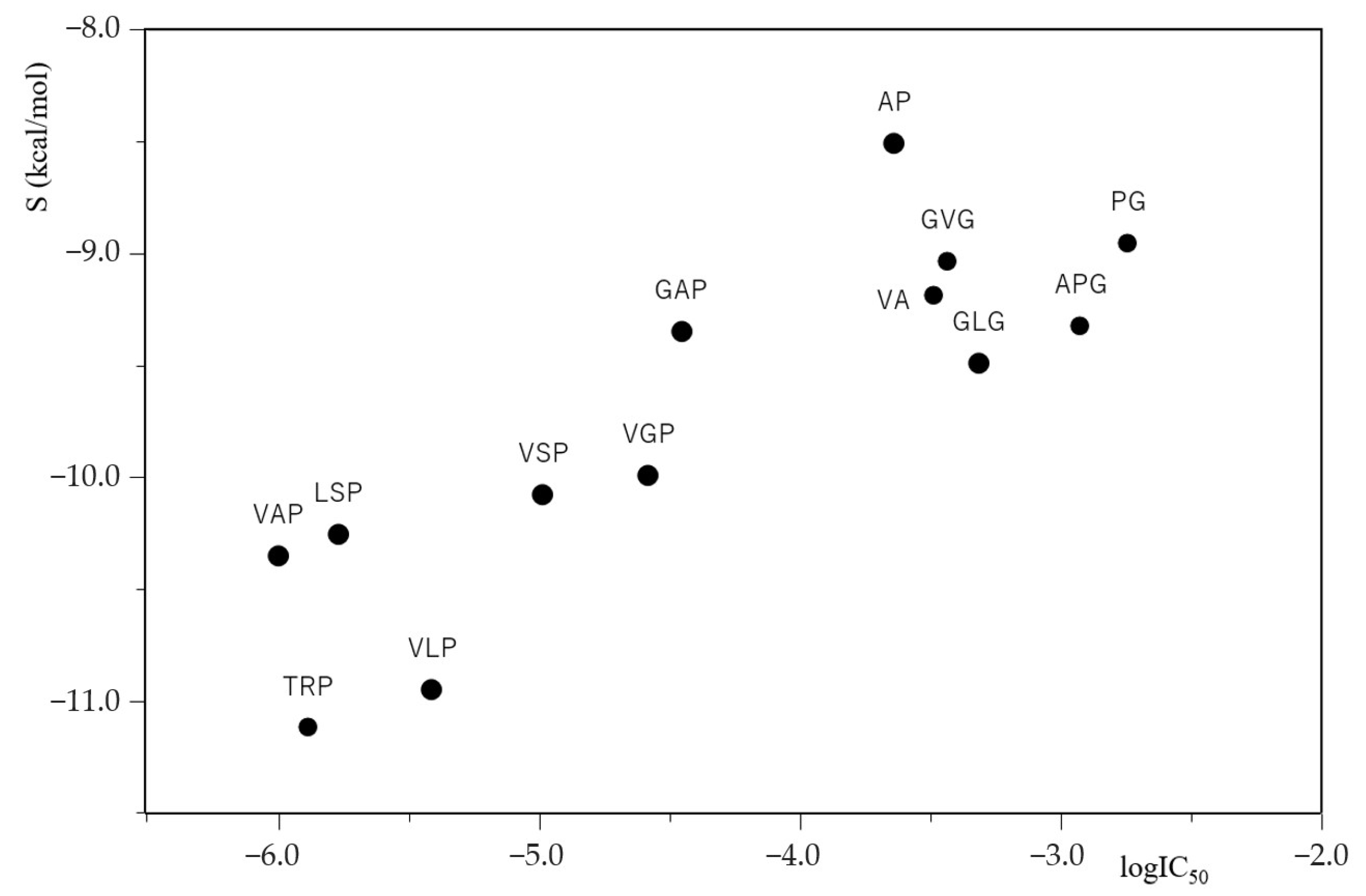
2.3. Relationship between the IC50 and Docking Score S of ACE Inhibitory Peptides
2.4. Informative Data by Online Tools on ACE Inhibitory Peptides
2.5. Molecular Docking Simulation on ACE Inhibitory Peptides
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Assay of ACE Inhibitory Activity
4.3. Molecular Docking
4.4. Online Analysis for ACE Inhibitory Peptides
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Egan, B.M.; Kjeldsen, S.E.; Grassi, G.; Esler, M.; Mancia, G. The Global Burden of Hypertension Exceeds 1.4 Billion People. J. Hypertens. 2019, 37, 1148–1153. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Toldrá, F. ACEI-Inhibitory Peptides Naturally Generated in Meat and Meat Products and Their Health Relevance. Nutrients 2018, 10, 1259. [Google Scholar] [CrossRef] [Green Version]
- Pihlanto-Leppälä, A. Bioactive Peptides Derived from Bovine Whey Proteins. Trends Food Sci. Technol. 2000, 11, 347–356. [Google Scholar] [CrossRef]
- Wood, R. Bronchospasm and Cough as Adverse Reactions to the ACE Inhibitors Captopril, Enalapril and Lisinopril. A Controlled Retrospective Cohort Study. Br. J. Clin. Pharmacol. 1995, 39, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and Characterization of Angiotensin I-Converting Enzyme Inhibitors from Sour Milk. J. Dairy Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Tu, M.; Wang, C.; Chen, C.; Zhang, R.; Liu, H.; Lu, W.; Jiang, L.; Du, M. Identification of a Novel ACE-Inhibitory Peptide from Casein and Evaluation of the Inhibitory Mechanisms. Food Chem. 2018, 256, 98–104. [Google Scholar] [CrossRef]
- Vukic, V.R.; Vukic, D.V.; Milanovic, S.D.; Ilicic, M.D.; Kanuric, K.G.; Johnson, M.S. In Silico Identification of Milk Antihypertensive Di- and Tripeptides Involved in Angiotensin I–Converting Enzyme Inhibitory Activity. Nutr. Res. 2017, 46, 22–30. [Google Scholar] [CrossRef]
- Baba, W.N.; Baby, B.; Mudgil, P.; Gan, C.-Y.; Vijayan, R.; Maqsood, S. Pepsin Generated Camel Whey Protein Hydrolysates with Potential Antihypertensive Properties: Identification and Molecular Docking of Antihypertensive Peptides. LWT 2021, 143, 111135. [Google Scholar] [CrossRef]
- Yu, Z.; Wu, S.; Zhao, W.; Ding, L.; Shiuan, D.; Chen, F.; Li, J.; Liu, J. Identification and the Molecular Mechanism of a Novel Myosin-Derived ACE Inhibitory Peptide. Food Funct. 2018, 9, 364–370. [Google Scholar] [CrossRef]
- Ohta, T.; Iwashita, A.; Sasaki, S.; Kawamura, Y. Antihypertensive Action of the Orally Administered Protease Hydrolysates of Chum Salmon Head and Their Angiotensin I-Converting Enzyme Inhibitory Peptides. Food Sci. Technol. Int. Tokyo 1997, 3, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Byun, H.-G.; Kim, S.-K. Structure and Activity of Angiotensin I Converting Enzyme Inhibitory Peptides Derived from Alaskan Pollack Skin. BMB Rep. 2002, 35, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Qin, S.; Li, W. Purification and Characterization of a Novel Angiotensin I-converting Enzyme-inhibitory Peptide Derived from Alaska Pollack Skins. J. Food Sci. 2021, 86, 2457–2467. [Google Scholar] [CrossRef]
- Nomura, A.; Noda, N.; Maruyama, S. Purification of Angiotensin I-Converting Enzyme Inhibitors in Pelagic Thresher Alopias Pelagicus Muscle Hydrolysate and Viscera Extracts. Fish. Sci. 2002, 68, 954–956. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Song, C.; Zhao, J.; Shi, X.; Sun, M.; Liu, J.; Fu, Y.; Jin, W.; Zhu, B. Separation and Characterization of Antioxidative and Angiotensin Converting Enzyme Inhibitory Peptide from Jellyfish Gonad Hydrolysate. Molecules 2018, 23, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-Inhibitory Peptides from Hemp Seed (Cannabis Sativa L.) Proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-J.; Yin, L.-J.; Lu, X.; Li, L.-T. Changes in Angiotensin I-Converting Enzyme Inhibitory Activities During the Ripening of Douchi (a Chinese Traditional Soybean Product) Fermented by Various Starter Cultures. Int. J. Food Prop. 2010, 13, 512–524. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, C.; Sun-Waterhouse, D.; Zhao, T.; Waterhouse, G.I.N.; Zhao, M.; Su, G. Identification of Post-Digestion Angiotensin-I Converting Enzyme (ACE) Inhibitory Peptides from Soybean Protein Isolate: Their Production Conditions and in Silico Molecular Docking with ACE. Food Chem. 2021, 345, 128855. [Google Scholar] [CrossRef] [PubMed]
- Nakano, D.; Ogura, K.; Miyakoshi, M.; Ishii, F.; Kawanishi, H.; Kurumazuka, D.; Kwak, C.; Ikemura, K.; Takaoka, M.; Moriguchi, S.; et al. Antihypertensive Effect of Angiotensin I-Converting Enzyme Inhibitory Peptides from a Sesame Protein Hydrolysate in Spontaneously Hypertensive Rats. Biosci. Biotechnol. Biochem. 2006, 70, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE inhibitory peptides derived from simulated gastrointestinal digestion in vitro of sesame (Sesamum Indicum L.) protein and molecular docking study. Int. J. Mol. Sci. 2020, 21, 1059. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, R.; Toji, K.; Kumagai, Y.; Kishimura, H. ACE Inhibitory Effect of the Protein Hydrolysates Prepared from Commercially Available Nori Product by Pepsin–Trypsin Digestion. Eur. Food Res. Technol. 2022, 248, 243–251. [Google Scholar] [CrossRef]
- Wang, C.; Tu, M.; Wu, D.; Chen, H.; Chen, C.; Wang, Z.; Jiang, L. Identification of an ACE-Inhibitory Peptide from Walnut Protein and Its Evaluation of the Inhibitory Mechanism. Int. J. Mol. Sci. 2018, 19, 1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wei, Y.; Chang, Q.; Sun, H.; Chai, K.; Huang, Z.; Zhao, Z.; Zhao, Z. Ultrafast Screening of a Novel, Moderately Hydrophilic Angiotensin-Converting-Enzyme-Inhibitory Peptide, RYL, from Silkworm Pupa Using an Fe-Doped-Silkworm-Excrement-Derived Biocarbon: Waste Conversion by Waste. J. Agric. Food Chem. 2017, 65, 11202–11211. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A Novel Angiotensin-I Converting Enzyme (ACE) Inhibitory Peptide from Gastrointestinal Protease Hydrolysate of Silkworm Pupa (Bombyx mori) Protein: Biochemical Characterization and Molecular Docking Study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef]
- Senior, R.M.; Griffin, G.L.; Mecham, R.P. Chemotactic Responses of Fibroblasts to Tropoelastin and Elastin-Derived Peptides. J. Clin. Investig. 1982, 70, 614–618. [Google Scholar] [CrossRef]
- Sato, Y.; Toyoda, T.; Shimizu-Ibuka, A.; Tamura, T.; Kobayashi-Hattori, K.; Nakamura, T.; Arai, S.; Mura, K. Novel Angiotensin I-Converting Enzyme Inhibitory Peptides Found in a Thermolysin-Treated Elastin with Antihypertensive Activity. Biosci. Biotechnol. Biochem. 2012, 76, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Maeda, I.; Kai, S.; Taniguchi, S.; Inoue, A.; Li, H.; Kesamaru, H.; Nose, T. Angiotensin I Converting Enzyme-Inhibiting Peptides Purified from Elastase-Degraded Elastin Prepared from Pig Aorta. Curr. Enzym. Inhib. 2018, 14, 67–74. [Google Scholar] [CrossRef]
- Hatakenaka, T.; Kato, T.; Okamoto, K. Novel Oligopeptides with Angiotensin I-Converting Enzyme Inhibitory Activity Found in an Elastase-Treated Hydrolysate of Porcine Aortic Elastin. In Proceedings of the 35th European Peptide Symposium, Dublin, Ireland, 26–31 August 2018; 2018; pp. 251–252. [Google Scholar]
- Miyoshi, S.; Ishikawa, H.; Kaneko, T.; Fukui, F.; Tanaka, H.; Maruyama, S. Structures and Activity of Angiotensin-Converting Enzyme Inhibitors in an α-Zein Hydrolysate. Agric. Biol. Chem. 1991, 55, 1313–1318. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure−Activity Relationship Study of Di- and Tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Kamal, H.; Mudgil, P.; Bhaskar, B.; Fisayo, A.F.; Gan, C.-Y.; Maqsood, S. Amaranth Proteins as Potential Source of Bioactive Peptides with Enhanced Inhibition of Enzymatic Markers Linked with Hypertension and Diabetes. J. Cereal Sci. 2021, 101, 103308. [Google Scholar] [CrossRef]
- Soubrier, F.; Alhenc-Gelas, F.; Hubert, C.; Allegrini, J.; John, M.; Tregear, G.; Corvol, P. Two Putative Active Centers in Human Angiotensin I-Converting Enzyme Revealed by Molecular Cloning. Proc. Natl. Acad. Sci. USA 1988, 85, 9386–9390. [Google Scholar] [CrossRef] [Green Version]
- Yathisha, U.G.; Srinivasa, M.G.; Siddappa BC, R.; Mandal, S.P.; Dixit, S.R.; Pujar, G.V.; Sheshappa, M.B. Isolation and Characterization of ACE-I Inhibitory Peptides from Ribbonfish for a Potential Inhibitor of the Main Protease of SARS-CoV-2: An in Silico Analysis. Proteins Struct. Funct. Bioinform. 2022, 90, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Geng, M.; Liu, C.; Wang, J.; Min, W.; Liu, J. Structural and Molecular Basis of Angiotensin-Converting Enzyme by Computational Modeling: Insights into the Mechanisms of Different Inhibitors. PLoS ONE 2019, 14, e0215609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrifield, R.B. Solid Phase Peptide Synthesis. I. The Synthesis of a Tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]
- Isidro-Llobet, A.; Álvarez, M.; Albericio, F. Amino Acid-Protecting Groups. Chem. Rev. 2009, 109, 2455–2504. [Google Scholar] [CrossRef] [Green Version]
- Montalbetti, C.A.G.N.; Falque, V. Amide Bond Formation and Peptide Coupling. Tetrahedron 2005, 61, 10827–10852. [Google Scholar] [CrossRef]
- Sobocińska, M.; Giełdoń, A.; Fichna, J.; Kamysz, E. 1-Substituted Sialorphin Analogues—Synthesis, Molecular Modelling and In Vitro Effect on Enkephalins Degradation by NEP. Amino Acids 2019, 51, 1201–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, K.; Ota, J.; Fujita, S.; Shiomi, Y.; Inaba, H. Construction of Ribonuclease-Decorated Artificial Virus-like Capsid by Peptide Self-Assembly. J. Org. Chem. 2020, 85, 1668–1673. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Li, X.; Yan, P.; Sun, R.; Kan, G.; Zhou, Y. Identification and Functional Mechanism of Novel Angiotensin I Converting Enzyme Inhibitory Dipeptides from Xerocomus Badius Cultured in Shrimp Processing Waste Medium. Biomed Res. Int. 2018, 2018, 5089270. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Shi, Y.; Shi, J.; Zhang, T.; Zhang, R. A Novel Angiotensin-I-Converting Enzyme (ACE) Inhibitory Peptide IAF (Ile-Ala-Phe) from Pumpkin Seed Proteins: In Silico Screening, Inhibitory Activity, and Molecular Mechanisms. Eur. Food Res. Technol. 2021, 247, 2227–2237. [Google Scholar] [CrossRef]
- Yousafi, Q.; Batool, J.; Khan, M.S.; Perveen, T.; Sajid, M.W.; Hussain, A.; Mehmood, A.; Saleem, S. In Silico Evaluation of Food Derived Bioactive Peptides as Inhibitors of Angiotensin Converting Enzyme (ACE). Int. J. Pept. Res. Ther. 2021, 27, 341–349. [Google Scholar] [CrossRef]
- Clark, A.M.; Labute, P. 2D Depiction of Protein−Ligand Complexes. J. Chem. Inf. Model. 2007, 47, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Dey, T.K.; Chatterjee, R.; Mandal, R.S.; Roychoudhury, A.; Paul, D.; Roy, S.; Pateiro, M.; Das, A.K.; Lorenzo, J.M.; Dhar, P. ACE Inhibitory Peptides from Bellamya Bengalensis Protein Hydrolysates: In Vitro and In Silico Molecular Assessment. Processes 2021, 9, 1316. [Google Scholar] [CrossRef]
- Yang, Y.J.; Mai, D.J.; Li, S.; Morris, M.A.; Olsen, B.D. Tuning Selective Transport of Biomolecules through Site-Mutated Nucleoporin-like Protein (NLP) Hydrogels. Biomacromolecules 2021, 22, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Xue, D.; Zhang, Z.; Shan, K.; Ke, W.; Zhang, M.; Zhao, D.; Nian, Y.; Xu, X.; Zhou, G.; et al. Effect of Sous-Vide Cooking on the Quality and Digestion Characteristics of Braised Pork. Food Chem. 2022, 375, 131683. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Peptide a | Purity (%) b | MS c Calcd | MS c Found |
|---|---|---|---|
| VAPGVG | 98.8 | 498.50 | 498.80 |
| VAP | 99.8 | 285.34 | 285.33 |
| VA | 95.0 | 188.23 | 188.25 |
| AP | 95.2 | 186.21 | 186.20 |
| PG | 98.3 | 172.18 | 172.20 |
| APG | 95.2 | 243.26 | 243.20 |
| GAP | 99.0 | 243.26 | 243.20 |
| VSP | 99.8 | 301.34 | 301.20 |
| TRP | 90.4 | 372.42 | 372.30 |
| GVG | 98.9 | 231.25 | 231.20 |
| GLG | 97.8 | 245.28 | 245.20 |
| Peptide a | IC50 (μM) | S (kcal/mol) |
|---|---|---|
| VAP | 1.0 | −10.3512 |
| VA | 326 | −9.1838 |
| AP | 230 | −8.5065 |
| PG | 1814 | −8.9506 |
| APG | 1194 | −9.3226 |
| GAP | 35.0 | −9.3464 |
| VSP | 10.4 | −10.0776 |
| TRP | 1.3 | −11.1199 |
| LSP b | 1.7 d | −10.2541 |
| VGP b | 26.3 d | −9.9914 |
| VLP c | 3.9 e | −10.9502 |
| GVG | 369 | −9.0309 |
| GLG | 485 | −9.4905 |
| Peptide | Toxicity a | The Number of Peptide in Tropoelastin b | Start Location b | Net Charge at pH 8.3 c | pI c | PepSite2 p Value d |
|---|---|---|---|---|---|---|
| VAP | Non-Toxin | 6 | 492, 498, 504, 510, 516, 690 | −0.7 | 5.50 | 0.000028 |
| GAP | Non-Toxin | 5 | 249, 283, 306, 468, 531 | −0.7 | 5.50 | 0.000063 |
| VSP | Non-Toxin | 2 | 370, 659 | −0.7 | 5.50 | 0.000130 |
| TRP | Non-Toxin | 1 | 682 | 0.3 | 10.00 | 0.000056 |
| LSP | Non-Toxin | 2 | 421, 698 | −0.7 | 5.50 | 0.000167 |
| VGP | Non-Toxin | 2 | 173, 620 | −0.7 | 5.50 | 0.000146 |
| VLP | Non-Toxin | 3 | 133, 146, 255 | −0.7 | 5.50 | 0.000221 |
| GVG | Non-Toxin | 39 | 40, 89, 92, 110, 122, 143, 172, 230, 258, 260, 263, 313, 318, 323, 328, 333, 338, 343, 348, 353, 358, 363, 390, 392, 400, 410, 412, 466, 495, 501, 507, 513, 562, 571, 604, 619, 635, 638, 643 | −0.7 | 5.50 | 0.008118 |
| GLG | Non-Toxin | 11 | 21, 24, 48, 52, 86, 95, 485, 641, 646, 674, 709 | −0.7 | 5.50 | 0.002949 |
| Peptide | State | Docking Score S (kcal/mol) | Amino Acid Residues and Zn(II) in the ACE Active Pocket | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ala356 | Asp358 | Pro407 | Glu411 * | His387 * | Asn70 | His353 | Zn(II)701 | |||
| TRP | 1 | −11.1199 | ✓✓ | ✓ | ○○○○ | |||||
| 2 | −11.0033 | ✓✓ | ✓ | ✓ | ✓◎ | ○○○○ | ||||
| 3 | −10.8337 | ✓ | ◎ | ◎◎◎◎ | ○○○○ | |||||
| 4 | −10.7977 | ✓ | ✓✓◎◎ | ◎ | ○○○○ | |||||
| APG | 1 | −9.3226 | ✓✓ | ✓ | ○○○○ | |||||
| 2 | −9.2340 | ✓ | ✓ | ○○○○ | ||||||
| 3 | −9.1327 | ✓ | ○○○○ | |||||||
| 4 | −9.1070 | ✓ | ○○○ | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatakenaka, T.; Kato, T.; Okamoto, K. In Vitro and In Silico Studies on Angiotensin I-Converting Enzyme Inhibitory Peptides Found in Hydrophobic Domains of Porcine Elastin. Molecules 2023, 28, 3337. https://doi.org/10.3390/molecules28083337
Hatakenaka T, Kato T, Okamoto K. In Vitro and In Silico Studies on Angiotensin I-Converting Enzyme Inhibitory Peptides Found in Hydrophobic Domains of Porcine Elastin. Molecules. 2023; 28(8):3337. https://doi.org/10.3390/molecules28083337
Chicago/Turabian StyleHatakenaka, Toshiya, Tamaki Kato, and Kouji Okamoto. 2023. "In Vitro and In Silico Studies on Angiotensin I-Converting Enzyme Inhibitory Peptides Found in Hydrophobic Domains of Porcine Elastin" Molecules 28, no. 8: 3337. https://doi.org/10.3390/molecules28083337
APA StyleHatakenaka, T., Kato, T., & Okamoto, K. (2023). In Vitro and In Silico Studies on Angiotensin I-Converting Enzyme Inhibitory Peptides Found in Hydrophobic Domains of Porcine Elastin. Molecules, 28(8), 3337. https://doi.org/10.3390/molecules28083337