

Comprehensive Analysis of 11 Species of Euodia (Rutaceae) by Untargeted LC-IT-TOF/MS Metabolomics and In Vitro Functional Methods
 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. LC-IT-TOF/MS Profiles of 11 Euodia Species
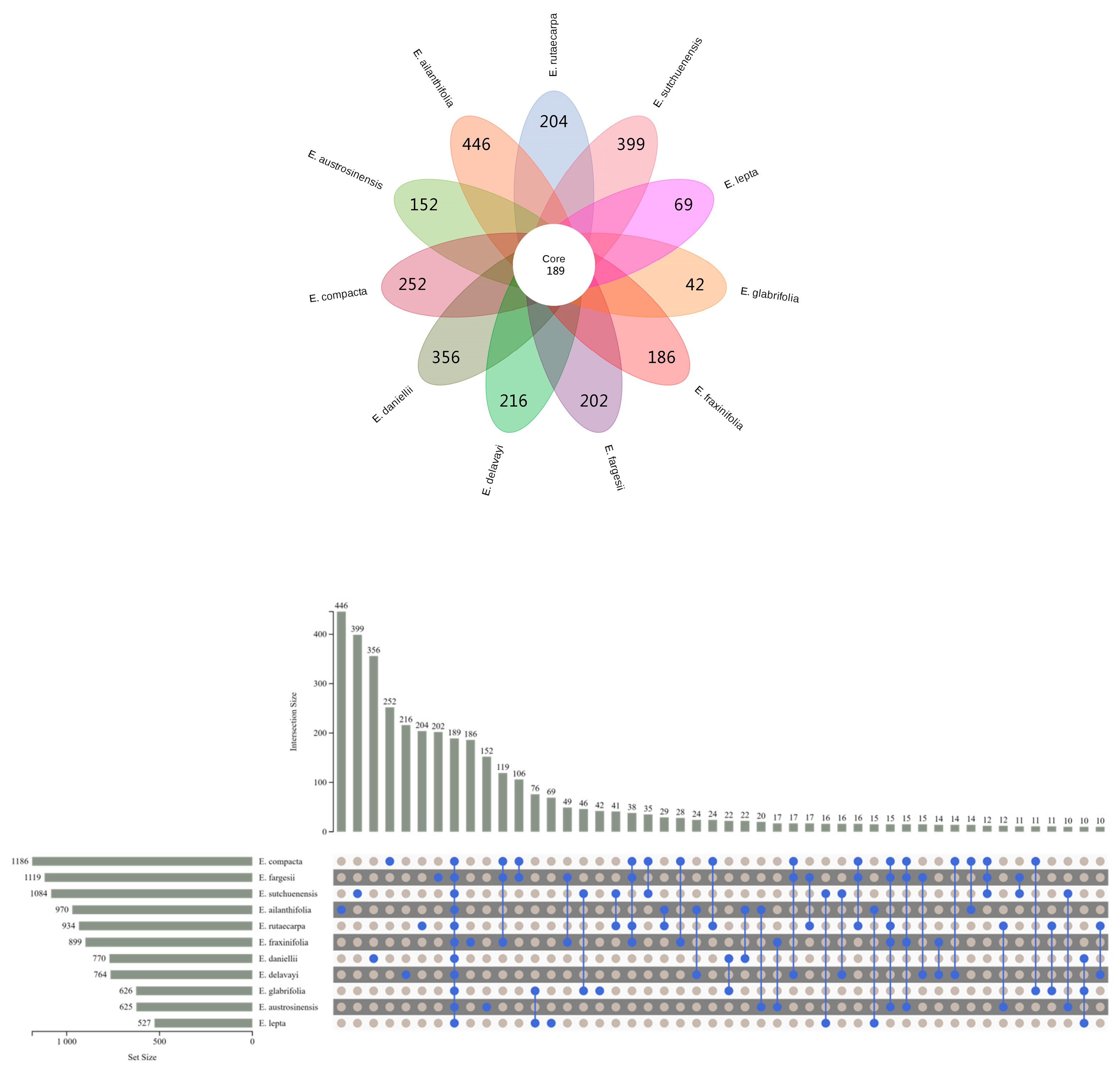
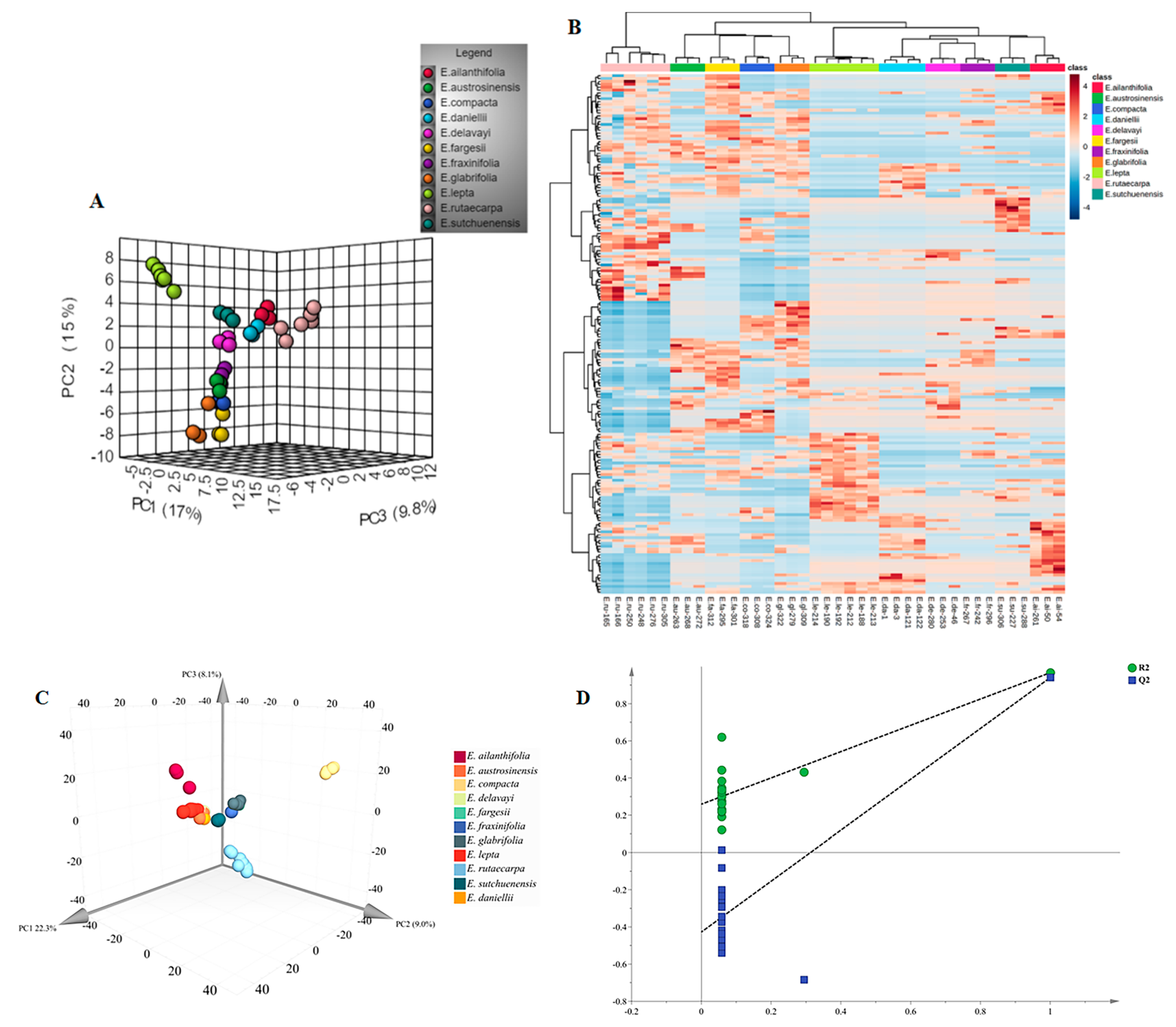
2.2. Metabolome Analysis of Euodia Species
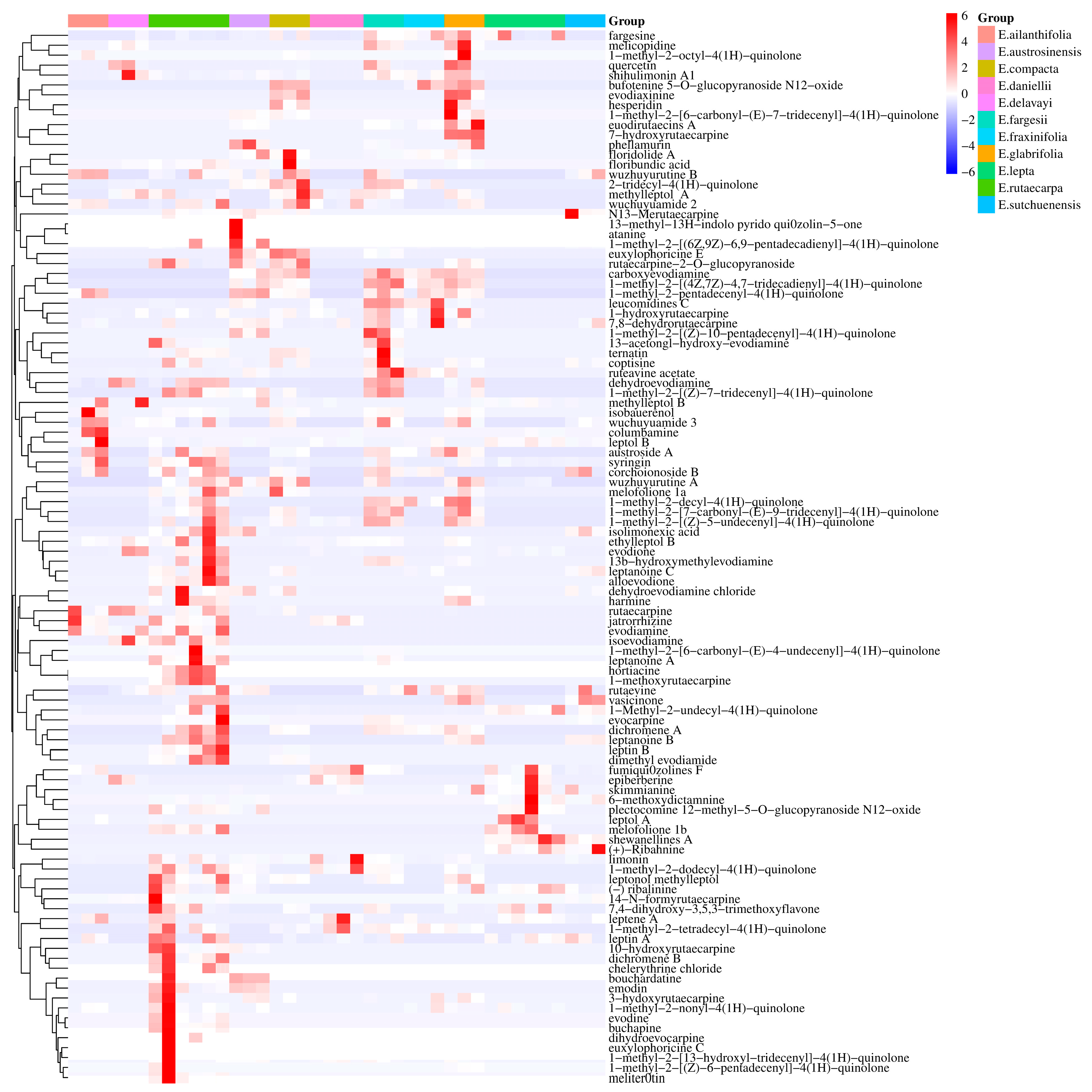
2.3. Differentially Accumulated Metabolites between Euodia Species
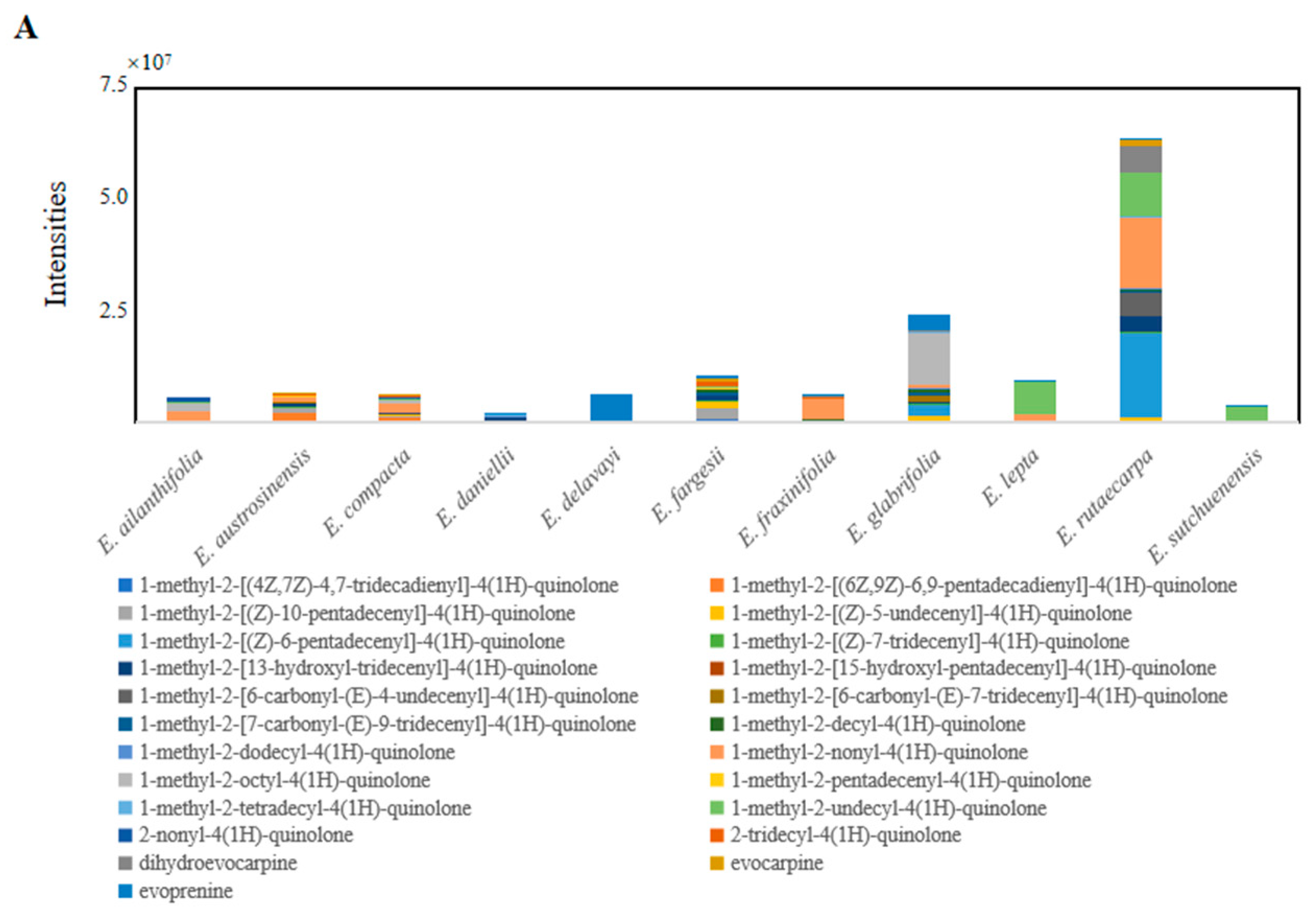
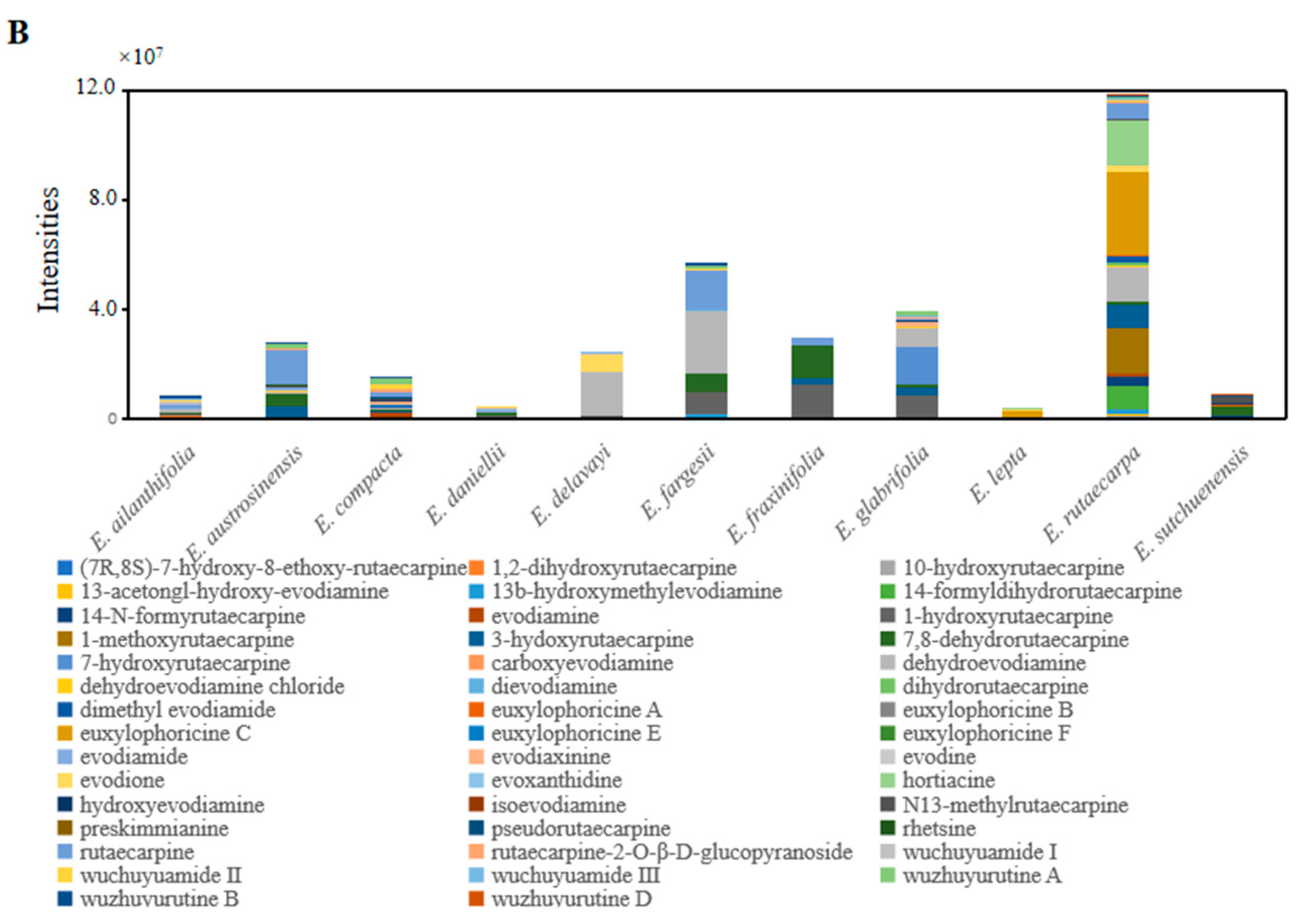
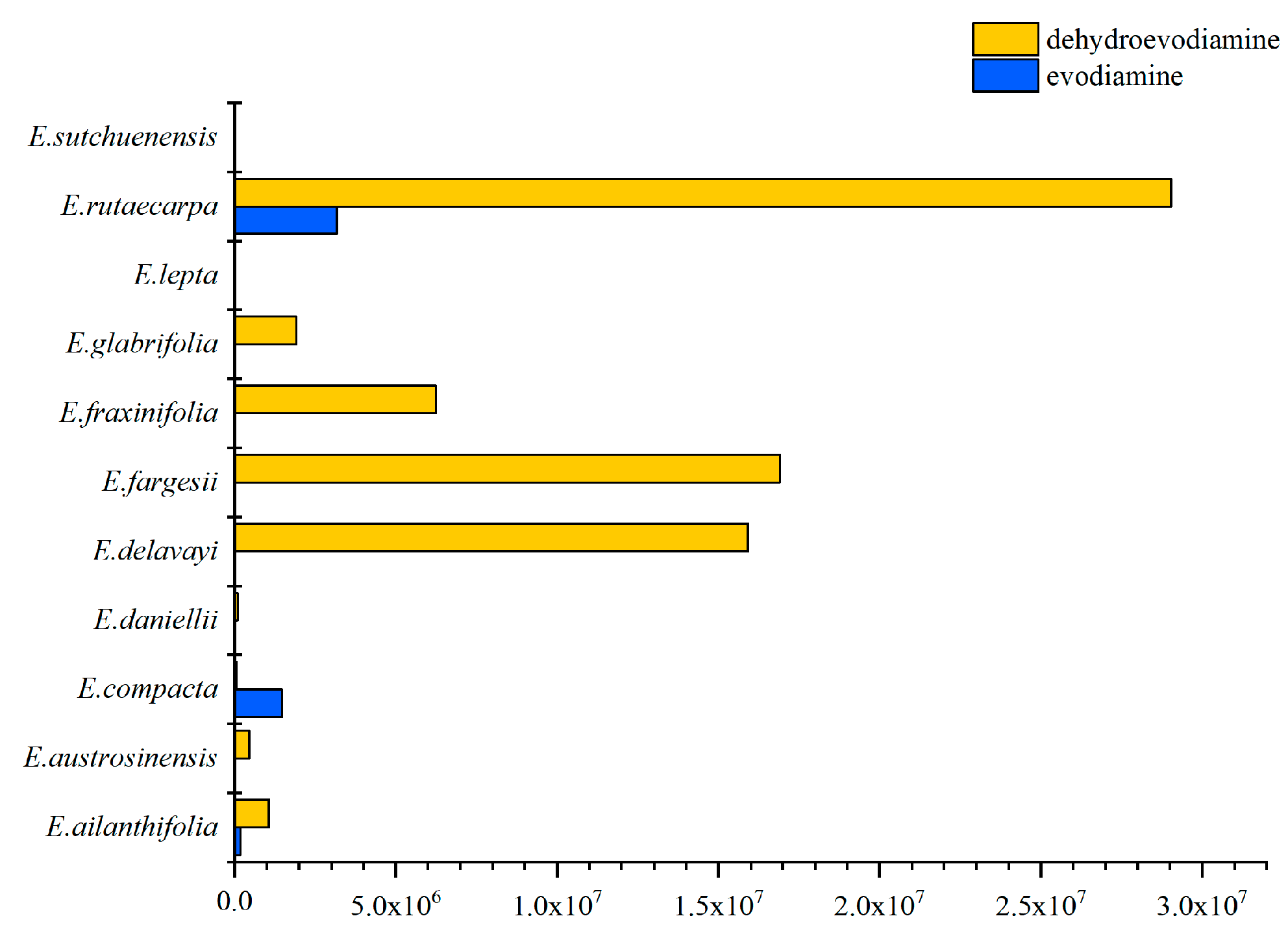
2.4. Specific Characteristics of Metabolites in Euodia Species
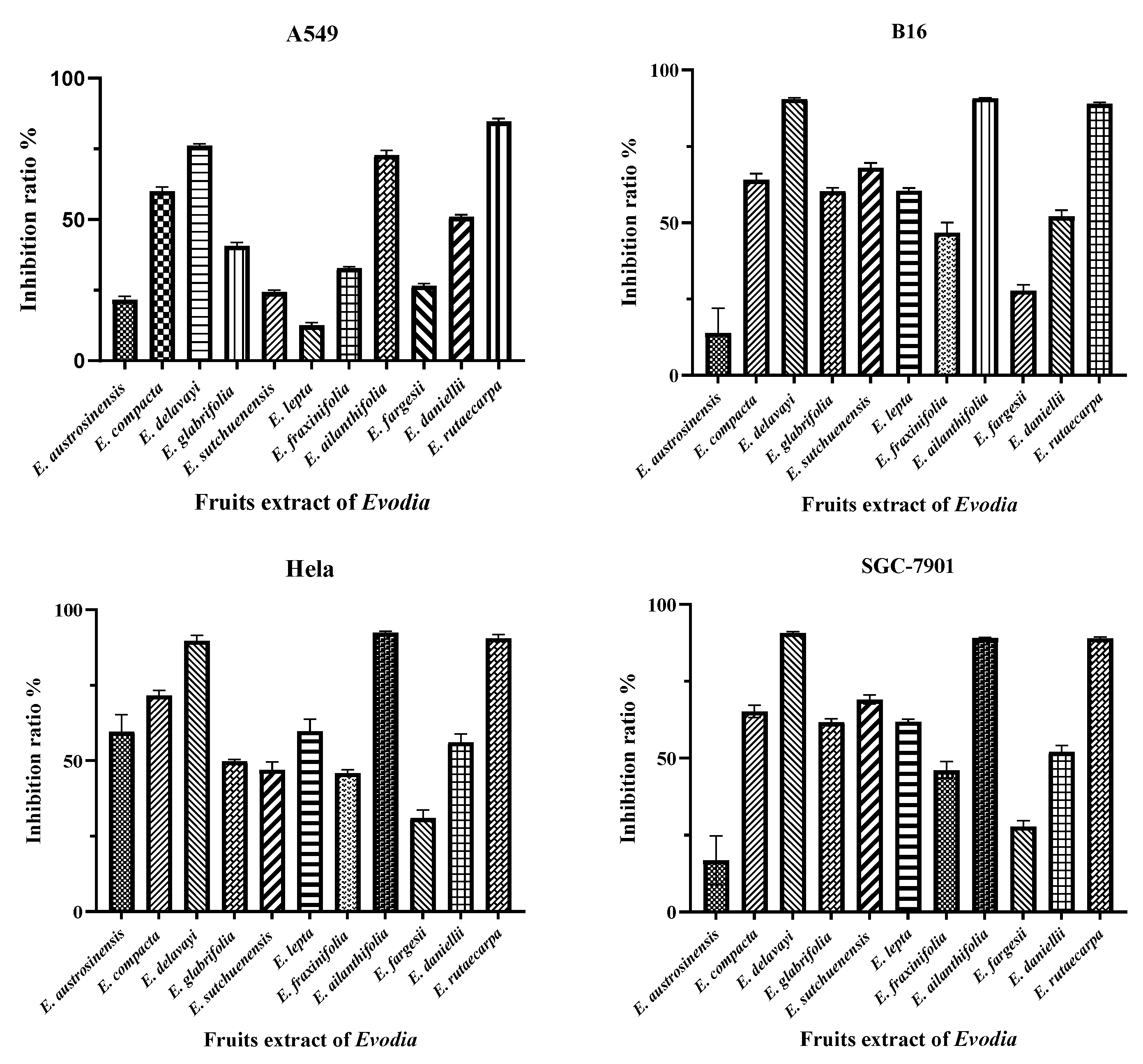
2.5. Biological Activity Determination
2.5.1. Anti-Tumor Activity
2.5.2. Antibacterial Activity
3. Materials and Methods
3.1. Solvents and Reagents
3.2. Plant Material
3.3. Sample Preparation
3.4. LC-IT-TOF/MS Apparatus and Conditions
3.5. Method Validation
3.6. Statistical Analysis of Metabolite Data
3.7. Determination of Biological Activities of Euodia Extracts
3.7.1. Cytotoxic Activity
3.7.2. Activity against Plant Pathogenic Fungi
3.7.3. Activity against Plant Pathogenic Bacteria
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Editorial Committee of Flora of China, Chinese Academy of Sciences. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1997. [Google Scholar]
- Liu, L.; Sun, X.; Guo, Y.; Ge, K. Evodiamine induces ROS-Dependent cytotoxicity in human gastric cancer cells via TRPV1/Ca2+ pathway. Chem. Biol. Interact. 2022, 351, 109756. [Google Scholar] [CrossRef]
- Byun, W.S.; Bae, E.S.; Kim, W.K.; Lee, S.K. Antitumor activity of rutaecarpine in human colorectal cancer cells by suppression of wnt/?-Catenin signaling. J. Nat. Prod. 2022, 85, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Wang, X.; Jin, L.; Yang, L.; Zhu, J.; Wang, H.; Zheng, F.; Cui, H.; Li, X.; et al. Evodiamine suppresses the progression of non-small cell lung carcinoma via endoplasmic reticulum stress-mediated apoptosis pathway in vivo and in vitro. Int. J. Immunopathol. Pharmacol. 2022, 36, 3946320221086079. [Google Scholar] [CrossRef] [PubMed]
- Panda, M.; Biswal, B.K. Evodiamine inhibits sternness and metastasis by altering the SOX9-β-catenin axis in non-small-cell lung cancer. J. Cell. Biochem. 2022, 123, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Kim, J.; Lee, P.; Kim, S. Evodiamine Inhibits Helicobacter pylori Growth and Helicobacter pylori-Induced Inflammation. Int. J. Mol. Sci. 2021, 22, 3385. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, H.; Zhang, X.; Peng, Y.; Deng, J.; Wang, Y.; Li, R.; Liu, L.; Wang, Z. Discovery of evodiamine derivatives as potential lead antifungal agents for the treatment of superficial fungal infections. Bioorg. Chem. 2022, 127, 105981. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, P.; Yang, J.; Yong, X.; Yin, M.; Chen, Y.; Feng, X.; Wang, Q. Inhibition efficacy of Tetradium glabrifolium fruit essential oil against Phytophthora capsici and potential mechanism. Ind. Crop. Prod. 2022, 176, 114310. [Google Scholar] [CrossRef]
- Jayakumar, T.; Yang, C.; Yen, T.; Hsu, C.; Sheu, J.; Hsia, C.; Manubolu, M.; Huang, W.; Hsieh, C.; Hsia, C. Anti-Inflammatory Mechanism of an Alkaloid Rutaecarpine in LTA-Stimulated RAW 264.7 Cells: Pivotal Role on NF-κB and ERK/p38 Signaling Molecules. Int. J. Mol. Sci. 2022, 23, 5889. [Google Scholar] [CrossRef]
- Lee, C.; Liao, J.; Chen, S.; Yen, C.; Lee, Y.; Huang, S.; Huang, S.; Lin, C.; Chang, V.H. Fluorine-Modified rutaecarpine exerts cyclooxygenase-2 inhibition and anti-inflammatory effects in lungs. Front. Pharmacol. 2019, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Guo, S.; Wang, Y.; Pang, X.; Geng, Z.; Du, S. Toxicity and repellency of essential oil from Evodia lenticellata Huang fruits and its major monoterpenes against three stored-product insects. Ecotox. Environ. Saf. 2018, 160, 342–348. [Google Scholar] [CrossRef]
- Liu, J.; Shi, Y.; Tian, Z.; Li, F.; Hao, Z.; Wen, W.; Zhang, L.; Wang, Y.; Li, Y.; Fan, Z. Bioactivity-Guided synthesis accelerates the discovery of evodiamine derivatives as potent insecticide candidates. J. Agric. Food Chem. 2022, 70, 5197–5206. [Google Scholar] [CrossRef]
- Fang, Z.; Tang, Y.; Ying, J.; Tang, C.; Wang, Q. Traditional Chinese medicine for anti-Alzheimer’s disease: Berberine and evodiamine from Evodia rutaecarpa. Chin. Med. 2020, 15, 82. [Google Scholar] [CrossRef] [PubMed]
- Bak, E.J.; Park, H.G.; Kim, J.M.; Kim, J.M.; Yoo, Y.; Cha, J. Inhibitory effect of evodiamine alone and in combination with rosiglitazone on in vitro adipocyte differentiation and in vivo obesity related to diabetes. Int. J. Obesity 2010, 34, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Li, J.; Xu, S. Rutaecarpine: A promising cardiovascular protective alkaloid from Evodia rutaecarpa (Wu Zhu Yu). Pharmacol. Res. 2019, 141, 541–550. [Google Scholar] [CrossRef]
- Kim, D.; Lee, Y.; Park, S.H.; Lee, M.J.; Kim, M.J.; Jang, H.; Lee, J.; Lee, H.; Han, B.S.; Son, W.; et al. Subchronic oral toxicity of evodia fruit powder in rats. J. Ethnopharmacol. 2014, 151, 1072–1078. [Google Scholar] [CrossRef]
- Yamashita, H.; Kusudo, T.; Takeuchi, T.; Qiao, S.; Tsutsumiuchi, K.; Wang, T.; Wang, Y. Dietary supplementation with evodiamine prevents obesity and improves insulin resistance in ageing mice. J. Funct. Foods 2015, 19, 320–329. [Google Scholar] [CrossRef]
- Tian, H.; Lam, S.M.; Shui, G. Metabolomics, a powerful tool for agricultural research. Int. J. Mol. Sci. 2016, 17, 1871. [Google Scholar] [CrossRef]
- Wang, P.; Wang, Q.; Yang, B.; Zhao, S.; Kuang, H. The progress of metabolomics study in traditional Chinese medicine research. Am. J. Chin. Med. 2015, 43, 1281–1310. [Google Scholar] [CrossRef]
- Tsugawa, H.; Rai, A.; Saito, K.; Nakabayashi, R. Metabolomics and complementary techniques to investigate the plant phytochemical cosmos. Nat. Prod. Rep. 2021, 38, 1729–1759. [Google Scholar] [CrossRef]
- Wen, J.; Tong, Y.; Ma, X.; Wang, R.; Li, R.; Song, H.; Zhao, Y. Therapeutic effects and potential mechanism of dehydroevodiamine on N-methyl-N′-nitro-N-nitrosoguanidine-induced chronic atrophic gastritis. Phytomedicine 2021, 91, 153619. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Chen, L.; He, Y.; He, T.; Wang, R.; Wei, S.; Jing, M.; Zhou, X.; Li, H.; et al. Integrated network pharmacology and metabolomics to investigate the effects and possible mechanisms of Dehydroevodiamine against ethanol-induced gastric ulcers. J. Ethnopharmacol. 2024, 319, 117340. [Google Scholar] [CrossRef]
- Luo, J.; Zou, W.; Li, J.; Liu, W.; Huang, J.; Wu, H.; Shen, J.; Li, F.; Yuan, J.; Tao, A.; et al. Untargeted serum and liver metabolomics analyses reveal the gastroprotective effect of polysaccharide from Evodiae fructus on ethanol-induced gastric ulcer in mice. Int. J. Biol. Macromol. 2023, 232, 123481. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lu, Q.; Chen, F.; Wang, S.; Niu, C.; Liao, J.; Wang, H.; Chen, F. Serum untargeted metabolomics analysis of the mechanisms of evodiamine on type 2 diabetes mellitus model rats. Food Funct. 2022, 13, 6623–6635. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, Y.; He, G.; Cao, Y.; Bi, M.; Song, M.; Yang, P.; Xu, L.; Ming, J. Comprehensive analysis of secondary metabolites in the extracts from different lily bulbs and their antioxidant ability. Antioxidants 2021, 10, 1634. [Google Scholar] [CrossRef]
- Jiang, N.; Zhu, H.; Liu, W.; Fan, C.; Jin, F.; Xiang, X. Metabolite Differences of Polyphenols in Different Litchi Cultivars (Litchi chinensis Sonn.) Based on Extensive Targeted Metabonomics. Molecules 2021, 26, 1181. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Deng, J.; Lin, X.; Li, Y.; Lin, Y.; Xia, B.; Lin, L. Metabolomics Reveals Distinct Metabolites between Lonicera japonica and Lonicera macranthoides Based on GC-MS. J. Chem. 2020, 2020, 6738571. [Google Scholar] [CrossRef]
- Pereira, A.G.; Cassani, L.; Oludemi, T.; Chamorro, F.; Calhelha, R.C.; Prieto, M.A.; Barros, L.; Simal-Gandara, J.; Lucini, L.; Garcia-Perez, P. Untargeted metabolomics and in vitro functional analysis unravel the intraspecific bioactive potential of flowers from underexplored Camellia japonica cultivars facing their industrial application. Ind. Crop. Prod. 2023, 204, 117389. [Google Scholar] [CrossRef]
- Chen, X.; Xu, Y.; Meng, L.; Chen, X.; Yuan, L.; Cai, Q.; Shi, W.; Huang, G. Non-parametric partial least squares-discriminant analysis model based on sum of ranking difference algorithm for tea grade identification using electronic tongue data. Sens. Actuat. B Chem. 2020, 311, 127924. [Google Scholar] [CrossRef]
- Truzzi, E.; Marchetti, L.; Fratagnoli, A.; Rossi, M.C.; Bertelli, D. Novel application of 1H NMR spectroscopy coupled with chemometrics for the authentication of dark chocolate. Food Chem. 2023, 404, 134522. [Google Scholar] [CrossRef]
- Yan, Q.; Yong, X.; Yin, M.; Shan, Y.; Xu, S.; Su, X.; Feng, X.; Wang, Q. Phytochemical and chemotaxonomic study on Evodia rutaecarpa and Evodia rutaecarpa var. Bodinieri. Biochem. Syst. Ecol. 2022, 104, 104472. [Google Scholar] [CrossRef]
- Lin, L.; Ren, L.; Wen, L.; Wang, Y.; Qi, J. Effect of evodiamine on the proliferation and apoptosis of A549 human lung cancer cells. Mol. Med. Rep. 2016, 14, 2832–2838. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Park, S.H.; Min, H.; Park, H.J.; Lee, S.K. Anti-proliferative effects of evodiamine in human lung cancer cells. J. Cancer Prev. 2014, 19, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Zeng, C.; Qu, J.; Yan, X.; Lin, Z. Rutaecarpine increases anticancer drug sensitivity in Drug-Resistant cells through MARCH8-Dependent ABCB1 degradation. Biomedicines 2021, 9, 1143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, L.J.; Tashiro, S.; Onodera, S.; Ikejima, T. Evodiamine induces tumor cell death through different pathways: Apoptosis and necrosis. Acta Pharmacol. Sin. 2004, 25, 83–89. [Google Scholar] [PubMed]
- Fei, X.F.; Wang, B.X.; Li, T.J.; Tashiro, S.; Minami, M.; Xing, D.J.; Ikejima, T. Evodiamine, a constituent of Evodiae Fructus, induces anti-proliferating effects in tumor cells. Cancer Sci. 2003, 94, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lin, L.; Choi, Y.; Michniak-Kohn, B. Development and in-vitro evaluation of co-loaded berberine chloride and evodiamine ethosomes for treatment of melanoma. Int. J. Pharmaceut. 2020, 581, 119278. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, M.; Matsunaga, T.; Takahashi, S.; Saiki, I.; Suzuki, H. Anti-invasive and metastatic activities of evodiamine. Biol. Pharm. Bull. 2002, 25, 1491–1493. [Google Scholar] [CrossRef]
- Yang, L.; Liu, X.; Wu, D.; Zhang, M.; Ran, G.; Bi, Y.; Huang, H. Growth inhibition and induction of apoptosis in SGC-7901 human gastric cancer cells by evodiamine. Mol. Med. Rep. 2014, 9, 1147–1152. [Google Scholar] [CrossRef]
- Rasul, A.; Yu, B.; Zhong, L.; Khan, M.; Yang, H.; Ma, T. Cytotoxic effect of evodiamine in SGC-7901 human gastric adenocarcinoma cells via simultaneous induction of apoptosis and autophagy. Oncol. Rep. 2012, 27, 1481–1487. [Google Scholar]
- Deng, J.; Long, L.; Peng, X.; Jiang, W.; Peng, Y.; Zhang, X.; Zhao, Y.; Tian, Y.; Wang, Z.; Zhuo, L. N(14)-substituted evodiamine derivatives as dual topoisomerase 1/tubulin-Inhibiting anti-gastrointestinal tumor agents. Eur. J. Med. Chem. 2023, 255, 115366. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Z.; Tan, H. Novel zinc(II)-curcumin molecular probes bearing berberine and jatrorrhizine derivatives as potential mitochondria-targeting anti-neoplastic drugs. Eur. J. Med. Chem. 2022, 243, 114736. [Google Scholar] [CrossRef]
- Qin, Q.; Zou, B.; Wang, Z.; Huang, X.; Zhang, Y.; Tan, M.; Wang, S.; Liang, H. High in vitro and in vivo antitumor activities of luminecent platinum(II) complexes with jatrorrhizine derivatives. Eur. J. Med. Chem. 2019, 183, 111727. [Google Scholar] [CrossRef]
- Chen, Y.; Li, E.; Xu, L. Guide to metabolomics analysis: A bioinformatics workflow. Metabolites 2022, 12, 357. [Google Scholar] [CrossRef]
- Allevato, D.M.; Kiyota, E.; Mazzafera, P.; Nixon, K.C. Ecometabolomic Analysis of Wild Populations of Pilocarpus pennatifolius (Rutaceae) Using Unimodal Analyses. Front. Plant Sci. 2019, 10, 258. [Google Scholar] [CrossRef]
- Wang, J.; Hou, J.; Huang, C.; Wang, W.; Liu, Y.; Zhang, H.; Yan, D.; Zeng, K.; Yao, S. Activation of the phenylpropanoid pathway in Citrus sinensis collapsed vesicles during segment drying revealed by physicochemical and targeted metabolomics analysis. Food Chem. 2023, 409, 135297. [Google Scholar] [CrossRef]
- Cao, Y.; Ren, M.; Yang, J.; Guo, L.; Lin, Y.; Wu, H.; Wang, B.; Lv, R.; Zhang, C.; Gong, X.; et al. Comparative metabolomics analysis of pericarp from four varieties of Zanthoxylum bungeanum Maxim. Bioengineered 2022, 13, 14815–14826. [Google Scholar] [CrossRef]
- Shang, X.; Morris-Natschke, S.L.; Yang, G.; Liu, Y.; Guo, X.; Xu, X.; Goto, M.; Li, J.; Zhang, J.; Lee, K. Biologically active quinoline and quinazoline alkaloids part II. Med. Res. Rev. 2018, 38, 1614–1660. [Google Scholar] [CrossRef]
- Yang, X.W.; Zhang, H.; Li, M.; Du, L.J.; Yang, Z.; Xiao, S.Y. Studies on the alkaloid constituents of Evodia rutaecarpa (Juss) Benth var. Bodinaieri (Dode) Huang and their acute toxicity in mice. J. Asian Nat. Prod. Res. 2006, 8, 697–703. [Google Scholar]
- An, C.; Li, X.; Luo, H.; Li, C.; Wang, M.; Xu, G.; Wang, B. 4-Phenyl-3,4-dihydroquinolone Derivatives from Aspergillus nidulans MA-143, an Endophytic Fungus Isolated from the Mangrove Plant Rhizophora stylosa. J. Nat. Prod. 2013, 76, 1896–1901. [Google Scholar] [CrossRef]
- Zhao, N.; Li, Z.; Li, D.; Sun, Y.; Shan, D.; Bai, J.; Pei, Y.; Jing, Y.; Hua, H. Quinolone and indole alkaloids from the fruits of Euodia rutaecarpa and their cytotoxicity against two human cancer cell lines. Phytochemistry 2015, 109, 133–139. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0-making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.; Yao, J.; Wang, Q.; Li, L.; Wang, B.; Feng, X.; Wang, Z.; Yin, M.; Chen, Y.; Xu, S. Antifungal effects of amaryllidaceous alkaloids from bulbs of Lycoris spp. Against Magnaporthe oryzae. Pest Manag. Sci. 2023, 79, 2423–2432. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Xu, S.; Shan, Y.; Yin, M.; Chen, Y.; Feng, X.; Wang, Q. Three new quinazolines from Evodia rutaecarpa and their biological activity. Fitoterapia 2018, 127, 186–192. [Google Scholar] [CrossRef]
- Yan, Q.; Shan, Y.; Yin, M.; Xu, S.; Ma, C.; Tong, H.; Feng, X.; Wang, Q. Phytochemical and chemotaxonomic study on Evodia rutaecarpa var. officinalis. Biochem. Syst. Ecol. 2020, 88, 103961. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytotoxic Activity (IC50, µg/mL) | ||||
|---|---|---|---|---|
| Species | A549 | B16 | Hela | SGC-7901 |
| E. delavayi | 67.23 ± 1.52 | 50.37 ± 0.97 | 52.20 ± 1.12 | 51.87 ± 0.47 |
| E. ailanthifolia | 69.07 ± 2.31 | 51.23 ± 0.45 | 50.07 ± 0.53 | 53.25 ± 0.38 |
| E. rutaecarpa | 62.16 ± 1.44 | 53.15 ± 0.64 | 51.01 ± 1.35 | 51.21 ± 0.35 |
| cisplatin a | 3.34 ± 0.12 | 2.44 ± 1.22 | 4.22 ± 1.92 | 3.12 ± 0.12 |
| Compounds | Sclerotinia sclerotiorum (10 μg/mL, %) | Rhizoctonia solani (10 μg/mL, %) | Fusarium graminearum (10 μg/mL, %) | Botrytis cinerea (10 μg/mL, %) | Colletotrichum gloeosporioides (10 μg/mL, %) | Xanthomonas oryzae pv. Oryzae (10 μg/mL, %) |
|---|---|---|---|---|---|---|
| rutaecarpine | 8.20 ± 1.21 | 11.11 ± 1.11 | 17.78 ± 1.67 | 8.89 ± 4.44 | 13.33 ± 2.22 | 4.88 ± 2.51 |
| evodiamine | 40.00 ± 5.56 | 33.33 ± 2.78 | 22.22 ± 2.78 | 22.22 ± 3.89 | 37.77 ± 3.89 | 13.13 ± 1.24 |
| dihydroevocarpine | 0.56 ± 2.32 | 5.56 ± 1.11 | 4.24 ± 2.14 | 8.75 ± 4.44 | 3.70 ± 1.11 | 8.0 ± 2.11 |
| dehydroevodiamine | 6.67 ± 1.11 | 5.12 ± 4.12 | 13.33 ± 1.67 | 20.00 ± 3.89 | 20.00 ± 3.33 | 84.20 ± 6.87 |
| evodiaxinxine | 15.71 ± 1.21 | 8.91 ± 2.22 | 9.97 ± 1.67 | 2.70 ± 1.24 | 11.07 ± 3.33 | 1.16 ± 2.22 |
| wuzhuyurutine B | 10.12 ± 1.11 | 0.66 ± 0.54 | 4.79 ± 0.67 | 7.27 ± 3.33 | 4.20 ± 1.11 | 4.2 ± 1.11 |
| wuchuyuamide III | 10.90 ± 2.22 | 12.54 ± 1.67 | 17.87 ± 2.14 | 6.08 ± 2.22 | 18.70 ± 2.22 | 0.83 ± 1.24 |
| No. | Species | Sample Symbol | Collection Year | GPS Coordinates of Collection Site |
|---|---|---|---|---|
| 1 | Euodia lepta (E. lepta) | ELE20171008-A | 8 October 2017 | 21°55′27.26″ N 101°15′27.44″ E |
| 2 | Euodia lepta (E. lepta) | ELE20171011-B | 11 October 2017 | 22°50′7.36″ N 100°59′56.94″ E |
| 3 | Euodia lepta (E. lepta) | ELE20171017-C | 17 October 2017 | 24°53′49.81″ N 110°57′51.90″ E |
| 4 | Euodia lepta (E. lepta) | ELE20160801-D | 1 August 2016 | 18°42′17.72″ N 109°51′17.81″ E |
| 5 | Euodia lepta (E. lepta) | ELE20160801-E | 1 August 2016 | 18°39′20.93″ N 109°54′29.38″ E |
| 6 | Euodia lepta (E. lepta) | ELE20160801-F | 1 August 2016 | 18°40′28.84″ N 109°54′10.84″ E |
| 7 | Euodia rutaecarpa (E. rutaecarpa) | ERU20181024-A | 24 September 2018 | 32°03′09.68″ N 118°50′03.20″ E |
| 8 | Euodia rutaecarpa (E. rutaecarpa) | ERU20160715-B | 15 September 2016 | 26°16′12.57″ N 106°58′49.82″ E |
| 9 | Euodia rutaecarpa (E. rutaecarpa) | ERU20160727-C | 27 August 2016 | 27°36′20.64″ N 105°50′46.37″ E |
| 10 | Euodia rutaecarpa (E. rutaecarpa) | ERU20160731-D | 31 August 2016 | 25°54′0.55″ N 104°59′43.88″ E |
| 11 | Euodia rutaecarpa (E. rutaecarpa) | ERU20180721-E | 21 September 2018 | 27°50′46.12″ N 109°14′29.63″ E |
| 12 | Euodia rutaecarpa (E. rutaecarpa) | ERU20180722-F | 22 September 2018 | 27°13′16.53″ N 107°56′22.79″ E |
| 13 | Euodia compacta (E. compacta) | ECO20180521-A | 21 August 2018 | 30°32′45.75″ N 114°25′18.06″ E |
| 14 | Euodia compacta (E. compacta) | ECO20180910-B | 10 September 2018 | 30°15′10.26″ N 120°07′05.53″ E |
| 15 | Euodia compacta (E. compacta) | ECO20180910-C | 10 September 2018 | 30°15′13.40″ N 120°07′02.75″ E |
| 16 | Euodia austrosinensis (E. austrosinensis) | EAU20181008-A | 8 October 2018 | 21°55′27.26″ N 101°15′27.44″ E |
| 17 | Euodia austrosinensis (E. austrosinensis) | EAU20181017-B | 17 October 2018 | 24°53′54.01″ N 110°59′19.65″ E |
| 18 | Euodia austrosinensis (E. austrosinensis) | EAU20180730-C | 30 August 2018 | 21°55′27.26″ N 101°15′27.44″ E |
| 19 | Euodia glabrifolia (E. glabrifolia) | EGL20160801-A | 1 August 2016 | 18°41′16.85″ N 109°43′44.21″ E |
| 20 | Euodia glabrifolia (E. glabrifolia) | EGL20160801-B | 1 August 2016 | 18°40′51.09″ N 109°51′35.11″ E |
| 21 | Euodia glabrifolia (E. glabrifolia) | EGL20160801-C | 1 August 2016 | 18°42′31.58″ N 109°50′28.71″ E |
| 22 | Euodia ailanthifolia (E. ailanthifolia) | EAI20181008-A | 8 October 2018 | 21°55′27.26″ N 101°15′27.44″ E |
| 23 | Euodia ailanthifolia (E. ailanthifolia) | EAI20181011-B | 22 October 2018 | 22°50′7.36″ N 100°59′56.94″ E |
| 24 | Euodia ailanthifolia (E. ailanthifolia) | EAI20180729-C | 29 August 2018 | 22°46′21.55″ N 100°59′31.66″ E |
| 25 | Euodia fargesii (E. fargesii) | EFA20160910-A | 10 September 2016 | 30°14′57.93″ N 120°07′06.80″ E |
| 26 | Euodia fargesii (E. fargesii) | EFA20160731-B | 31 August 2016 | 25°47′13.18″ N 104°58′17.48″ E |
| 27 | Euodia fargesii (E. fargesii) | EFA20180518-C | 18 September 2016 | 28°06′11.01″ N 113°01′55.86″ E |
| 28 | Euodia sutchuenensis (E. sutchuenensis) | ESU20160816-A | 16 August 2016 | 29°04′20.90″ N 107°09′38.44″ E |
| 29 | Euodia sutchuenensis (E. sutchuenensis) | ESU20160817-B | 17 August 2016 | 29°05′59.06″ N 107°10′06.24″ E |
| 30 | Euodia sutchuenensis (E. sutchuenensis) | ESU20180811-C | 11 August 2018 | 29°04′51.28″ N 107°08′49.46″ E |
| 31 | Euodia daniellii (E. daniellii) | EDA20170916-A | 16 September 2017 | 41°54′33.51″ N 123°36′01.37″ E |
| 32 | Euodia daniellii (E. daniellii) | EDA20170920-B | 20 September 2017 | 34°42′49.50″ N 119°22′41.94″ E |
| 33 | Euodia daniellii (E. daniellii) | EDA20170924-C | 24 September 2017 | 36°38′48.54″ N 117°01′18.37″ E |
| 34 | Euodia daniellii (E. daniellii) | EDA20170924-D | 24 September 2017 | 36°38′46.09″ N 117°01′21.31″ E |
| 35 | Euodia delavayi (E. delavayi) | EDE20181024-A | 24 October 2018 | 28°26′32.58″ N 98°54′49.57″ E |
| 36 | Euodia delavayi (E. delavayi) | EDE20180804-B | 4 August 2018 | 26°52′10.29″ N 100°13′54.36″ E |
| 37 | Euodia delavayi (E. delavayi) | EDE20180810-C | 10 August 2018 | 29°03′59.43″ N 107°08′43.13″ E |
| 38 | Euodia fraxinifolia (E. fraxinifolia) | EFR20180808-A | 8 August 2018 | 24°45′51.16″ N 100°30′10.87″ E |
| 39 | Euodia fraxinifolia (E. fraxinifolia) | EFR20180802-B | 2 August 2018 | 24°23′28.77″ N 100°46′47.38″ E |
| 40 | Euodia fraxinifolia (E. fraxinifolia) | EFR20180806-C | 6 August 2018 | 25°27′45.18″ N 98°45′54.14″ E |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yong, X.; Wang, B.; Wang, M.; Lyu, H.; Yin, M.; Jin, T.; Feng, X.; Shan, Y.; Liang, Y.; Wang, Q. Comprehensive Analysis of 11 Species of Euodia (Rutaceae) by Untargeted LC-IT-TOF/MS Metabolomics and In Vitro Functional Methods. Molecules 2024, 29, 1059. https://doi.org/10.3390/molecules29051059
Yong X, Wang B, Wang M, Lyu H, Yin M, Jin T, Feng X, Shan Y, Liang Y, Wang Q. Comprehensive Analysis of 11 Species of Euodia (Rutaceae) by Untargeted LC-IT-TOF/MS Metabolomics and In Vitro Functional Methods. Molecules. 2024; 29(5):1059. https://doi.org/10.3390/molecules29051059
Chicago/Turabian StyleYong, Xuhong, Bi Wang, Mengdi Wang, Hui Lyu, Min Yin, Tong Jin, Xu Feng, Yu Shan, Yan Liang, and Qizhi Wang. 2024. "Comprehensive Analysis of 11 Species of Euodia (Rutaceae) by Untargeted LC-IT-TOF/MS Metabolomics and In Vitro Functional Methods" Molecules 29, no. 5: 1059. https://doi.org/10.3390/molecules29051059
APA StyleYong, X., Wang, B., Wang, M., Lyu, H., Yin, M., Jin, T., Feng, X., Shan, Y., Liang, Y., & Wang, Q. (2024). Comprehensive Analysis of 11 Species of Euodia (Rutaceae) by Untargeted LC-IT-TOF/MS Metabolomics and In Vitro Functional Methods. Molecules, 29(5), 1059. https://doi.org/10.3390/molecules29051059