
Dendrobium Nobile Alcohol Extract Extends the Lifespan of Caenorhabditis elegans via hsf-1 and daf-16

 , ,
, ,
Abstract
:1. Introduction
2. Results
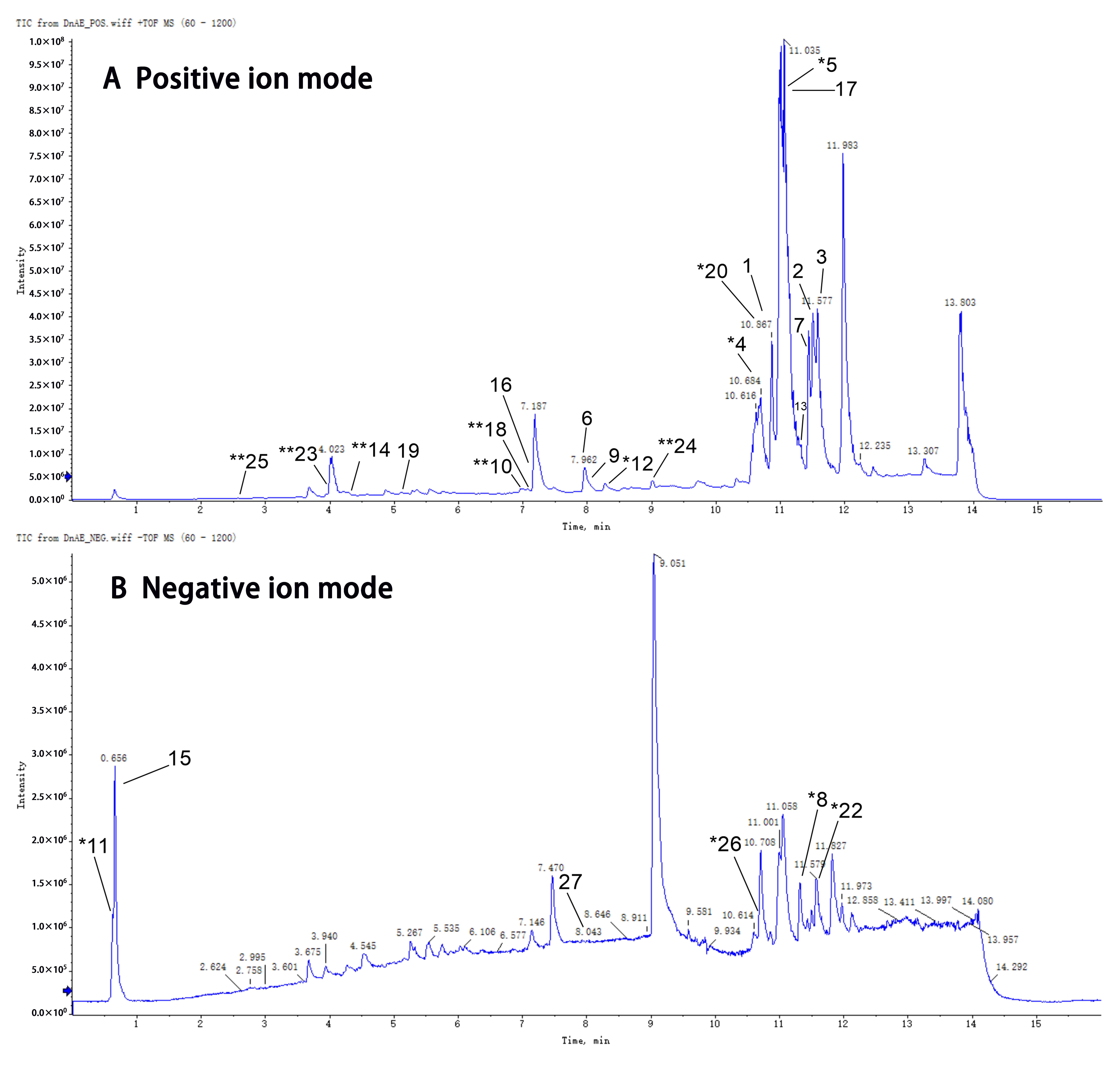
2.1. Chemical Composition of DnAE
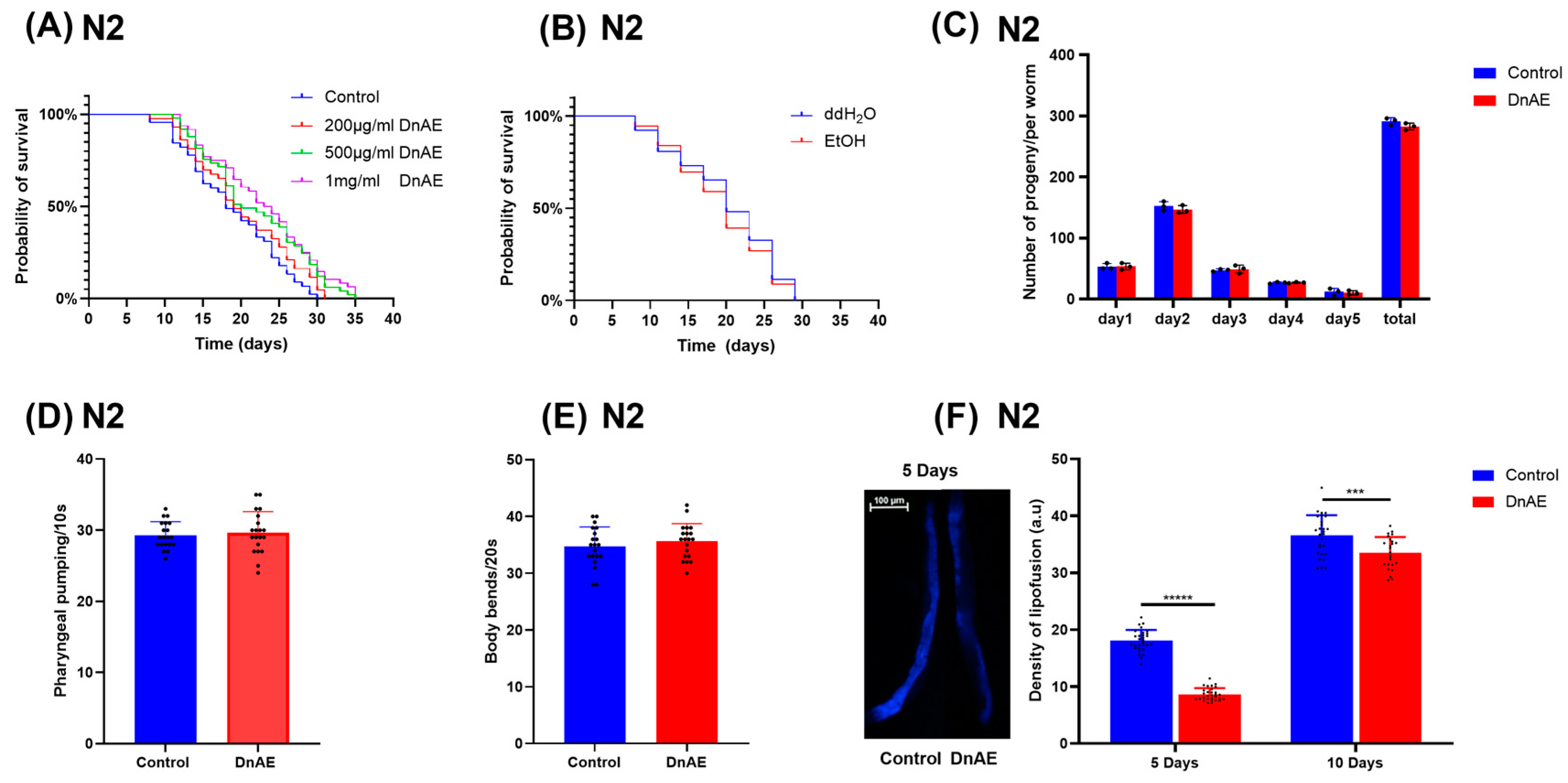
2.2. DnAE Increases the Lifespan and Improves the Health Span of C. elegans
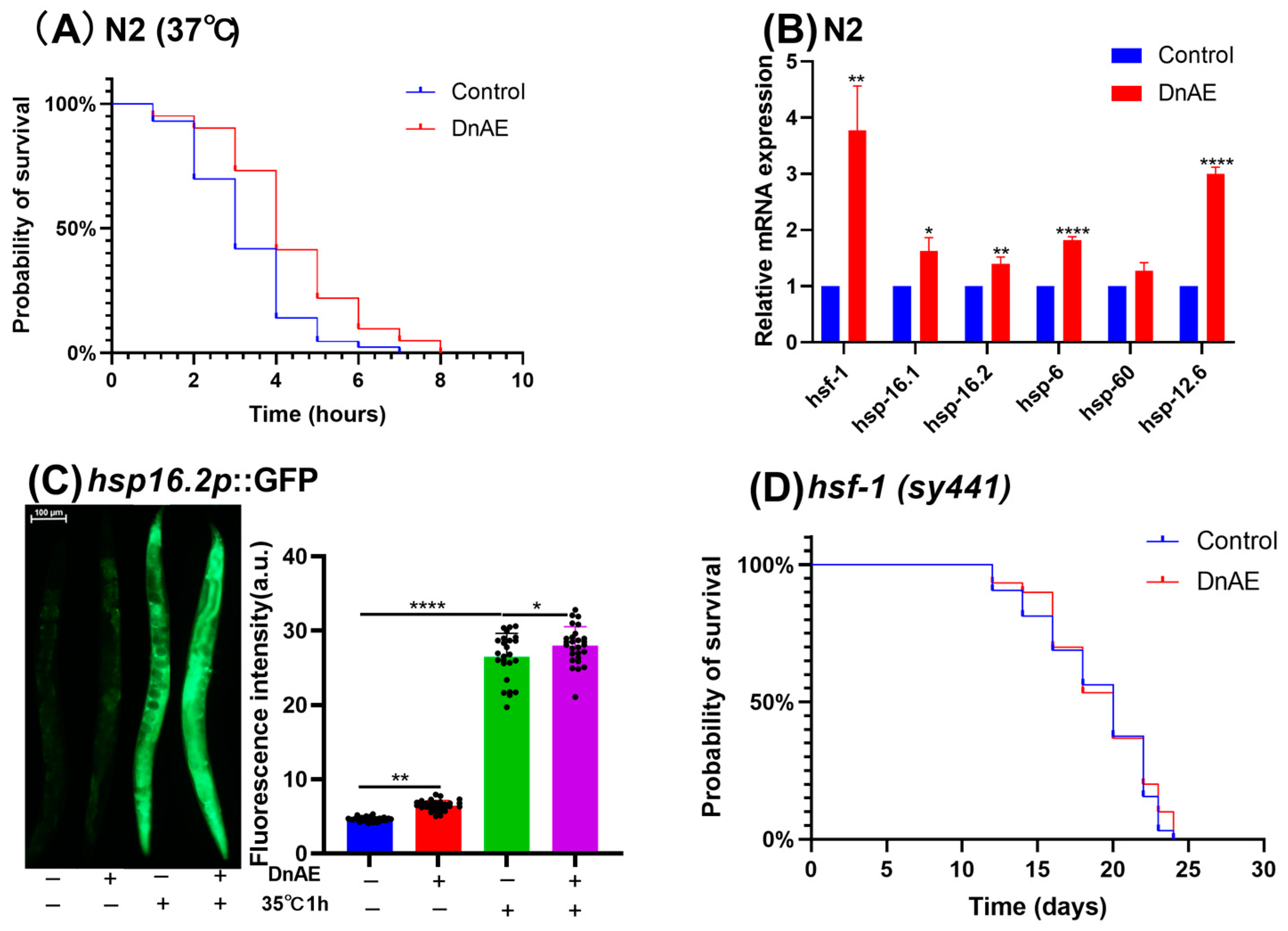
2.3. DnAE Increased Heat Stress Tolerance by Upregulating the Expression of hsf-1 and HSPs
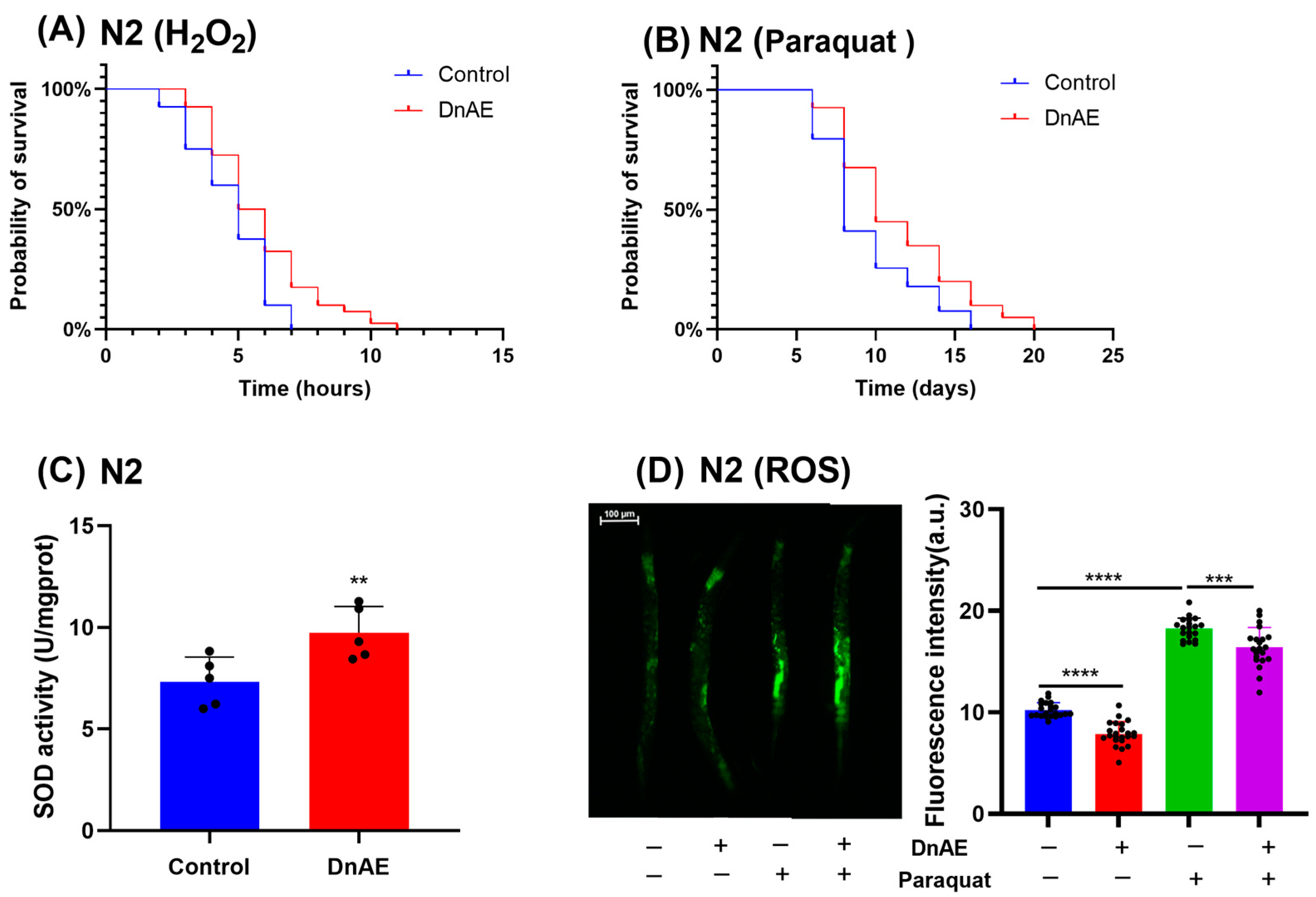
2.4. DnAE Enhances Antioxidant Activity in C. elegans
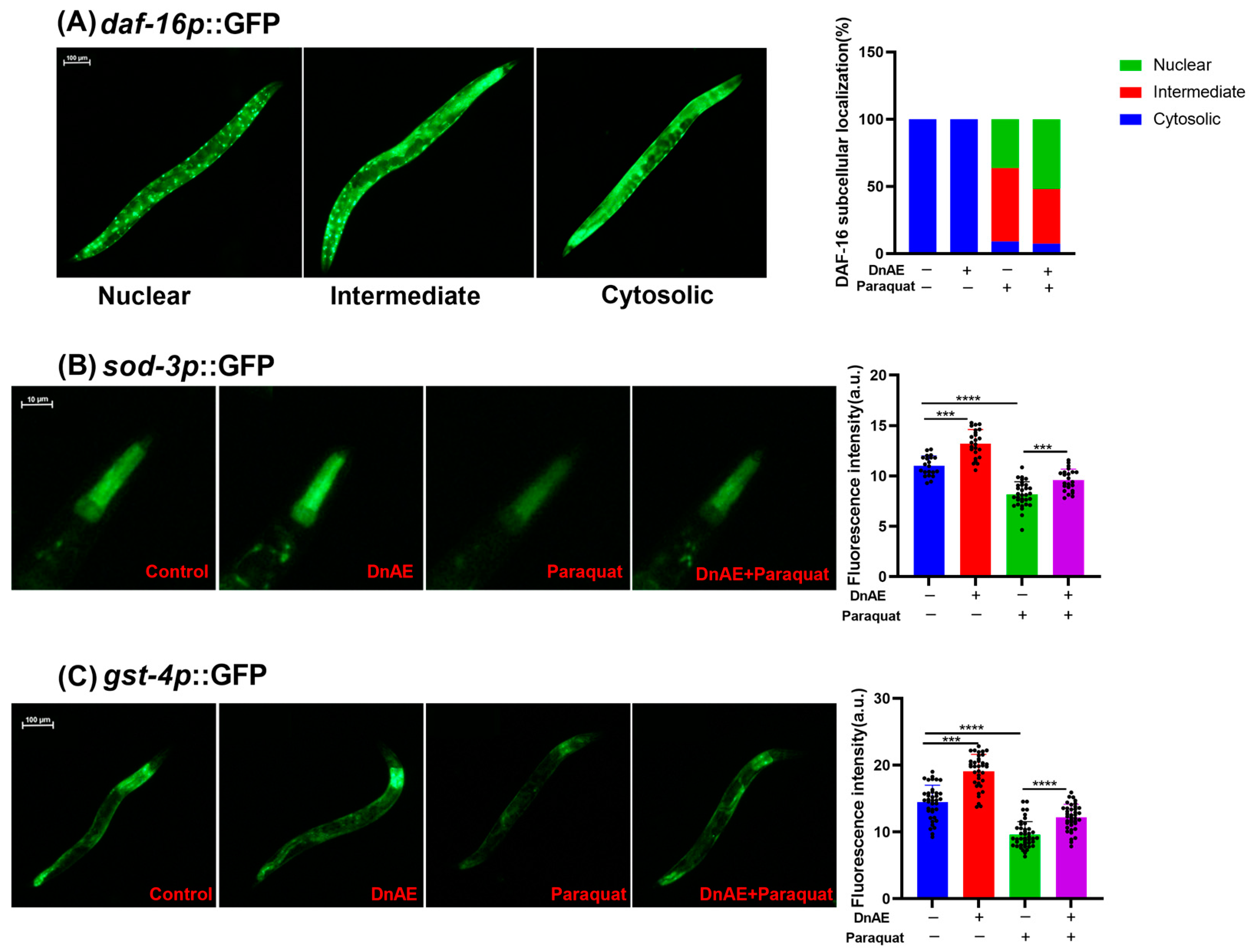
2.5. DnAE Promotes the Nuclear Localization of DAF-16 and the Expression of SOD-3 and GST-4
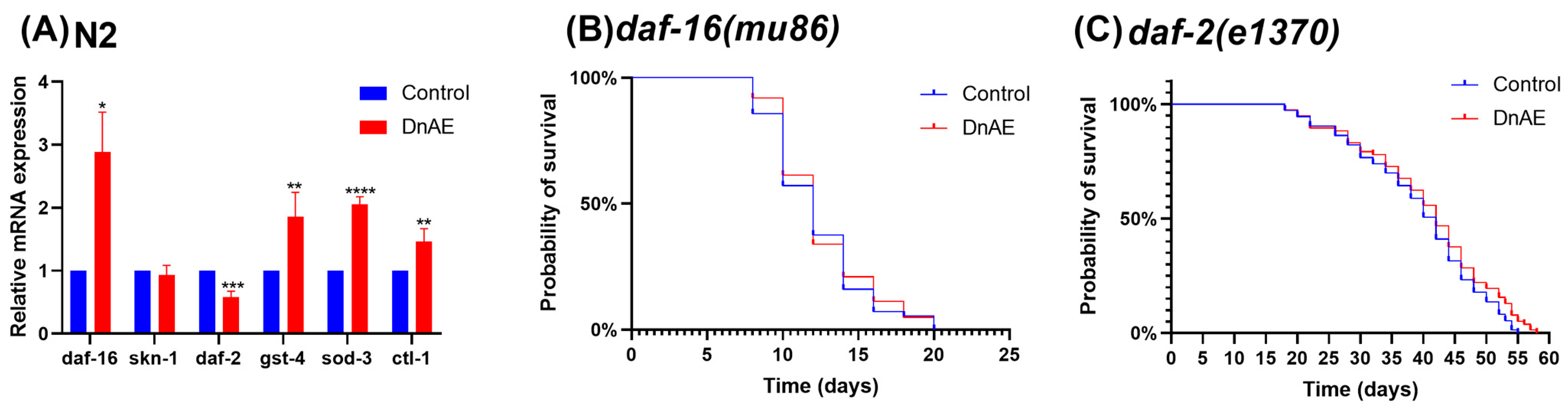
2.6. DnAE-Mediated Lifespan Extension in C. elegans Is Dependent on DAF-16
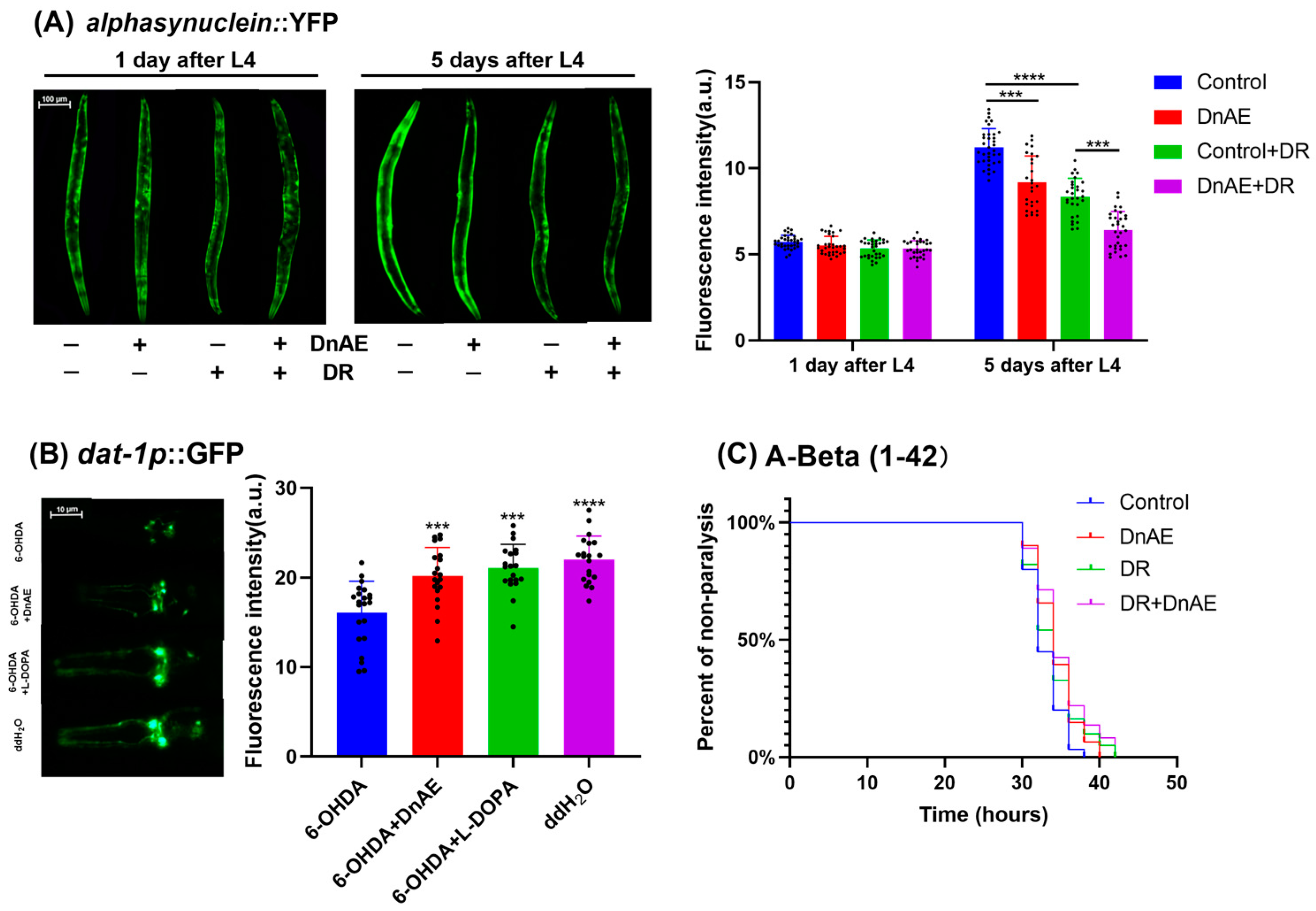
2.7. DnAE Decreases the Progression of Aging-Related Diseases
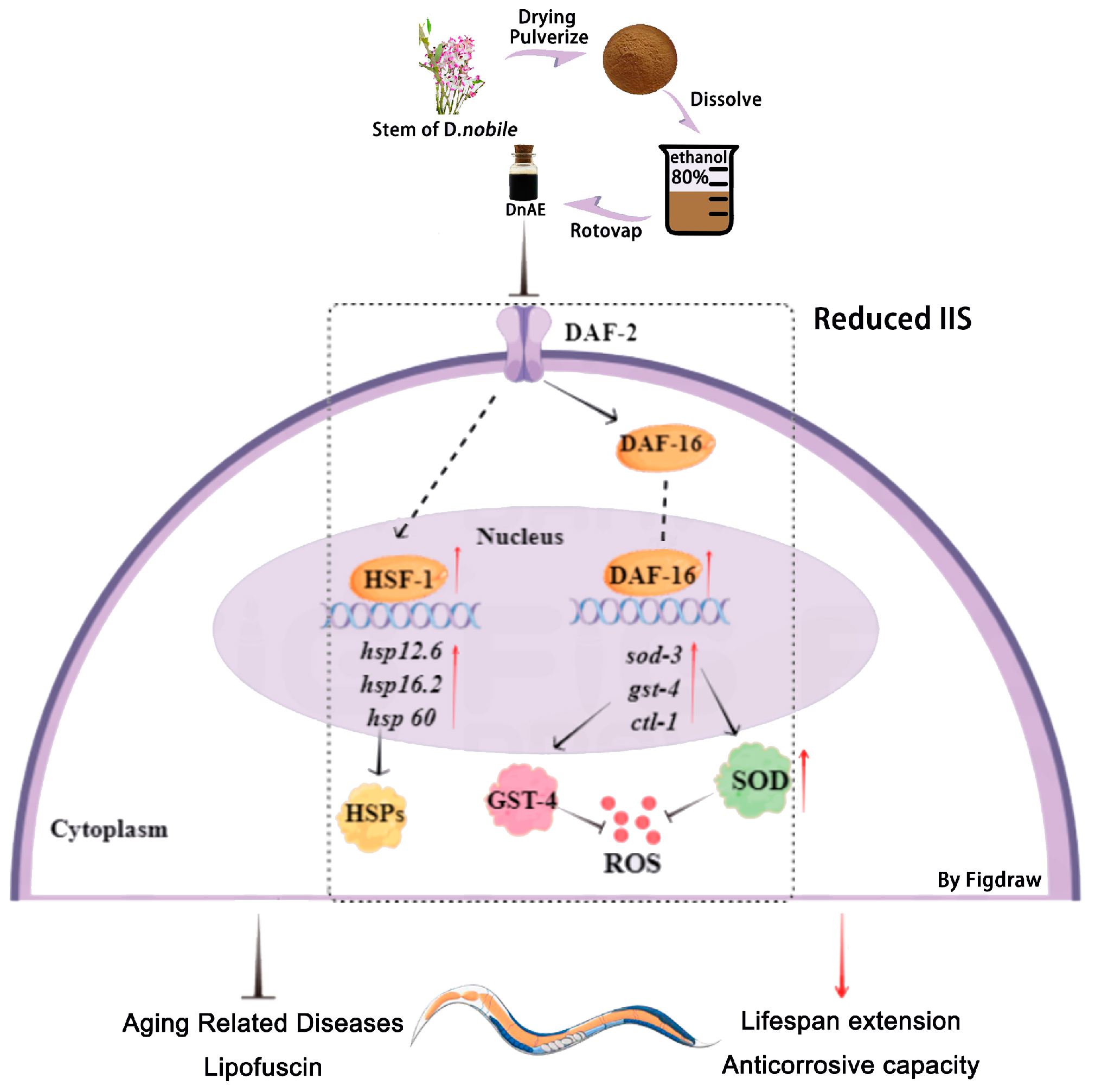
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and DnAE Preparation
5.2. UHPLC-Q-TOF-MS/MS Compound Identification
5.3. Lifespan Assay
5.4. Reproduction Assay
5.5. Bending Assay
5.6. Pharyngeal Pumping Assay
5.7. Oxidative Stress Resistance Assay
5.8. Chronic Oxidative Stress Resistance Assay
5.9. Thermal Stress Resistance Assay
5.10. Determination of Lipofuscin
5.11. Determination of ROS Levels
5.12. Determination of SOD Activity
5.13. Quantitative Analysis of Fluorescence Intensity
5.14. Nuclear Localization of DAF-16
5.15. Paralysis Assay
5.16. Quantitative Real-Time PCR Assay
5.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrucci, L.; Gonzalez-Freire, M.; Fabbri, E.; Simonsick, E.; Tanaka, T.; Moore, Z.; Salimi, S.; Sierra, F.; de Cabo, R. Measuring biological aging in humans: A quest. Aging Cell 2020, 19, 21. [Google Scholar] [CrossRef] [PubMed]
- North, B.J.; Sinclair, D.A. The Intersection Between Aging and Cardiovascular Disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Lautrup, S.; Sinclair, D.A.; Mattson, M.P.; Fang, E.F. NAD+ in Brain Aging and Neurodegenerative Disorders. Cell Metab. 2019, 30, 630–655. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.B.; Ke, H.M.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol Ameliorates Aging-Related Metabolic Phenotypes by Inhibiting cAMP Phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef]
- Wang, H.L.; Liu, J.; Li, T.; Liu, R.H. Blueberry extract promotes longevity and stress tolerance via DAF-16 in Caenorhabditis elegans. Food Funct. 2018, 9, 5273–5282. [Google Scholar] [CrossRef]
- Yu, X.X.; Li, H.; Lin, D.F.; Guo, W.Z.; Xu, Z.H.; Wang, L.P.; Guan, S.W. Ginsenoside Prolongs the Lifespan of C. elegans via Lipid Metabolism and Activating the Stress Response Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 9668. [Google Scholar] [CrossRef]
- Song, S.B.; Shim, W.; Hwang, E.S. Lipofuscin Granule Accumulation Requires Autophagy Activation. Mol. Cells 2023, 46, 486–495. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, J.Y.; Wong, J.H.; Ye, X.J.; Sze, S.C.W.; Tong, Y.; Zhang, K.Y. Review of research on Dendrobium, a prized folk medicine. Appl. Microbiol. Biotechnol. 2012, 93, 1795–1803. [Google Scholar] [CrossRef]
- Zhao, R.X.; Zheng, S.G.; Li, Y.; Zhang, X.Q.; Rao, D.; Chun, Z.; Hu, Y.D. As a novel anticancer candidate, ether extract of Dendrobium nobile overstimulates cellular protein biosynthesis to induce cell stress and autophagy. J. Appl. Biomed. 2023, 21, 23–35. [Google Scholar] [CrossRef]
- Lei, X.C.; Huo, P.; Xie, Y.J.; Wang, Y.H.; Liu, G.H.; Tu, H.Y.; Shi, Q.X.; Mo, Z.C.; Zhang, S. Dendrobium nobile Lindl polysaccharides improve testicular spermatogenic function in streptozotocin-induced diabetic rats. Mol. Reprod. Dev. 2022, 89, 202–213. [Google Scholar] [CrossRef]
- Hsu, W.H.; Chung, C.P.; Wang, Y.Y.; Kuo, Y.H.; Yeh, C.H.; Lee, I.J.; Lin, Y.L. Dendrobium nobile protects retinal cells from UV-induced oxidative stress damage via Nrf2/HO-1 and MAPK pathways. J. Ethnopharmacol. 2022, 288, 10. [Google Scholar] [CrossRef]
- Li, D.D.; Zheng, C.Q.; Zhang, F.; Shi, J.S. Potential neuroprotection by Dendrobium nobile Lindl alkaloid in Alzheimer’s disease models. Neural Regen. Res. 2022, 17, 972–977. [Google Scholar]
- Xu, Y.Y.; Xu, Y.S.; Wang, Y.; Wu, Q.; Lu, Y.F.; Liu, J.; Shi, J.S. Dendrobium nobile Lindl. alkaloids regulate metabolism gene expression in livers of mice. J. Pharm. Pharmacol. 2017, 69, 1409–1417. [Google Scholar] [CrossRef]
- Yang, S.; Gong, Q.H.; Wu, Q.; Li, F.; Lu, Y.F.; Shi, J.S. Alkaloids enriched extract from Dendrobium nobile Lindl. attenuates tau protein hyperphosphorylation and apoptosis induced by lipopolysaccharide in rat brain. Phytomedicine 2014, 21, 712–716. [Google Scholar] [CrossRef]
- Yoo, S.-R.; Jeong, S.-J.; Lee, N.-R.; Shin, H.-K.; Seo, C.-S. Simultaneous determination and anti-inflammatory effects of four phenolic compounds in Dendrobii Herba. Nat. Prod. Res. 2017, 31, 2923–2926. [Google Scholar] [CrossRef]
- Vayndorf, E.M.; Lee, S.S.; Liu, R.H. Whole apple extracts increase lifespan, healthspan and resistance to stress in Caenorhabditis elegans. J. Funct. Foods 2013, 5, 1235–1243. [Google Scholar] [CrossRef]
- Gao, X.P.; Liu, J.; Luo, Y.N.; Lei, Y.; Long, W.W.; Wang, K.; Zhou, J.; Lei, M.; Yang, N.; Zou, H.Y.; et al. Various Fractions of Alcoholic Extracts from Dendrobium nobile Functionalized Antioxidation and Antiaging in D-Galactose-Induced Aging Mice. Front. Biosci. Landmark 2022, 27, 315. [Google Scholar] [CrossRef]
- Martins, R.; Lithgow, G.J.; Link, W. Long live FOXO: Unraveling the role of FOXO proteins in aging and longevity. Aging Cell 2016, 15, 196–207. [Google Scholar] [CrossRef]
- Kwon, H.C.; Bae, Y.; Lee, S.V. The Role of mRNA Quality Control in the Aging of Caenorhabditis elegans. Mol. Cells 2023, 46, 664–671. [Google Scholar] [CrossRef]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 464, 504–512. [Google Scholar] [CrossRef]
- Henderson, S.T.; Johnson, T.E. daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Curr. Biol. 2001, 11, 1975–1980. [Google Scholar] [CrossRef]
- Harada, C.N.; Love, M.C.N.; Triebel, K.L. Normal Cognitive Aging. Clin. Geriatr. Med. 2013, 29, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, J.; McGhee, J.D. Recent Advances in Understanding the Molecular Mechanisms Regulating C. elegans Transcription. Dev. Dyn. 2010, 239, 1388–1404. [Google Scholar] [CrossRef]
- Prahlad, V.; Cornelius, T.; Morimoto, R.I. Regulation of the cellular heat shock response in Caenorhabditis elegans by thermosensory neurons. Science 2008, 320, 811–814. [Google Scholar] [CrossRef]
- Song, X.M.; Wang, H.X.; Ma, C.Y.; Kou, X.R.; Jia, Q.H. Analysis on volatile components from flower of Dendrobium nobile Lindl. by GC-MS. J. Food Sci. Biotechnol. 2019, 38, 133–138. [Google Scholar]
- Wang, Y.H.; Avula, B.; Abe, N.; Wei, F.; Wang, M.; Ma, S.C.; Ali, Z.; Elsohly, M.; Khan, I. Tandem mass spectrometry for structural identification of Sesquiterpene Alkaloids from the stems of Dendrobium nobile using LC-Q-TOF. Planta Med. 2016, 82, 662–670. [Google Scholar] [PubMed]
- Lu, A.-J.; Jiang, Y.; Wu, J.; Tan, D.-p.; Qin, L.; Lu, Y.-l.; Qian, Y.; Bai, C.-j.; Yang, J.-y.; Ling, H.; et al. Opposite trends of glycosides and alkaloids in Dendrobium nobile of different age based on UPLC-Q/TOF-MS combined with multivariate statistical analyses. Phytochem. Anal. 2022, 33, 619–634. [Google Scholar] [CrossRef]
- Ye, Q.H.; Qin, G.W.; Zhao, W.M. Immunomodulatory sesquiterpene glycosides from Dendrobium nobile. Phytochemistry 2002, 61, 885–890. [Google Scholar] [CrossRef]
- Okamoto, T.; Natsume, M.; Onaka, T.; Uchimaru, F.; Shimizu, M. The structure of dendroxine. The third alkaloid from Dendrobium nobile. Chem. Pharm. Bull. 1966, 14, 672–675. [Google Scholar] [CrossRef]
- Xia, J.; Yang, Z.; Zeng, Q.F.; Liang, Y.; Hao, X.Y.; Zhou, W. Analysis of chemical constituents in Dendrobium nobile by UPLC-Q-TOF. J. Chin. Med. Mater. 2018, 41, 600–607. [Google Scholar]
- Wang, D.F.; Chou, G.X.; Zhao, N.Y.; Zhang, T.; Xu, H. Study on chemical constituents in stems of Dendrobium nobile. Chin. Tradit. Herb. Drugs 2012, 43, 1492–1495. [Google Scholar]
- Wan, Q.-L.; Meng, X.; Fu, X.; Chen, B.; Yang, J.; Yang, H.; Zhou, Q. Intermediate metabolites of the pyrimidine metabolism pathway extend the lifespan of C. elegans through regulating reproductive signals. Aging-US 2019, 11, 3993–4010. [Google Scholar] [CrossRef]
- Park, J.-W.; Jeong, J.; Bae, Y.-S. Protein Kinase CK2 Is Upregulated by Calorie Restriction and Induces Autophagy. Mol. Cells 2022, 45, 112–121. [Google Scholar] [CrossRef]
- Papaevgeniou, N.; Hoehn, A.; Grune, T.; Chondrogianni, N. Lipofuscin effects in Caenorhabditis elegans ageing model. Free Radic. Biol. Med. 2017, 108, S48. [Google Scholar] [CrossRef]
- Fasnacht, M.; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front. Mol. Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Webb, A.E. Neuronal functions of FOXO/DAF-16. Nutr. Healthy Aging 2017, 4, 113–126. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhang, W.H.; Zhang, P.; Li, Q.; Sun, Y.; Wang, J.W.; Zhang, S.B.O.; Cai, T.; Zhan, C.; Dong, M.Q. Intestine-specific removal of DAF-2 nearly doubles lifespan in Caenorhabditis elegans with little fitness cost. Nat. Commun. 2022, 13, 6339. [Google Scholar] [PubMed]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed]
- van Ham, T.J.; Thijssen, K.L.; Breitling, R.; Hofstra, R.M.; Plasterk, R.H.; Nollen, E.A. C. elegans model identifies genetic modifiers of alpha-synuclein inclusion formation during aging. PLoS Genet. 2008, 4, e1000027. [Google Scholar] [CrossRef]
- Steinkraus, K.A.; Smith, E.D.; Davis, C.; Carr, D.; Pendergrass, W.R.; Sutphin, G.L.; Kennedy, B.K.; Kaeberlein, M. Dietary restriction suppresses proteotoxicity and enhances longevity by an hsf-1-dependent mechanism in Caenorhabditis elegans. Aging Cell 2008, 7, 394–404. [Google Scholar] [CrossRef]
- Blesa, J.; Przedborski, S. Parkinson’s disease: Animal models and dopaminergic cell vulnerability. Front. Neuroanat. 2014, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.; Link, C.D.; Butterfield, D.A. Oxidative stress precedes fibrillar deposition of Alzheimer’s disease amyloid beta-peptide (1-42) in a transgenic Caenorhabditis elegans model. Neurobiol. Aging 2003, 24, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-M.; Li, X.; Su, S.-Q.; Wang, Y.-T.; Xu, T.; Zha, X.-Q.; Pan, L.-H.; Shang, Z.-Z.; Zhang, F.-Y.; Luo, J.-P. Dendrobine inhibits dopaminergic neuron apoptosis via MANF-mediated ER stress suppression in MPTP/MPP+-induced Parkinson’s disease models. Phytomedicine 2022, 102, 154193. [Google Scholar]
- Chen, H.; Tu, M.; Liu, S.; Wen, Y.; Chen, L. Dendrobine Alleviates Cellular Senescence and Osteoarthritis via the ROS/NF-κB Axis. Int. J. Mol. Sci. 2023, 24, 2365. [Google Scholar] [CrossRef]
- Zhang, C.-C.; Kong, Y.-L.; Zhang, M.-S.; Wu, Q.; Shi, J.-S. Two new alkaloids from Dendrobium nobile Lindl. exhibited neuroprotective activity, and dendrobine alleviated Aβ1-42-induced apoptosis by inhibiting CDK5 activation in PC12 cells. Drug Dev. Res. 2023, 84, 262–274. [Google Scholar] [CrossRef]
- Li, Y.; Li, F.; Gong, Q.; Wu, Q.; Shi, J. Inhibitory Effects of DendrobiumAlkaloids on Memory Impairment Induced by Lipopolysaccharide in Rats. Planta Medica 2011, 77, 117–121. [Google Scholar] [CrossRef]
- Sales-Campos, H.; de Souza, P.R.; Peghini, B.C.; da Silva, J.S.; Cardoso, C.R. An Overview of the Modulatory Effects of Oleic Acid in Health and Disease. Mini-Rev. Med. Chem. 2013, 13, 201–210. [Google Scholar] [PubMed]
- Cabreiro, F.; Au, C.; Leung, K.Y.; Vergara-Irigaray, N.; Cocheme, H.M.; Noori, T.; Weinkove, D.; Schuster, E.; Greene, N.D.E.; Gems, D. Metformin Retards Aging in C. elegans by Altering Microbial Folate and Methionine Metabolism. Cell 2013, 153, 228–239. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Yu, W. Heat shock proteins and viral infection. Front. Immunol. 2022, 13, 11. [Google Scholar] [CrossRef]
- Li, Z.R.; Xiang, J.; Hu, D.Y.; Song, B.A. Naturally potential antiviral agent polysaccharide from Dendrobium nobile Lindl. Pest. Biochem. Physiol. 2020, 167, 9. [Google Scholar] [CrossRef]
- Luo, A.X.; He, X.J.; Zhou, S.; Fan, Y.J.; Luo, A.S.; Chun, Z. Purification, composition analysis and antioxidant activity of the polysaccharides from Dendrobium nobile Lindl. Carbohydr. Polym. 2010, 79, 1014–1019. [Google Scholar] [CrossRef]
- Suzuki, T.; Takahashi, J.; Yamamoto, M. Molecular Basis of the KEAP1-NRF2 Signaling Pathway. Mol. Cells 2023, 46, 133–141. [Google Scholar] [CrossRef]
- Tullet, J.M.A.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef]
- Li, S.Y.; Zhou, J.X.; Xu, S.F.; Li, J.; Liu, J.; Lu, Y.F.; Shi, J.S.; Zhou, S.Y.; Wu, Q. Induction of Nrf2 pathway by Dendrobium nobile Lindl. alkaloids protects against carbon tetrachloride induced acute liver injury. Biomed. Pharmacother. 2019, 117, 8. [Google Scholar] [CrossRef]
- Zhou, J.X.; Zhang, Y.; Li, S.Y.; Zhou, Q.; Lu, Y.F.; Shi, J.S.; Liu, J.; Wu, Q.; Zhou, S.Y. Dendrobium nobile Lindl. alkaloids-mediated protection against CCl4-induced liver mitochondrial oxidative damage is dependent on the activation of Nrf2 signaling pathway. Biomed. Pharmacother. 2020, 129, 10. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, S.-J.V. Recent Progress in Regulation of Aging by Insulin/ IGF-1 Signaling in Caenorhabditis elegans. Mol. Cells 2022, 45, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Hong, M. Aqueous extract of Dendrobium officinale confers neuroprotection against hypoxic-ischemic brain damage in neonatal rats. Kaohsiung J. Med. Sci. 2020, 36, 43–53. [Google Scholar] [CrossRef]
- Li, D.-d.; Wang, G.-q.; Wu, Q.; Shi, J.-S.; Zhang, F. Dendrobium nobile Lindl alkaloid attenuates 6-OHDA-induced dopamine neurotoxicity. Biotechnol. Appl. Biochem. 2021, 68, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.Y.; Yang, Z.S.; Zhao, N.H.; Li, Z.W.; Cao, X. Gastrodin protects dopaminergic neurons via insulin-like pathway in a Parkinson’s disease model. BMC Neurosci. 2019, 20, 11. [Google Scholar] [CrossRef]
- Wang, F.R.; Jin, T.; Li, H.Y.; Long, H.F.; Liu, Y.; Jin, S.; Lu, Y.Y.; Peng, Y.H.; Liu, C.; Zhao, L.H.; et al. Cannabidivarin alleviates alpha-synuclein aggregation via DAF-16 in Caenorhabditis elegans. FASEB J. 2023, 37, 14. [Google Scholar]
- Wang, X.X.; Cao, M.; Dong, Y.Q. Royal jelly promotes DAF-16-mediated proteostasis to tolerate beta-amyloid toxicity in C. elegans model of Alzheimer’s disease. Oncotarget 2016, 7, 54183–54193. [Google Scholar] [CrossRef]
- Fu, Y.; Yu, Y.; Tan, H.; Wang, B.; Peng, W.H.; Sun, Q. Metabolomics reveals dopa melanin involved in the enzymatic browning of the yellow cultivars of East Asian golden needle mushroom (Flammulina filiformis). Food Chem. 2022, 370, 12. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Leiers, B.; Kampkötter, A.; Grevelding, C.G.; Link, C.D.; Johnson, T.E.; Henkle-Dührsen, K. A stress-responsive glutathione S-transferase confers resistance to oxidative stress in Caenorhabditis elegans. Free Radic. Biol. Med. 2003, 34, 1405–1415. [Google Scholar] [CrossRef]
- Lin, K.; Dorman, J.B.; Rodan, A.; Kenyon, C. daf-16: An HNF-3/forkhead family member that can function to double the life-span of Caenorhabditis elegans. Science 1997, 278, 1319–1322. [Google Scholar] [CrossRef]
- Li, J.; Le, W. Modeling neurodegenerative diseases in Caenorhabditis elegans. Exp. Neurol. 2013, 250, 94–103. [Google Scholar] [CrossRef]
- Motta, H.S.; Roos, D.; Tabarelli, G.; Rodrigues, O.E.D.; Ávila, D.; Quines, C.B. Activation of SOD-3 is involved in the antioxidant effect of a new class of β-aryl-chalcogenium azide compounds in Caenorhabditis elegans. An. Acad. Bras. Cienc. 2020, 92 (Suppl. 2), e20181147. [Google Scholar] [CrossRef]
- Yang, Z.Z.; Yu, Y.T.; Lin, H.R.; Liao, D.C.; Cui, X.H.; Wang, H.B. Lonicera japonica extends lifespan and healthspan in Caenorhabditis elegans. Free Radic. Biol. Med. 2018, 129, 310–322. [Google Scholar] [CrossRef]
- Liao, V.H.C.; Yu, C.W.; Chu, Y.J.; Li, W.H.; Hsieh, Y.C.; Wang, T.T. Curcumin-mediated lifespan extension in Caenorhabditis elegans. Mech. Ageing Dev. 2011, 132, 480–487. [Google Scholar] [CrossRef]
- Xu, C.; Liu, S.; Liu, Z.Q.; Song, F.R.; Liu, S.Y. Superoxide generated by pyrogallol reduces highly water-soluble tetrazolium salt to produce a soluble formazan: A simple assay for measuring superoxide anion radical scavenging activities of biological and abiological samples. Anal. Chim. Acta 2013, 793, 53–60. [Google Scholar] [CrossRef]
- Senchuk, M.M.; Dues, D.J.; Scheer, C.E.; Johnson, B.K.; Madaj, Z.B.; Bowman, M.J.; Winn, M.E.; Van Reamsdonk, J.M. Activation of DAF-16/FOX0 by reactive oxygen species contributes to longevity in long-lived mitochondrial mutants in Caenorhabditis elegans. PLoS Genet. 2018, 14, e1007268. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Compounds | Adduct | Formula | m/z | Retention Time (Min) | Peak Area | Reference |
|---|---|---|---|---|---|---|---|
| 1 | Palmitic amide ** | [M+H]+ | C16H33NO | 256.2637 | 10.865 | 75,870,888 | |
| 2 | C2 ** | [M−H2O+H]+ | C44H86NO7P | 284.2942 | 11.453 | 74,055,912 | |
| 3 | Glyceryl monostearate ** | [M+H]+ | C21H42O4 | 359.3146 | 11.586 | 19,812,174 | |
| 4 | Linoleic acid * | [M+H−H2O]+ | C18H32O2 | 263.2356 | 10.686 | 11,779,023 | [25] |
| 5 | 2-Palmitoylglycerol * | [M+H]+ | C19H38O4 | 331.2829 | 11.069 | 4,608,534 | |
| 6 | C6 ** | [M+Na]+ | C24H30O6 | 437.1923 | 7.965 | 3,485,238 | |
| 7 | Triethanolamine ** | [M+H]+ | C6H15NO3 | 150.1106 | 11.376 | 2,495,882 | |
| 8 | Palmitic acid * | [M-H]− | C16H32O2 | 255.2328 | 11.324 | 2,262,816 | |
| 9 | C9 ** | [M+ACN+H]+ | C22H30O6 | 432.2376 | 7.965 | 1,424,730 | |
| 10 | Mubironine B ** | [M+H]+ | C15H23NO2 | 250.1789 | 7.038 | 1,252,497 | [26] |
| 11 | Quinic acid * | [M−H]− | C7H12O6 | 191.0573 | 0.617 | 897,605 | [27] |
| 12 | 2-Aminooctadecane-1,3,4-triol * | [M+H]+ | C18H39NO3 | 318.2985 | 8.293 | 759,759 | |
| 13 | C13 ** | [M+H]+ | C22H39NO | 334.3099 | 11.38 | 707,127 | |
| 14 | Dendroside G ** | [M+Na]+ | C21H34O10 | 469.2029 | 4.291 | 626,858 | [28] |
| 15 | Sucrose ** | [M−H]− | C12H22O11 | 341.1098 | 0.659 | 613,066 | |
| 16 | C16 ** | [M+H]+ | C21H33N3O3 | 376.2586 | 7.091 | 608,355 | |
| 17 | Glyceryl palmitate ** | [M+NH4]+ | C19H38O4 | 348.307 | 11.069 | 439,586 | |
| 18 | Dendroxine ** | [M+H]+ | C17H25NO3 | 292.1917 | 7.07 | 436,709 | [29] |
| 19 | C19 ** | [M+Na]+ | C21H32O9 | 451.1933 | 5.221 | 397,304 | |
| 20 | Dendrobine * | [M+H]+ | C16H25NO2 | 264.1934 | 10.865 | 376,165 | [30] |
| 21 | 13′-Hydroxy-α-tocopherol | [M+H−H2O]+ | C29H50O3 | 429.3703 | 11.453 | 230,873 | |
| 22 | Oleic acid * | [M-H]− | C18H34O2 | 281.2469 | 11.586 | 188,108 | |
| 23 | Nobilonine ** | [M+H]+ | C17H27NO3 | 294.2039 | 4.174 | 157,383 | [26] |
| 24 | Dibutyl phthalate ** | [M+H]+ | C16H22O4 | 279.1582 | 8.906 | 127,790 | [31] |
| 25 | Dendroside F ** | [M+Na]+ | C21H34O9 | 453.20569 | 2.746 | 124,435 | [28] |
| 26 | Gamma-Linolenic acid * | [M−H]− | C18H30O2 | 277.2161 | 10.686 | 32,712 | |
| 27 | Dendronobilin F ** | [M−H]− | C15H22O5 | 281.1403 | 8.168 | 7220 | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Liu, Z.; Hu, H.; Cai, R.; Bi, J.; Wang, Q.; Zhou, X.; Luo, H.; Zhang, C.; Wan, R. Dendrobium Nobile Alcohol Extract Extends the Lifespan of Caenorhabditis elegans via hsf-1 and daf-16. Molecules 2024, 29, 908. https://doi.org/10.3390/molecules29040908
Li L, Liu Z, Hu H, Cai R, Bi J, Wang Q, Zhou X, Luo H, Zhang C, Wan R. Dendrobium Nobile Alcohol Extract Extends the Lifespan of Caenorhabditis elegans via hsf-1 and daf-16. Molecules. 2024; 29(4):908. https://doi.org/10.3390/molecules29040908
Chicago/Turabian StyleLi, Linfeng, Zhen Liu, Huiling Hu, Renming Cai, Jingdou Bi, Qin Wang, Xiaogang Zhou, Huairong Luo, Chun Zhang, and Runlan Wan. 2024. "Dendrobium Nobile Alcohol Extract Extends the Lifespan of Caenorhabditis elegans via hsf-1 and daf-16" Molecules 29, no. 4: 908. https://doi.org/10.3390/molecules29040908
APA StyleLi, L., Liu, Z., Hu, H., Cai, R., Bi, J., Wang, Q., Zhou, X., Luo, H., Zhang, C., & Wan, R. (2024). Dendrobium Nobile Alcohol Extract Extends the Lifespan of Caenorhabditis elegans via hsf-1 and daf-16. Molecules, 29(4), 908. https://doi.org/10.3390/molecules29040908