
Glycosylation Modulates the Structure and Functions of Collagen: A Review


Abstract
:1. Introduction
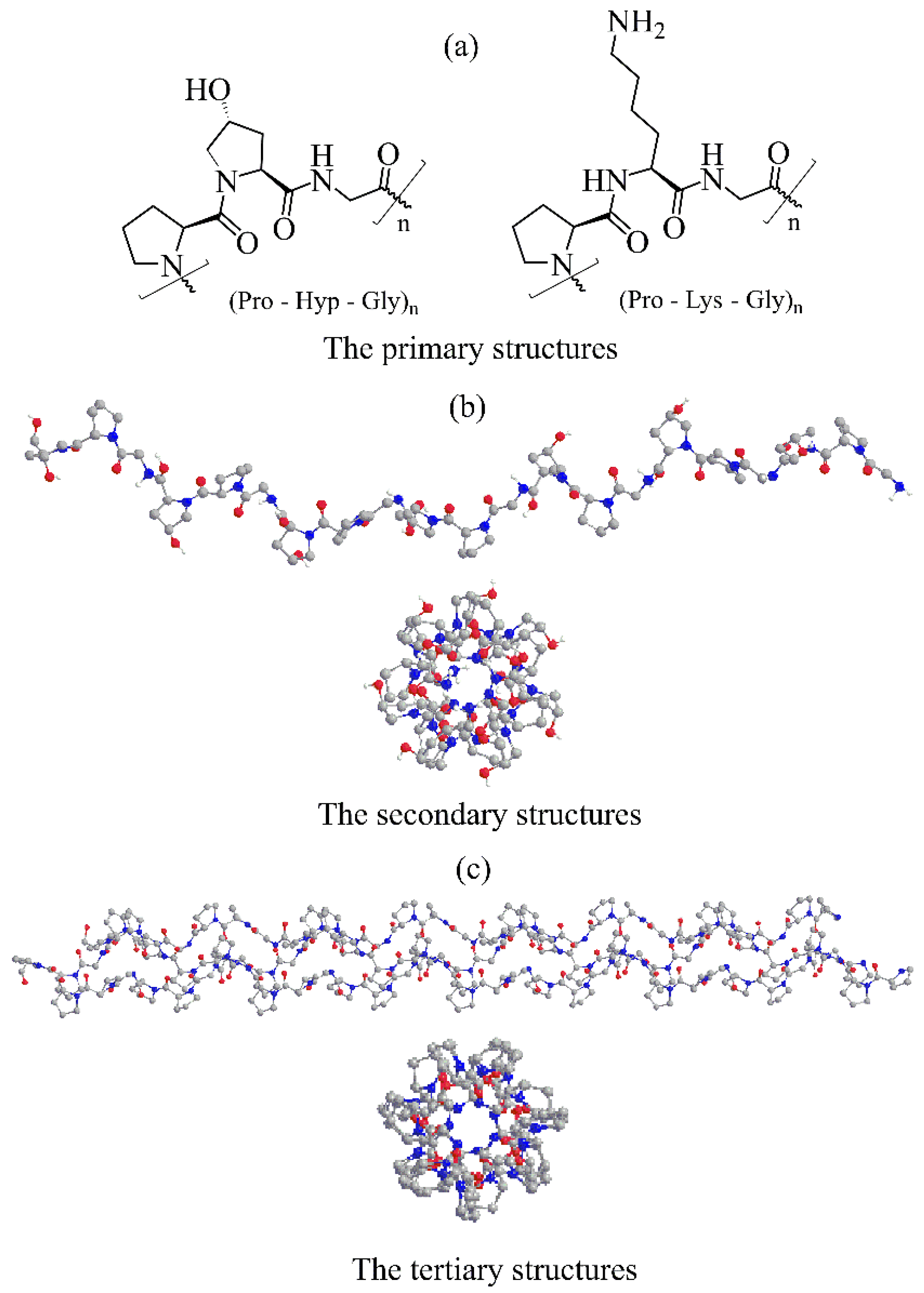
2. Collagen Structure
2.1. The Primary Structure
2.2. The Secondary Structure
2.3. The Tertiary Structure
2.4. The Quarternary Structure
2.5. Supramolecular Assemblies
3. Post-Translation Modifications of Collagen
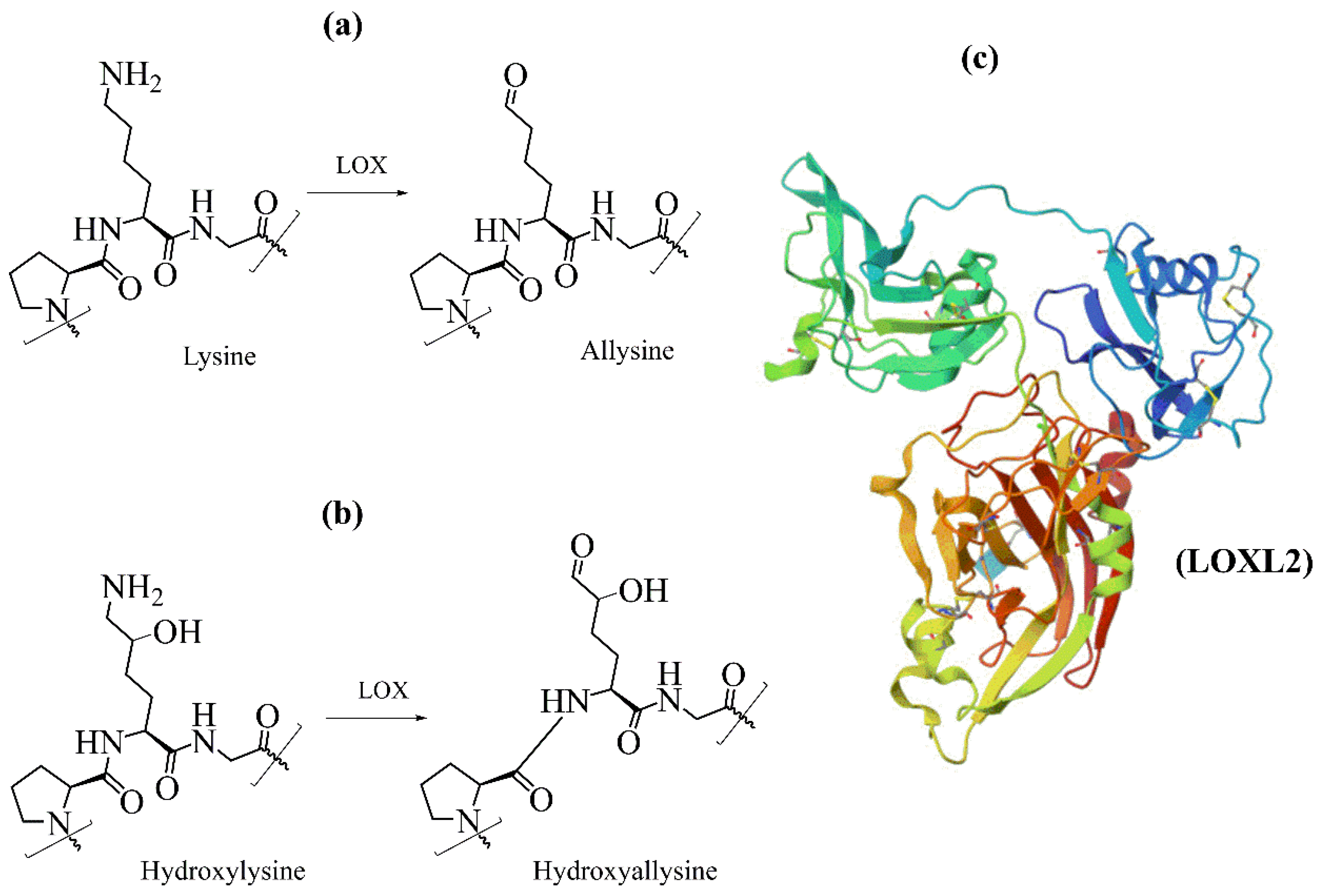
4. Hydroxylation of Proline and Lysine Residues
4.1. Prolyl 4-Hydroxylase
4.2. Prolyl 3-Hydroxylase
4.3. Lysyl Hydroxylases
5. Oxidation of Lysyl Residues
6. Glycosylation of Collagens
6.1. Glycation
6.2. Enzymatic Glycosylation
6.2.1. N-Glycan
6.2.2. O-Glycans
6.3. Procollagen Galactosyltransferases
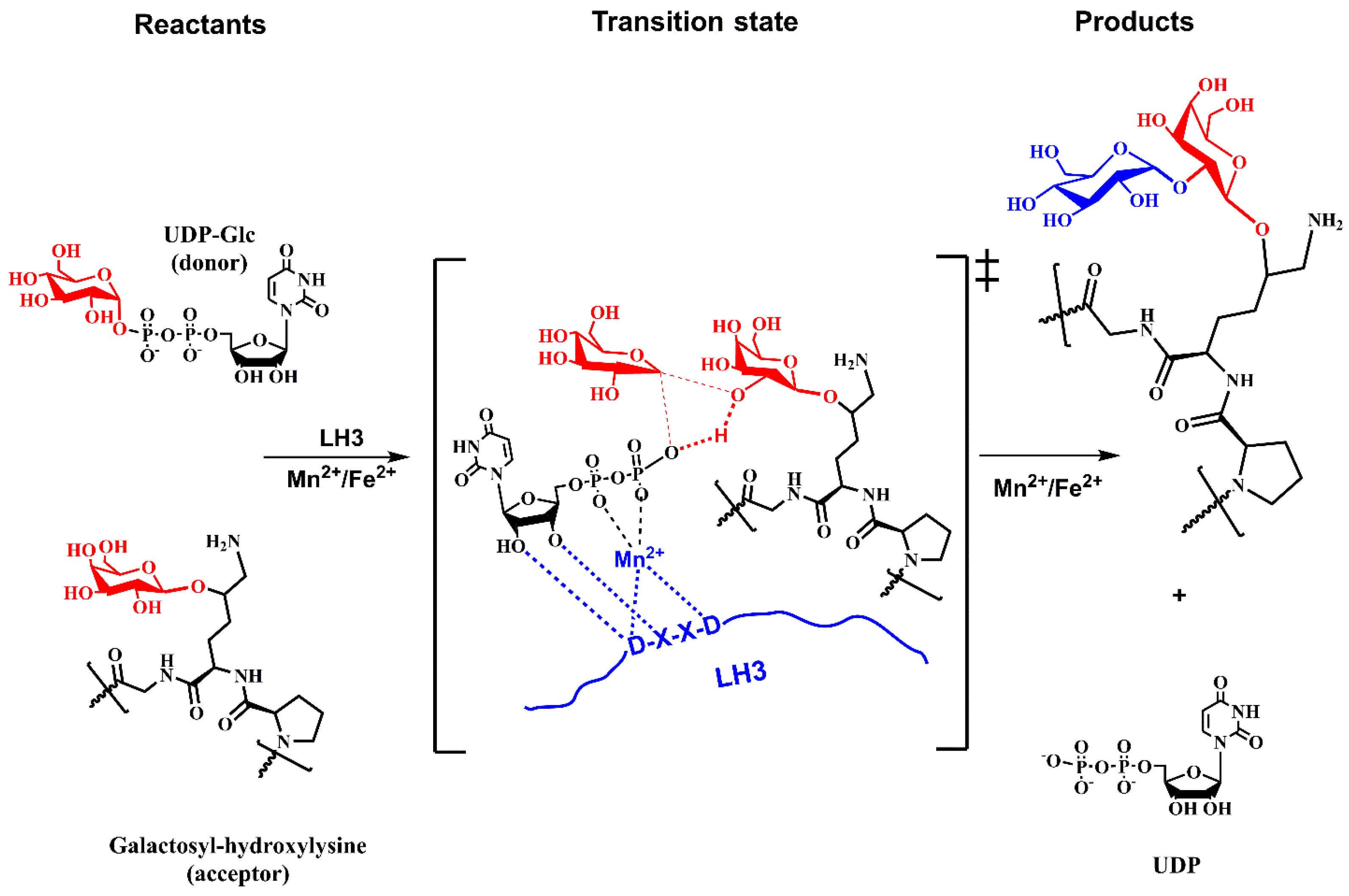
6.4. Lysyl Hydroxylases
6.5. Collagen Glucosidase
7. Functional Role of Collagen Glycosylation
7.1. N-Glycan Functions
7.2. O-Glycan Functions
8. Glycosylation Diseases
9. Concluding Remarks
Funding
Conflicts of Interest
References
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E.H.; et al. A guide to the composition and functions of the extracellular matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma 2014, 23, S20–S23. [Google Scholar] [CrossRef]
- Kular, J.K.; Basu, S.; Sharma, R.I. The extracellular matrix: Structure, composition, age-related differences, tools for analysis and applications for tissue engineering. J. Tissue Eng. 2014, 5, 2041731414557112. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Dandara, C. The Extracellular Matrix: Its Composition, Function, Remodeling, and Role in Tumorigenesis. Biomimetics 2023, 8, 146. [Google Scholar] [CrossRef]
- Van der Rest, M.; Garrone, R. Collagen Family of Proteins. FASEB J. 1991, 5, 2814. [Google Scholar] [CrossRef] [PubMed]
- Beachley, V.Z.; Wolf, M.T.; Sadtler, K.; Manda, S.S.; Jacobs, H.; Blatchley, M.R.; Bader, J.S.; Pandey, A.; Pardoll, D.; Elisseeff, J.H. Tissue matrix arrays for high-throughput screening and systems analysis of cell function. Nat. Methods 2015, 12, 1197–1204. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef]
- Mienaltowski, M.J.; Birk, D.E. Structure, physiology, and biochemistry of collagens. Adv. Exp. Med. Biol. 2014, 802, 5–29. [Google Scholar] [CrossRef]
- Gelse, K.; Poschl, E.; Aigner, T. Collagens--structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef]
- Wolff, D. Collagen Diseases. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2008; pp. 1–9. [Google Scholar] [CrossRef]
- Fidler, A.L.; Boudko, S.P.; Rokas, A.; Hudson, B.G. The triple helix of collagens—An ancient protein structure that enabled animal multicellularity and tissue evolution. J. Cell Sci. 2018, 131, jcs203950. [Google Scholar] [CrossRef]
- Arseni, L.; Lombardi, A.; Orioli, D. From Structure to Phenotype: Impact of Collagen Alterations on Human Health. Int. J. Mol. Sci. 2018, 19, 1407. [Google Scholar] [CrossRef] [PubMed]
- Romero-Ortuno, R.; Kenny, R.A.; McManus, R. Collagens and elastin genetic variations and their potential role in aging-related diseases and longevity in humans. Exp. Gerontol. 2020, 129, 110781. [Google Scholar] [CrossRef] [PubMed]
- Jobling, R.; D’Souza, R.; Baker, N.; Lara-Corrales, I.; Mendoza-Londono, R.; Dupuis, L.; Savarirayan, R.; Ala-Kokko, L.; Kannu, P. The collagenopathies: Review of clinical phenotypes and molecular correlations. Curr. Rheumatol. Rep. 2014, 16, 394. [Google Scholar] [CrossRef] [PubMed]
- Necula, L.; Matei, L.; Dragu, D.; Pitica, I.; Neagu, A.; Bleotu, C.; Diaconu, C.C.; Chivu-Economescu, M. Collagen Family as Promising Biomarkers and Therapeutic Targets in Cancer. Int. J. Mol. Sci. 2022, 23, 12415. [Google Scholar] [CrossRef] [PubMed]
- Romer, A.M.A.; Thorseth, M.L.; Madsen, D.H. Immune Modulatory Properties of Collagen in Cancer. Front. Immunol. 2021, 12, 791453. [Google Scholar] [CrossRef] [PubMed]
- Leikina, E.; Mertts, M.V.; Kuznetsova, N.; Leikin, S. Type I collagen is thermally unstable at body temperature. Proc. Natl. Acad. Sci. USA 2002, 99, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G. Characterization and quantitative determination of the hydroxylysine-linked carbohydrate units of several collagens. J. Biol. Chem. 1969, 244, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Hennet, T. Collagen glycosylation. Curr. Opin. Struct. Biol. 2019, 56, 131–138. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Gabrielli, A.; Avvedimento, E.V.; Krieg, T. Scleroderma. N. Engl. J. Med. 2009, 360, 1989–2003. [Google Scholar] [CrossRef] [PubMed]
- Lauer-Fields, J.L.; Malkar, N.B.; Richet, G.; Drauz, K.; Fields, G.B. Melanoma cell CD44 interaction with the alpha 1(IV)1263-1277 region from basement membrane collagen is modulated by ligand glycosylation. J. Biol. Chem. 2003, 278, 14321–14330. [Google Scholar] [CrossRef] [PubMed]
- Scietti, L.; Chiapparino, A.; De Giorgi, F.; Fumagalli, M.; Khoriauli, L.; Nergadze, S.; Basu, S.; Olieric, V.; Cucca, L.; Banushi, B.; et al. Molecular architecture of the multifunctional collagen lysyl hydroxylase and glycosyltransferase LH3. Nat. Commun. 2018, 9, 3163. [Google Scholar] [CrossRef]
- Guo, H.-F.; Tsai, C.-L.; Miller, M.D.; Philips, G.; Tainer, J.; Kurie, J. The Crystal Structure of a Collagen Galactosylhydroxylysyl Glucosyltransferase from Human. 2021. Available online: https://www.wwpdb.org/pdb?id=pdb_00006wfv (accessed on 25 February 2024).
- Mattoteia, D.; Chiapparino, A.; Fumagalli, M.; De Marco, M.; De Giorgi, F.; Negro, L.; Pinnola, A.; Faravelli, S.; Roscioli, T.; Scietti, L.; et al. Identification of Regulatory Molecular “Hot Spots” for LH/PLOD Collagen Glycosyltransferase Activity. Int. J. Mol. Sci. 2023, 24, 11213. [Google Scholar] [CrossRef] [PubMed]
- Chiapparino, F.; De Giorgi, F.; Scietti, L.; Faravelli, S.; Roscioli, T.; Forneris, F. A cooperative network of molecular “hot spots” highlights the complexity of LH3 collagen glycosyltransferase activities. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bella, J. Collagen structure: New tricks from a very old dog. Biochem. J. 2016, 473, 1001–1025. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Prockop, D.J.; Kivirikko, K.I. Collagens: Molecular biology, diseases, and potentials for therapy. Annu. Rev. Biochem. 1995, 64, 403–434. [Google Scholar] [CrossRef]
- Myllyharju, J.; Kivirikko, K.I. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 2004, 20, 33–43. [Google Scholar] [CrossRef]
- Fratzl, P. (Ed.) Collagen, Structure and Mechanism; Springer Science+Business Media: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Kielty, C.M.; Grant, M.E. The Collagen Family: Structure, Assembly, and Organization in the Extracellular Matrix. In Connective Tissue and Its Heritable Disorders: Molecular, Genetic, and Medical Aspects; Royce, P.M., Steinmann, B., Eds.; Wiley-Liss, Inc.: Hoboken, NJ, USA, 2002; pp. 159–221. [Google Scholar] [CrossRef]
- Sagi, I.; Afratis, N.A. (Eds.) Collagen, Methods and Protocols; Humana: New York, NY, USA, 2019. [Google Scholar] [CrossRef]
- Wang, H. A Review of the Effects of Collagen Treatment in Clinical Studies. Polymers 2021, 13, 3868. [Google Scholar] [CrossRef] [PubMed]
- Abou Neel, E.A.; Bozec, L.; Knowles, J.C.; Syed, O.; Mudera, V.; Day, R.; Hyun, J.K. Collagen–emerging collagen based therapies hit the patient. Adv. Drug Deliv. Rev. 2013, 65, 429–456. [Google Scholar] [CrossRef]
- Onursal, C.; Dick, E.; Angelidis, I.; Schiller, H.B.; Staab-Weijnitz, C.A. Collagen Biosynthesis, Processing, and Maturation in Lung Ageing. Front. Med. 2021, 8, 593874. [Google Scholar] [CrossRef]
- Bornstein, P.; Sage, H. Structurally distinct collagen types. Annu. Rev. Biochem. 1980, 49, 957–1003. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, B.; Persikov, A.V. Fibrous Proteins: Coiled-Coils, Collagen and Elastomers. Adv. Protein Chem. 2005, 70, 301–339. [Google Scholar] [CrossRef]
- Yang, W.; Meyers, M.A.; Ritchie, R.O. Structural architectures with toughening mechanisms in nature: A review of the materials science of type-I collagenous materials. Prog. Mat. Sci. 2019, 103, 425–483. [Google Scholar] [CrossRef]
- Bächinger, H.P.; Mizuno, K.; Vranka, J.A.; Boudko, S.P. Collagen Formation and Structure. Compr. Nat. Prod. II 2010, 5, 469–539. [Google Scholar] [CrossRef]
- Mienaltowski, M.J.; Gonzales, N.L.; Beall, J.M.; Pechanec, M.Y. Basic Structure, Physiology, and Biochemistry of Connective Tissues and Extracellular Matrix Collagens. In Progress in Heritable Soft Connective Tissue Diseases; Springer: Berlin/Heidelberg, Germany, 2021; pp. 5–43. [Google Scholar] [CrossRef]
- Ramshaw, J.A.; Shah, N.K.; Brodsky, B. Gly-X-Y tripeptide frequencies in collagen: A context for host-guest triple-helical peptides. J. Struct. Biol. 1998, 122, 86–91. [Google Scholar] [CrossRef]
- Suzuki, H.; Mahapatra, D.; Board, A.J.; Steel, P.J.; Dyer, J.M.; Gerrard, J.A.; Dobson, R.C.J.; Valery, C. Sub-Angstrom structure of collagen model peptide (GPO)(10) shows a hydrated triple helix with pitch variation and two proline ring conformations. Food Chem. 2020, 319, 126598. [Google Scholar] [CrossRef]
- Berisio, R.; Vitagliano, L.; Mazzarella, L.; Zagari, A. Crystal structure of the collagen triple helix model [(Pro-Pro-Gly)(10)](3). Protein Sci. 2002, 11, 262–270. [Google Scholar] [CrossRef]
- Adzhubei, A.A.; Sternberg, M.J.; Makarov, A.A. Polyproline-II helix in proteins: Structure and function. J. Mol. Biol. 2013, 425, 2100–2132. [Google Scholar] [CrossRef]
- Leitinger, B.; Hohenester, E. Mammalian collagen receptors. Matrix Biol. 2007, 26, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Vesentini, S.; Redaelli, A.; Buehler, M.J. Hierarchical structure and nanomechanics of collagen microfibrils from the atomistic scale up. Nano Lett. 2011, 11, 757–766. [Google Scholar] [CrossRef]
- Petruska, J.A.; Hodge, A.J. A Subunit Model for the Tropocollagen Macromolecule. Proc. Natl. Acad. Sci. USA 1964, 51, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.W. Molecular pattern in native collagen. Nature 1968, 219, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, C.; Luo, X.; Wang, X.; Jiang, H. Recent advances of collagen-based biomaterials: Multi-hierarchical structure, modification and biomedical applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 99, 1509–1522. [Google Scholar] [CrossRef]
- Bozec, L.; van der Heijden, G.; Horton, M. Collagen fibrils: Nanoscale ropes. Biophys. J. 2007, 92, 70–75. [Google Scholar] [CrossRef]
- Sorokina, L.V.; Shahbazian-Yassar, R.; Shokuhfar, T. Collagen biomineralization: Pathways, mechanisms, and thermodynamics. Emergent Mater. 2021, 4, 1205–1224. [Google Scholar] [CrossRef]
- Amirrah, I.N.; Lokanathan, Y.; Zulkiflee, I.; Wee, M.; Motta, A.; Fauzi, M.B. A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines 2022, 10, 2307. [Google Scholar] [CrossRef]
- Koide, T.; Nagata, K. Collagen Biosynthesis. Top. Curr. Chem. 2005, 247, 85–114. [Google Scholar] [CrossRef]
- Salo, A.M.; Myllyharju, J. Prolyl and lysyl hydroxylases in collagen synthesis. Exp. Dermatol. 2021, 30, 38–49. [Google Scholar] [CrossRef]
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef]
- Gjaltema, R.A.; Bank, R.A. Molecular insights into prolyl and lysyl hydroxylation of fibrillar collagens in health and disease. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 74–95. [Google Scholar] [CrossRef]
- Yamauchi, M.; Terajima, M.; Shiiba, M. Lysine Hydroxylation and Cross-Linking of Collagen. Methods Mol. Biol. 2019, 1934, 309–324. [Google Scholar] [CrossRef]
- Kivirriko, K.I. Posttranslational processing of collagens. In Principles of Medical Biology; Bittar, E.E., Bittar, N., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; Volume 3, pp. 233–254. [Google Scholar]
- Kivirikko, K.I.; Myllyla, R. Collagen glycosyltransferases. Int. Rev. Connect. Tissue Res. 1979, 8, 23–72. [Google Scholar] [CrossRef]
- Rappu, P.; Salo, A.M.; Myllyharju, J.; Heino, J. Role of prolyl hydroxylation in the molecular interactions of collagens. Essays Biochem. 2019, 63, 325–335. [Google Scholar] [CrossRef]
- Islam, M.S.; Leissing, T.M.; Chowdhury, R.; Hopkinson, R.J.; Schofield, C.J. 2-Oxoglutarate-Dependent Oxygenases. Annu. Rev. Biochem. 2018, 87, 585–620. [Google Scholar] [CrossRef]
- Myllyharju, J. Collagen Hydroxylases. In 2-Oxoglutarate-Dependent Oxygenases; Royal Society of Chemistry: London, UK, 2015; pp. 149–168. [Google Scholar] [CrossRef]
- Markolovic, S.; Wilkins, S.E.; Schofield, C.J. Protein Hydroxylation Catalyzed by 2-Oxoglutarate-dependent Oxygenases. J. Biol. Chem. 2015, 290, 20712–20722. [Google Scholar] [CrossRef]
- Salo, A.M.; Rappu, P.; Koski, M.K.; Karjalainen, E.; Izzi, V.; Drushinin, K.; Miinalainen, I.; Käpylä, J.; Heino, J.; Myllyharju, J. Collagen prolyl 4-hydroxylases have sequence specificity towards different X-Pro-Gly triplets. bioRxiv 2023. [Google Scholar] [CrossRef]
- Murthy, A.V.; Sulu, R.; Koski, M.K.; Tu, H.; Anantharajan, J.; Sah-Teli, S.K.; Myllyharju, J.; Wierenga, R.K. Structural enzymology binding studies of the peptide-substrate-binding domain of human collagen prolyl 4-hydroxylase (type-II): High affinity peptides have a PxGP sequence motif. Protein Sci. 2018, 27, 1692–1703. [Google Scholar] [CrossRef]
- Myllyharju, J. Prolyl 4-hydroxylases, the key enzymes of collagen biosynthesis. Matrix Biol. 2003, 22, 15–24. [Google Scholar] [CrossRef]
- Sipila, K.H.; Drushinin, K.; Rappu, P.; Jokinen, J.; Salminen, T.A.; Salo, A.M.; Kapyla, J.; Myllyharju, J.; Heino, J. Proline hydroxylation in collagen supports integrin binding by two distinct mechanisms. J. Biol. Chem. 2018, 293, 7645–7658. [Google Scholar] [CrossRef] [PubMed]
- Gorres, K.L.; Raines, R.T. Prolyl 4-hydroxylase. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 106–124. [Google Scholar] [CrossRef] [PubMed]
- Murthy, A.V.; Sulu, R.; Lebedev, A.; Salo, A.M.; Korhonen, K.; Venkatesan, R.; Tu, H.; Bergmann, U.; Janis, J.; Laitaoja, M.; et al. Crystal structure of the collagen prolyl 4-hydroxylase (C-P4H) catalytic domain complexed with PDI: Toward a model of the C-P4H alpha(2)beta(2) tetramer. J. Biol. Chem. 2022, 298, 102614. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Ren, J.; Fang, J.; Ke, H.; Gong, W.; Feng, W.; Wang, C.C. Structural insights into the redox-regulated dynamic conformations of human protein disulfide isomerase. Antioxid. Redox Signal. 2013, 19, 36–45. [Google Scholar] [CrossRef]
- Longbotham, J.E.; Levy, C.; Johannissen, L.O.; Tarhonskaya, H.; Jiang, S.; Loenarz, C.; Flashman, E.; Hay, S.; Schofield, C.J.; Scrutton, N.S. Structure and Mechanism of a Viral Collagen Prolyl Hydroxylase. Biochemistry 2015, 54, 6093–6105. [Google Scholar] [CrossRef] [PubMed]
- Koski, M.K.; Hieta, R.; Hirsila, M.; Ronka, A.; Myllyharju, J.; Wierenga, R.K. The crystal structure of an algal prolyl 4-hydroxylase complexed with a proline-rich peptide reveals a novel buried tripeptide binding motif. J. Biol. Chem. 2009, 284, 25290–25301. [Google Scholar] [CrossRef]
- Anantharajan, J.; Koski, M.K.; Kursula, P.; Hieta, R.; Bergmann, U.; Myllyharju, J.; Wierenga, R.K. The structural motifs for substrate binding and dimerization of the alpha subunit of collagen prolyl 4-hydroxylase. Structure 2013, 21, 2107–2118. [Google Scholar] [CrossRef]
- Koski, M.K.; Anantharajan, J.; Kursula, P.; Dhavala, P.; Murthy, A.V.; Bergmann, U.; Myllyharju, J.; Wierenga, R.K. Assembly of the elongated collagen prolyl 4-hydroxylase alpha(2)beta(2) heterotetramer around a central alpha(2) dimer. Biochem. J. 2017, 474, 751–769. [Google Scholar] [CrossRef]
- Chowdhury, R.; McDonough, M.A.; Mecinovic, J.; Loenarz, C.; Flashman, E.; Hewitson, K.S.; Domene, C.; Schofield, C.J. Structural basis for binding of hypoxia-inducible factor to the oxygen-sensing prolyl hydroxylases. Structure 2009, 17, 981–989. [Google Scholar] [CrossRef]
- Timmins, A.; de Visser, S.P. How Are Substrate Binding and Catalysis Affected by Mutating Glu(127) and Arg(161) in Prolyl-4-hydroxylase? A QM/MM and MD Study. Front. Chem. 2017, 5, 94. [Google Scholar] [CrossRef]
- Stegen, S.; Laperre, K.; Eelen, G.; Rinaldi, G.; Fraisl, P.; Torrekens, S.; Van Looveren, R.; Loopmans, S.; Bultynck, G.; Vinckier, S.; et al. HIF-1alpha metabolically controls collagen synthesis and modification in chondrocytes. Nature 2019, 565, 511–515. [Google Scholar] [CrossRef]
- Vranka, J.A.; Sakai, L.Y.; Bachinger, H.P. Prolyl 3-hydroxylase 1, enzyme characterization and identification of a novel family of enzymes. J. Biol. Chem. 2004, 279, 23615–23621. [Google Scholar] [CrossRef]
- Tiainen, P.; Pasanen, A.; Sormunen, R.; Myllyharju, J. Characterization of recombinant human prolyl 3-hydroxylase isoenzyme 2, an enzyme modifying the basement membrane collagen IV. J. Biol. Chem. 2008, 283, 19432–19439. [Google Scholar] [CrossRef] [PubMed]
- Vranka, J.; Stadler, H.S.; Bachinger, H.P. Expression of prolyl 3-hydroxylase genes in embryonic and adult mouse tissues. Cell Struct. Funct. 2009, 34, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Forlino, A.; Marini, J.C. Osteogenesis imperfecta. Lancet 2016, 387, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Marini, J.C.; Forlino, A.; Bachinger, H.P.; Bishop, N.J.; Byers, P.H.; Paepe, A.; Fassier, F.; Fratzl-Zelman, N.; Kozloff, K.M.; Krakow, D.; et al. Osteogenesis imperfecta. Nat. Rev. Dis. Primers 2017, 3, 17052. [Google Scholar] [CrossRef] [PubMed]
- Weis, M.A.; Hudson, D.M.; Kim, L.; Scott, M.; Wu, J.J.; Eyre, D.R. Location of 3-hydroxyproline residues in collagen types I, II, III, and V/XI implies a role in fibril supramolecular assembly. J. Biol. Chem. 2010, 285, 2580–2590. [Google Scholar] [CrossRef] [PubMed]
- Hudson, D.M.; Eyre, D.R. Collagen prolyl 3-hydroxylation: A major role for a minor post-translational modification? Connect. Tissue Res. 2013, 54, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Rautavuoma, K.; Takaluoma, K.; Passoja, K.; Pirskanen, A.; Kvist, A.P.; Kivirikko, K.I.; Myllyharju, J. Characterization of three fragments that constitute the monomers of the human lysyl hydroxylase isoenzymes 1–3. The 30-kDa N-terminal fragment is not required for lysyl hydroxylase activity. J. Biol. Chem. 2002, 277, 23084–23091. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, J.; Risteli, M.; Lampela, O.; Alavesa, P.; Karppinen, M.; Juffer, A.H.; Myllyla, R. Dimerization of human lysyl hydroxylase 3 (LH3) is mediated by the amino acids 541-547. Matrix Biol. 2011, 30, 27–33. [Google Scholar] [CrossRef]
- Takaluoma, K.; Lantto, J.; Myllyharju, J. Lysyl hydroxylase 2 is a specific telopeptide hydroxylase, while all three isoenzymes hydroxylate collagenous sequences. Matrix Biol. 2007, 26, 396–403. [Google Scholar] [CrossRef]
- Terajima, M.; Taga, Y.; Nakamura, T.; Guo, H.F.; Kayashima, Y.; Maeda-Smithies, N.; Parag-Sharma, K.; Kim, J.S.; Amelio, A.L.; Mizuno, K.; et al. Lysyl hydroxylase 2 mediated collagen post-translational modifications and functional outcomes. Sci. Rep. 2022, 12, 14256. [Google Scholar] [CrossRef]
- Guo, H.F.; Bota-Rabassedas, N.; Terajima, M.; Leticia Rodriguez, B.; Gibbons, D.L.; Chen, Y.; Banerjee, P.; Tsai, C.L.; Tan, X.; Liu, X.; et al. A collagen glucosyltransferase drives lung adenocarcinoma progression in mice. Commun. Biol. 2021, 4, 482. [Google Scholar] [CrossRef] [PubMed]
- Myllyla, R.; Wang, C.; Heikkinen, J.; Juffer, A.; Lampela, O.; Risteli, M.; Ruotsalainen, H.; Salo, A.; Sipila, L. Expanding the lysyl hydroxylase toolbox: New insights into the localization and activities of lysyl hydroxylase 3 (LH3). J. Cell. Physiol. 2007, 212, 323–329. [Google Scholar] [CrossRef]
- WangB, C.; Risteli, M.; Heikkinen, J.; Hussa, A.K.; Uitto, L.; Myllyla, R. Identification of amino acids important for the catalytic activity of the collagen glucosyltransferase associated with the multifunctional lysyl hydroxylase 3 (LH3). J. Biol. Chem. 2002, 277, 18568–18573. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, J.; Risteli, M.; Wang, C.; Latvala, J.; Rossi, M.; Valtavaara, M.; Myllyla, R. Lysyl hydroxylase 3 is a multifunctional protein possessing collagen glucosyltransferase activity. J. Biol. Chem. 2000, 275, 36158–36163. [Google Scholar] [CrossRef]
- WangA, C.; Luosujarvi, H.; Heikkinen, J.; Risteli, M.; Uitto, L.; Myllyla, R. The third activity for lysyl hydroxylase 3: Galactosylation of hydroxylysyl residues in collagens in vitro. Matrix Biol. 2002, 21, 559–566. [Google Scholar] [CrossRef]
- Schegg, B.; Hulsmeier, A.J.; Rutschmann, C.; Maag, C.; Hennet, T. Core glycosylation of collagen is initiated by two beta(1-O)galactosyltransferases. Mol. Cell. Biol. 2009, 29, 943–952. [Google Scholar] [CrossRef]
- Sricholpech, M.; Perdivara, I.; Nagaoka, H.; Yokoyama, M.; Tomer, K.B.; Yamauchi, M. Lysyl hydroxylase 3 glucosylates galactosylhydroxylysine residues in type I collagen in osteoblast culture. J. Biol. Chem. 2011, 286, 8846–8856. [Google Scholar] [CrossRef] [PubMed]
- Sricholpech, M.; Perdivara, I.; Yokoyama, M.; Nagaoka, H.; Terajima, M.; Tomer, K.B.; Yamauchi, M. Lysyl hydroxylase 3-mediated glucosylation in type I collagen: Molecular loci and biological significance. J. Biol. Chem. 2012, 287, 22998–23009. [Google Scholar] [CrossRef]
- Moon, H.J.; Finney, J.; Ronnebaum, T.; Mure, M. Human lysyl oxidase-like 2. Bioorg. Chem. 2014, 57, 231–241. [Google Scholar] [CrossRef]
- Rodriguez-Pascual, F.; Rosell-Garcia, T. Lysyl Oxidases: Functions and Disorders. J. Glaucoma 2018, 27 (Suppl. S1), S15–S19. [Google Scholar] [CrossRef]
- Vallet, S.D.; Ricard-Blum, S. Lysyl oxidases: From enzyme activity to extracellular matrix cross-links. Essays Biochem. 2019, 63, 349–364. [Google Scholar] [CrossRef]
- Csiszar, K. Lysyl oxidases: A novel multifunctional amine oxidase family. Prog. Nucleic Acid Res. Mol. Biol. 2001, 70, 1–32. [Google Scholar] [CrossRef]
- Avery, N.C.; Bailey, A.J. Restraining Cross-Links Responsible for the Mechanical Properties of Collagen Fibers: Natural and Artificial. In Collagen: Structure and Mechanics; Springer: Berlin/Heidelberg, Germany, 2008; pp. 81–110. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Wu, J.; Wang, J.; Shi, Y.; Liu, M. Crystal structure of human lysyl oxidase-like 2 (hLOXL2) in a precursor state. Proc. Natl. Acad. Sci. USA 2018, 115, 3828–3833. [Google Scholar] [CrossRef]
- Vallet, S.D.; Gueroult, M.; Belloy, N.; Dauchez, M.; Ricard-Blum, S. A Three-Dimensional Model of Human Lysyl Oxidase, a Cross-Linking Enzyme. ACS Omega 2020, 4, 8495–8505. [Google Scholar] [CrossRef]
- Steiner, D.; Macron, C.R.; Cohen Sabban, E.N. Diebetes, Non-enzymatic Glycation, and Aging. In Dermatology and Diabetes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 243–279. [Google Scholar] [CrossRef]
- Bailey, A.J. Molecular mechanisms of ageing in connective tissues. Mech. Ageing Dev. 2001, 122, 735–755. [Google Scholar] [CrossRef]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.G.; Bailey, A.J. Glycation of Collagen: The Basis of its Central Role in the Late Complications of Ageing and Diabetes. Int. J. Biochem. Cell Biol. 1996, 28, 1297–1310. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.J.; Paul, R.G.; Knott, L. Mechanisms of maturation and ageing of collagen. Mech. Ageing Dev. 1998, 106, 1–56. [Google Scholar] [CrossRef]
- Zheng, W.; Li, H.; Go, Y.; Chan, X.H.F.; Huang, Q.; Wu, J. Research Advances on the Damage Mechanism of Skin Glycation and Related Inhibitors. Nutrients 2022, 14, 4588. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, F.; Fumagalli, M.; Scietti, L.; Forneris, F. Collagen hydroxylysine glycosylation: Non-conventional substrates for atypical glycosyltransferase enzymes. Biochem. Soc. Trans. 2021, 49, 855–866. [Google Scholar] [CrossRef]
- Monnier, V.M.; Mustata, G.T.; Biemel, K.L.; Reihl, O.; Lederer, M.O.; Zhenyu, D.; Sell, D.R. Cross-linking of the extracellular matrix by the maillard reaction in aging and diabetes: An update on “a puzzle nearing resolution”. Ann. N. Y. Acad. Sci. 2005, 1043, 533–544. [Google Scholar] [CrossRef]
- Hudson, D.M.; Archer, M.; King, K.B.; Eyre, D.R. Glycation of type I collagen selectively targets the same helical domain lysine sites as lysyl oxidase-mediated cross-linking. J. Biol. Chem. 2018, 293, 15620–15627. [Google Scholar] [CrossRef]
- Tvaroska, I.; Kozmon, S.; Kona, J. Molecular Modeling Insights into the Structure and Behavior of Integrins: A Review. Cells 2023, 12, 324. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yun, J.S.; Ko, S.H. Advanced Glycation End Products and Their Effect on Vascular Complications in Type 2 Diabetes Mellitus. Nutrients 2022, 14, 3086. [Google Scholar] [CrossRef] [PubMed]
- Basta, G.; Schmidt, A.M.; De Caterina, R. Advanced glycation end products and vascular inflammation: Implications for accelerated atherosclerosis in diabetes. Cardiovasc. Res. 2004, 63, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.P. The Role of Advanced Glycation End-Products in Cancer Disparity. Adv. Cancer Res. 2017, 133, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Schroter, D.; Hohn, A. Role of Advanced Glycation End Products in Carcinogenesis and their Therapeutic Implications. Curr. Pharm. Des. 2018, 24, 5245–5251. [Google Scholar] [CrossRef] [PubMed]
- Muthyalaiah, Y.S.; Jonnalagadda, B.; John, C.M.; Arockiasamy, S. Impact of Advanced Glycation End products (AGEs) and its receptor (RAGE) on cancer metabolic signaling pathways and its progression. Glycoconj. J. 2021, 38, 717–734. [Google Scholar] [CrossRef] [PubMed]
- Dariya, B.; Nagaraju, G.P. Advanced glycation end products in diabetes, cancer and phytochemical therapy. Drug Discov. Today 2020, 25, 1614–1623. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Tvaroska, I. Glycosyltransferases as targets for therapeutic intervention in cancer and inflammation: Molecular modeling insights. Chem. Pap. 2022, 76, 1953–1988. [Google Scholar] [CrossRef]
- Lowe, J.B.; Marth, J.D. A genetic approach to Mammalian glycan function. Annu. Rev. Biochem. 2003, 72, 643–691. [Google Scholar] [CrossRef]
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities. Biochem. J. 1997, 326 Pt 3, 929–939. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Unligil, U.M.; Rini, J.M. Glycosyltransferase structure and mechanism. Curr. Opin. Struct. Biol. 2000, 10, 510–517. [Google Scholar] [CrossRef]
- Breton, C.; Snajdrova, L.; Jeanneau, C.; Koca, J.; Imberty, A. Structures and mechanisms of glycosyltransferases. Glycobiology 2006, 16, 29r–37r. [Google Scholar] [CrossRef] [PubMed]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.; Tvaroska, I. Carbohydrate-protein interactions: Molecular modeling insights. Adv. Carbohydr. Chem. Biochem. 2014, 71, 9–136. [Google Scholar] [CrossRef] [PubMed]
- Laine, R.A. A calculation of all possible oligosaccharide isomers both branched and linear yields 1.05 x 1012 structures for a reducing hexasaccharide: The Isomer Barrier to development of single-method saccharide sequencing or synthesis systems. Glycobiology 1994, 4, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef]
- Olsen, B.R.; Guzman, N.A.; Engel, J.; Condit, C.; Aase, S. Purification and characterization of a peptide from the carboxy-terminal region of chick tendon procollagen type I. Biochemistry 1977, 16, 3030–3036. [Google Scholar] [CrossRef]
- Clark, C.C. The distribution and initial characterization of oligosaccharide units on the COOH-terminal propeptide extensions of the pro-alpha 1 and pro-alpha 2 chains of type I procollagen. J. Biol. Chem. 1979, 254, 10798–10802. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.C.; Kefalides, N.A. Carbohydrate moieties of procollagen: Incorporation of isotopically labeled mannose and glucosamine into propeptides of procollagen secreted by matrix-free chick embryo tendon cells. Proc. Natl. Acad. Sci. USA 1976, 73, 34–38. [Google Scholar] [CrossRef]
- Li, R.C.; Wong, M.Y.; DiChiara, A.S.; Hosseini, A.S.; Shoulders, M.D. Collagen’s enigmatic, highly conserved N-glycan has an essential proteostatic function. Proc. Natl. Acad. Sci. USA 2021, 118, e2026608118. [Google Scholar] [CrossRef]
- Anttinen, H.; Oikarinen, A.; Ryhanen, L.; Kivirikko, K.I. Evidence for the transfer of mannose to the extension peptides of procollagen within the cisternae of the rough endoplasmic reticulum. FEBS Lett. 1978, 87, 222–226. [Google Scholar] [CrossRef]
- Duksin, D.; Bornstein, P. Impaired conversion of procollagen to collagen by fibroblasts and bone treated with tunicamycin, an inhibitor of protein glycosylation. J. Biol. Chem. 1977, 252, 955–962. [Google Scholar] [CrossRef]
- Lamande, S.R.; Bateman, J.F. The type I collagen pro alpha 1(I) COOH-terminal propeptide N-linked oligosaccharide. Functional analysis by site-directed mutagenesis. J. Biol. Chem. 1995, 270, 17858–17865. [Google Scholar] [CrossRef]
- Clark, C.C. Asparagine-linked glycosides. Methods Enzymol. 1982, 82 Pt A, 346–360. [Google Scholar] [CrossRef]
- Clark, C.C.; Kefalides, N.A. Localization and partial composition of the oligosaccharide units on the propeptide extensions of type I procollagen. J. Biol. Chem. 1978, 253, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Nagae, M.; Yamaguchi, Y.; Taniguchi, N.; Kizuka, Y. 3D Structure and Function of Glycosyltransferases Involved in N-glycan Maturation. Int. J. Mol. Sci. 2020, 21, 437. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P.; Taniguchi, N.; Aebi, M. N-Glycans. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: Harbor, NY, USA, 2015; pp. 99–111. [Google Scholar] [CrossRef]
- Vasconcelos-Dos-Santos, A.; Oliveira, I.A.; Lucena, M.C.; Mantuano, N.R.; Whelan, S.A.; Dias, W.B.; Todeschini, A.R. Biosynthetic Machinery Involved in Aberrant Glycosylation: Promising Targets for Developing of Drugs Against Cancer. Front. Oncol. 2015, 5, 138. [Google Scholar] [CrossRef]
- Aebi, M. N-linked protein glycosylation in the ER. Biochim. Biophys. Acta 2013, 1833, 2430–2437. [Google Scholar] [CrossRef]
- Marti, T.; Kosen, P.A.; Honory, K.; Franzblau, C.; Schmid, K.; Van Halbeek, H.; Gerwig, G.J.; Vliegenthart, J.F. The structure of the carbohydrate units of the carboxyl-terminal peptide of procollagen as elucidated by 500 MHz 1H-NMR spectroscopy. Biochim. Biophys. Acta 1984, 799, 305–312. [Google Scholar] [CrossRef]
- Kornfeld, R.; Kornfeld, S. Assembly of asparagine-linked oligosaccharides. Annu. Rev. Biochem. 1985, 54, 631–664. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, W.; Schleich, H. Uber den Kohlenhydratgehalt des Kollagens. 2. Mitteilung zur Kenntnis des Kollagens. Biochem. Z. 1935, 277, 320–328. [Google Scholar]
- Graßmann, W.; Hörmann, H.; Hafter, R. Eine quantitative Bestimmung von Kohlenhydraten als Osazone. Hoppe-Seyler’s Z. Physiol. Chem. 1957, 307, 87–96. [Google Scholar] [CrossRef]
- Blumenfeld, O.O.; Paz, M.A.; Gallop, P.M.; Seifter, S. The Nature, Quantity, and Mode of Attachment of Hexoses in Ichthyocol. J. Biol. Chem. 1963, 238, 3835–3839. [Google Scholar] [CrossRef]
- Spiro, R.G.; Spiro, M.J. Studies on the biosynthesis of the hydroxylysine-linked disaccharide unit of basement membranes and collagens. 1. Tissue and subcellular distribution of glycosyltransferases and the effect of various conditions on the enzyme levels. J. Biol. Chem. 1971, 246, 4899–4910. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G.; Spiro, M.J. Studies on the biosynthesis of the hydroxylysine-linked disaccharide unit of basement membranes and collagens. 2. Tissue and subcellular distribution of glycosyltransferases and the effect of various conditions on the enzyme levels. J. Biol. Chem. 1971, 246, 4910–4918. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G.; Spiro, M.J. Studies on the biosynthesis of the hydroxylysine-linked disaccharide unit of basement membranes and collagens. 3. Tissue and subcellular distribution of glycosyltransferases and the effect of various conditions on the enzyme levels. J. Biol. Chem. 1971, 246, 4919–4925. [Google Scholar] [CrossRef] [PubMed]
- Bosmann, H.B.; Eylar, E.H. Glycoprotein biosynthesis: The biosynthesis of the hydroxylysine-galactose linkage in collagen. Biochem. Biophys. Res. Commun. 1968, 33, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Bosmann, H.B. Collagen-Glucosyl Transferase in Fibroblasts transformed by Oncogenic Viruses. Nature 1968, 218, 582–583. [Google Scholar] [CrossRef] [PubMed]
- Kivirriko, K.I.; Myllyla, R. Posttranslational enzymes in the biosynthesis of collagen: Intracellular enzymes. Methods Enzymol. 1982, 82, 245–304. [Google Scholar] [CrossRef]
- Liefhebber, J.M.; Punt, S.; Spaan, W.J.; van Leeuwen, H.C. The human collagen beta(1-O)galactosyltransferase, GLT25D1, is a soluble endoplasmic reticulum localized protein. BMC Cell Biol. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Myllyla, R.; Risteli, L.; Kivirikko, K.I. Collagen glucosyltransferase. Partial purification and characterization of the enzyme from whole chick embryos and chick-embryo cartilage. Eur. J. Biochem. 1976, 61, 59–67. [Google Scholar] [CrossRef]
- Risteli, A.L.; Myllya, R.; Kivirikko, K.I. Partial purification and characterization of collagen galactosyltransferase from chick embryos. Biochem. J. 1976, 155, 145–153. [Google Scholar] [CrossRef]
- Risteli, B.L.; Myllyla, R.; Kivirikko, K.I. Affinity chromatography of collagen glycosyltransferases on collagen linked to agarose. Eur. J. Biochem. 1976, 67, 197–202. [Google Scholar] [CrossRef]
- Perrin-Tricaud, C.; Rutschmann, C.; Hennet, T. Identification of domains and amino acids essential to the collagen galactosyltransferase activity of GLT25D1. PLoS ONE 2011, 6, e29390. [Google Scholar] [CrossRef] [PubMed]
- Breton, C.; Imberty, A. Structure/function studies of glycosyltransferases. Curr. Opin. Struct. Biol. 1999, 9, 563–571. [Google Scholar] [CrossRef]
- Qasba, P.K.; Ramakrishnan, B.; Boeggeman, E. Substrate-induced conformational changes in glycosyltransferases. Trends Biochem. Sci. 2005, 30, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Unligil, U.M.; Zhou, S.; Yuwaraj, S.; Sarkar, M.; Schachter, H.; Rini, J.M. X-ray crystal structure of rabbit N-acetylglucosaminyltransferase I: Catalytic mechanism and a new protein superfamily [In Process Citation]. EMBO J. 2000, 19, 5269–5280. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.F.; Tsai, C.L.; Terajima, M.; Tan, X.; Banerjee, P.; Miller, M.D.; Liu, X.; Yu, J.; Byemerwa, J.; Alvarado, S.; et al. Pro-metastatic collagen lysyl hydroxylase dimer assemblies stabilized by Fe(2+)-binding. Nat. Commun. 2018, 9, 512. [Google Scholar] [CrossRef]
- Rautavuoma, K.; Takaluoma, K.; Sormunen, R.; Myllyharju, J.; Kivirikko, K.I.; Soininen, R. Premature aggregation of type IV collagen and early lethality in lysyl hydroxylase 3 null mice. Proc. Natl. Acad. Sci. USA 2004, 101, 14120–14125. [Google Scholar] [CrossRef]
- Salo, A.M.; Wang, C.; Sipila, L.; Sormunen, R.; Vapola, M.; Kervinen, P.; Ruotsalainen, H.; Heikkinen, J.; Myllyla, R. Lysyl hydroxylase 3 (LH3) modifies proteins in the extracellular space, a novel mechanism for matrix remodeling. J. Cell. Physiol. 2006, 207, 644–653. [Google Scholar] [CrossRef]
- Aik, W.; McDonough, M.A.; Thalhammer, A.; Chowdhury, R.; Schofield, C.J. Role of the jelly-roll fold in substrate binding by 2-oxoglutarate oxygenases. Curr. Opin. Struct. Biol. 2012, 22, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, H.; Sipila, L.; Vapola, M.; Sormunen, R.; Salo, A.M.; Uitto, L.; Mercer, D.K.; Robins, S.P.; Risteli, M.; Aszodi, A.; et al. Glycosylation catalyzed by lysyl hydroxylase 3 is essential for basement membranes. J. Cell Sci. 2006, 119, 625–635. [Google Scholar] [CrossRef]
- Gómez, H.; Rojas, R.; Patel, D.; Tabak, L.A.; Lluch, J.M.; Masgrau, L. A computational and experimental study of O-glycosylation. Catalysis by human UDP-GalNAc polypeptide: GalNAc transferase-T2. Org. Biomol. Chem. 2014, 12, 2645–2655. [Google Scholar] [CrossRef]
- Tvaroska, I. Molecular modeling of retaining glycosyltransferases. ACS Symp. Ser. 2006, 930, 285–301. [Google Scholar] [CrossRef]
- Andre, I.; Tvaroska, I.; Carver, J.P. On the reaction pathways and determination of transition-state structures for retaining alpha-galactosyltransferases. Carbohydr. Res. 2003, 338, 865–877. [Google Scholar] [CrossRef]
- Monegal, A.; Planas, A. Chemical rescue of alpha 3-galactosyltransferase. Implications in the mechanism of retaining glycosyltransferases. J. Am. Chem. Soc. 2006, 128, 16030–16031. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Cervellera, V.; Ardevol, A.; Boero, M.; Planas, A.; Rovira, C. Formation of a covalent glycosyl-enzyme species in a retaining glycosyltransferase. Chemistry 2013, 19, 14018–14023. [Google Scholar] [CrossRef] [PubMed]
- Soya, N.; Fang, Y.; Palcic, M.M.; Klassen, J.S. Trapping and characterization of covalent intermediates of mutant retaining glycosyltransferases. Glycobiology 2011, 21, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Persson, K.; Ly, H.D.; Dieckelmann, M.; Wakarchuk, W.W.; Withers, S.G.; Strynadka, N.C.J. Crystal structure of the retaining galactosyltransferase LgtC from Neisseria meningitidis in complex with donor and acceptor sugar analogs: Towards a mechanism. Nat. Struct. Biol. 2001, 8, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Tvaroška, I. Molecular modeling insights into the catalytic mechanism of the retining galactosyltransferase LgtC. Carbohydr. Res. 2004, 339, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Gomez, H.; Polyak, I.; Thiel, W.; Lluch, J.M.; Masgrau, L. Retaining Glycosyltransferase Mechanism Studied by QM/MM Methods: Lipopolysaccharyl-alpha-1,4-galactosyltransferase C Transfers alpha-Galactose via an Oxocarbenium Ion-like Transition State. J. Am. Chem. Soc. 2012, 134, 4743–4752. [Google Scholar] [CrossRef] [PubMed]
- Gómez, H.; Lluch, J.M.; Masgrau, L. Substrate-Assisted and Nucleophilically Assisted Catalysis in Bovine alpha 1,3-Galactosyltransferase. Mechanistic Implications for Retaining Glycosyltransferases. J. Am. Chem. Soc. 2013, 135, 7053–7063. [Google Scholar] [CrossRef] [PubMed]
- Albesa-Jove, D.; Sainz-Polo, M.A.; Marina, A.; Guerin, M.E. Structural Snapshots of alpha-1,3-Galactosyltransferase with Native Substrates: Insight into the Catalytic Mechanism of Retaining Glycosyltransferases. Angew. Chem. Int. Ed. Engl. 2017, 56, 14853–14857. [Google Scholar] [CrossRef] [PubMed]
- Ardevol, A.; Rovira, C. The Molecular Mechanism of Enzymatic Glycosyl Transfer with Retention of Configuration: Evidence for a Short-Lived Oxocarbenium-Like Species. Angew. Chem.-Int. Ed. 2011, 50, 10897–10901. [Google Scholar] [CrossRef] [PubMed]
- Bobovska, A.; Tvaroska, I.; Kona, J. A theoretical study on the catalytic mechanism of the retaining alpha-1,2-mannosyltransferase Kre2p/Mnt1p: The impact of different metal ions on catalysis. Org. Biomol. Chem. 2014, 12, 4201–4210. [Google Scholar] [CrossRef] [PubMed]
- Janos, P.; Trnka, T.; Kozmon, S.; Tvaroska, I.; Koca, J. Different QM/MM Approaches To Elucidate Enzymatic Reactions: Case Study on ppGalNAcT2. J. Chem. Theory. Comput. 2016, 12, 6062–6076. [Google Scholar] [CrossRef]
- Lira-Navarrete, E.; Iglesias-Fernandez, J.; Zandberg, W.F.; Companon, I.; Kong, Y.; Corzana, F.; Pinto, B.M.; Clausen, H.; Peregrina, J.M.; Vocadlo, D.J.; et al. Substrate-guided front-face reaction revealed by combined structural snapshots and metadynamics for the polypeptide N-acetylgalactosaminyltransferase 2. Angew. Chem. Int. Ed. Engl. 2014, 53, 8206–8210. [Google Scholar] [CrossRef]
- Trnka, T.; Kozmon, S.; Tvaroska, I.; Koca, J. Stepwise catalytic mechanism via short-lived intermediate inferred from combined QM/MM MERP and PES calculations on retaining glycosyltransferase ppGalNAcT2. PLoS Comput. Biol. 2015, 11, e1004061. [Google Scholar] [CrossRef]
- Sternberg, M.; Spiro, R.G. Studies on the catabolism of the hydroxylysine-linked disaccharide units of basement membranes and collagens. Isolation and characterization of a rat kidney alpha-glucosidase of high specificity. J. Biol. Chem. 1979, 254, 10329–10336. [Google Scholar] [CrossRef]
- Sternberg, M.; Spiro, R.G. Studies on the Catabolism of the Hydroxylysine-Linked Disaccharide Units of Basement Membranes and Collagens: Isolation and Characterization of a New Rat-Kidney α-Glucosidase of High Specificity. Ren. Physiol. 1980, 3, 1–3. [Google Scholar] [CrossRef]
- Hamazaki, H.; Hotta, K. Purification and characterization of an alpha-glucosidase specific for hydroxylysine-linked disaccharide of collagen. J. Biol. Chem. 1979, 254, 9682–9687. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, H.; Hamazaki, M.H. Catalytic site of human protein-glucosylgalactosylhydroxylysine glucosidase: Three crucial carboxyl residues were determined by cloning and site-directed mutagenesis. Biochem. Biophys. Res. Commun. 2016, 469, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Nihira, T.; Kurata, R.; Nakai, H.; Funane, K.; Park, E.Y.; Miyazaki, T. Structure of a bacterial alpha-1,2-glucosidase defines mechanisms of hydrolysis and substrate specificity in GH65 family hydrolases. J. Biol. Chem. 2021, 297, 101366. [Google Scholar] [CrossRef]
- Fang, M.; Yuan, J.; Peng, C.; Li, Y. Collagen as a double-edged sword in tumor progression. Tumour Biol. 2014, 35, 2871–2882. [Google Scholar] [CrossRef]
- Stawikowski, M.J.; Aukszi, B.; Stawikowska, R.; Cudic, M.; Fields, G.B. Glycosylation modulates melanoma cell alpha2beta1 and alpha3beta1 integrin interactions with type IV collagen. J. Biol. Chem. 2014, 289, 21591–21604. [Google Scholar] [CrossRef]
- Tang, M.; Wang, X.; Gandhi, N.S.; Foley, B.L.; Burrage, K.; Woods, R.J.; Gu, Y. Effect of hydroxylysine-O-glycosylation on the structure of type I collagen molecule: A computational study. Glycobiology 2020, 30, 830–843. [Google Scholar] [CrossRef]
- An, H.J.; Froehlich, J.W.; Lebrilla, C.B. Determination of glycosylation sites and site-specific heterogeneity in glycoproteins. Curr. Opin. Chem. Biol. 2009, 13, 421–426. [Google Scholar] [CrossRef]
- Perdivara, I.; Yamauchi, M.; Tomer, K.B. Molecular Characterization of Collagen Hydroxylysine O-Glycosylation by Mass Spectrometry: Current Status. Aust. J. Chem. 2013, 66, 760–769. [Google Scholar] [CrossRef]
- Sharma, U.; Carrique, L.; Vadon-Le Goff, S.; Mariano, N.; Georges, R.N.; Delolme, F.; Koivunen, P.; Myllyharju, J.; Moali, C.; Aghajari, N.; et al. Structural basis of homo- and heterotrimerization of collagen I. Nat. Commun. 2017, 8, 14671. [Google Scholar] [CrossRef]
- Davies, D.; Tuckwell, D.S.; Calderwood, D.A.; Weston, S.A.; Takigawa, M.; Humphries, M.J. Molecular characterisation of integrin-procollagen C-propeptide interactions. Eur. J. Biochem. 1997, 246, 274–282. [Google Scholar] [CrossRef]
- Katayama, K.; Armendariz-Borunda, J.; Raghow, R.; Kang, A.H.; Seyer, J.M. A pentapeptide from type I procollagen promotes extracellular matrix production. J. Biol. Chem. 1993, 268, 9941–9944. [Google Scholar] [CrossRef] [PubMed]
- Smedsrod, B.; Melkko, J.; Risteli, L.; Risteli, J. Circulating C-terminal propeptide of type I procollagen is cleared mainly via the mannose receptor in liver endothelial cells. Biochem. J. 1990, 271, 345–350. [Google Scholar] [CrossRef]
- Notbohm, H.; Nokelainen, M.; Myllyharju, J.; Fietzek, P.P.; Muller, P.K.; Kivirikko, K.I. Recombinant human type II collagens with low and high levels of hydroxylysine and its glycosylated forms show marked differences in fibrillogenesis in vitro. J. Biol. Chem. 1999, 274, 8988–8992. [Google Scholar] [CrossRef]
- Torre-Blanco, A.; Adachi, E.; Hojima, Y.; Wootton, J.A.; Minor, R.R.; Prockop, D.J. Temperature-induced post-translational over-modification of type I procollagen. Effects of over-modification of the protein on the rate of cleavage by procollagen N-proteinase and on self-assembly of collagen into fibrils. J. Biol. Chem. 1992, 267, 2650–2655. [Google Scholar] [CrossRef]
- Batge, B.; Winter, C.; Notbohm, H.; Acil, Y.; Brinckmann, J.; Muller, P.K. Glycosylation of human bone collagen I in relation to lysylhydroxylation and fibril diameter. J. Biochem. 1997, 122, 109–115. [Google Scholar] [CrossRef]
- Sipila, L.; Ruotsalainen, H.; Sormunen, R.; Baker, N.L.; Lamande, S.R.; Vapola, M.; Wang, C.; Sado, Y.; Aszodi, A.; Myllyla, R. Secretion and assembly of type IV and VI collagens depend on glycosylation of hydroxylysines. J. Biol. Chem. 2007, 282, 33381–33388. [Google Scholar] [CrossRef]
- Risteli, M.; Ruotsalainen, H.; Salo, A.M.; Sormunen, R.; Sipila, L.; Baker, N.L.; Lamande, S.R.; Vimpari-Kauppinen, L.; Myllyla, R. Reduction of lysyl hydroxylase 3 causes deleterious changes in the deposition and organization of extracellular matrix. J. Biol. Chem. 2009, 284, 28204–28211. [Google Scholar] [CrossRef]
- Jurgensen, H.J.; Madsen, D.H.; Ingvarsen, S.; Melander, M.C.; Gardsvoll, H.; Patthy, L.; Engelholm, L.H.; Behrendt, N. A novel functional role of collagen glycosylation: Interaction with the endocytic collagen receptor uparap/ENDO180. J. Biol. Chem. 2011, 286, 32736–32748. [Google Scholar] [CrossRef]
- Gardelli, C.; Russo, L.; Cipolla, L.; Moro, M.; Andriani, F.; Rondinone, O.; Nicotra, F.; Sozzi, G.; Bertolini, G.; Roz, L. Differential glycosylation of collagen modulates lung cancer stem cell subsets through beta1 integrin-mediated interactions. Cancer Sci. 2021, 112, 217–230. [Google Scholar] [CrossRef]
- Wang, C.; Kovanen, V.; Raudasoja, P.; Eskelinen, S.; Pospiech, H.; Myllyla, R. The glycosyltransferase activities of lysyl hydroxylase 3 (LH3) in the extracellular space are important for cell growth and viability. J. Cell. Mol. Med. 2009, 13, 508–521. [Google Scholar] [CrossRef]
- Myers, L.K.; Myllyharju, J.; Nokelainen, M.; Brand, D.D.; Cremer, M.A.; Stuart, J.M.; Bodo, M.; Kivirikko, K.I.; Kang, A.H. Relevance of posttranslational modifications for the arthritogenicity of type II collagen. J. Immunol. 2004, 172, 2970–2975. [Google Scholar] [CrossRef]
- Myllyharju, J.; Kivirikko, K.I. Collagens and collagen-related diseases. Ann. Med. 2001, 33, 7–21. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Baffet, G.; Theret, N. Molecular and tissue alterations of collagens in fibrosis. Matrix Biol. 2018, 68–69, 122–149. [Google Scholar] [CrossRef]
- Williams, L.; Layton, T.; Yang, N.; Feldmann, M.; Nanchahal, J. Collagen VI as a driver and disease biomarker in human fibrosis. FEBS J. 2022, 289, 3603–3629. [Google Scholar] [CrossRef]
- Muschler, J.; Streuli, C.H. Cell-matrix interactions in mammary gland development and breast cancer. Cold Spring Harb. Perspect. Biol. 2010, 2, a003202. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Molecular pathways: Connecting fibrosis and solid tumor metastasis. Clin. Cancer. Res. 2014, 20, 3637–3643. [Google Scholar] [CrossRef]
- Wagner, G.K.; Pesnot, T. Glycosyltransferases and their Assays. Chembiochem 2010, 11, 1939–1949. [Google Scholar] [CrossRef]
- Li, Q.; Yang, Y.; Reis, C.; Tao, T.; Li, W.; Li, X.; Zhang, J.H. Cerebral Small Vessel Disease. Cell Transplant. 2018, 27, 1711–1722. [Google Scholar] [CrossRef]
- Gould, D.B.; Phalan, F.C.; van Mil, S.E.; Sundberg, J.P.; Vahedi, K.; Massin, P.; Bousser, M.G.; Heutink, P.; Miner, J.H.; Tournier-Lasserve, E.; et al. Role of COL4A1 in small-vessel disease and hemorrhagic stroke. N. Engl. J. Med. 2006, 354, 1489–1496. [Google Scholar] [CrossRef]
- Weng, Y.C.; Sonni, A.; Labelle-Dumais, C.; de Leau, M.; Kauffman, W.B.; Jeanne, M.; Biffi, A.; Greenberg, S.M.; Rosand, J.; Gould, D.B. COL4A1 mutations in patients with sporadic late-onset intracerebral hemorrhage. Ann. Neurol. 2012, 71, 470–477. [Google Scholar] [CrossRef]
- Jeanne, M.; Labelle-Dumais, C.; Jorgensen, J.; Kauffman, W.B.; Mancini, G.M.; Favor, J.; Valant, V.; Greenberg, S.M.; Rosand, J.; Gould, D.B. COL4A2 mutations impair COL4A1 and COL4A2 secretion and cause hemorrhagic stroke. Am. J. Hum. Genet. 2012, 90, 91–101. [Google Scholar] [CrossRef]
- Miyatake, S.; Schneeberger, S.; Koyama, N.; Yokochi, K.; Ohmura, K.; Shiina, M.; Mori, H.; Koshimizu, E.; Imagawa, E.; Uchiyama, Y.; et al. Biallelic COLGALT1 variants are associated with cerebral small vessel disease. Ann. Neurol. 2018, 84, 843–853. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Selected Heritable Disorders of Connective Tissue and Disability; The National Academies Press: Washington, DC, USA, 2022. [Google Scholar] [CrossRef]
- Salo, A.M.; Cox, H.; Farndon, P.; Moss, C.; Grindulis, H.; Risteli, M.; Robins, S.P.; Myllyla, R. A connective tissue disorder caused by mutations of the lysyl hydroxylase 3 gene. Am. J. Hum. Genet. 2008, 83, 495–503. [Google Scholar] [CrossRef]
- Ewans, L.J.; Colley, A.; Gaston-Massuet, C.; Gualtieri, A.; Cowley, M.J.; McCabe, M.J.; Anand, D.; Lachke, S.A.; Scietti, L.; Forneris, F.; et al. Pathogenic variants in PLOD3 result in a Stickler syndrome-like connective tissue disorder with vascular complications. J. Med. Genet. 2019, 56, 629–638. [Google Scholar] [CrossRef]
- Vahidnezhad, H.; Youssefian, L.; Saeidian, A.H.; Touati, A.; Pajouhanfar, S.; Baghdadi, T.; Shadmehri, A.A.; Giunta, C.; Kraenzlin, M.; Syx, D.; et al. Mutations in PLOD3, encoding lysyl hydroxylase 3, cause a complex connective tissue disorder including recessive dystrophic epidermolysis bullosa-like blistering phenotype with abnormal anchoring fibrils and type VII collagen deficiency. Matrix Biol. 2019, 81, 91–106. [Google Scholar] [CrossRef]
- Giunta, C.; Randolph, A.; Al-Gazali, L.I.; Brunner, H.G.; Kraenzlin, M.E.; Steinmann, B. Nevo syndrome is allelic to the kyphoscoliotic type of the Ehlers-Danlos syndrome (EDS VIA). Am. J. Med. Genet. A 2005, 133, 158–164. [Google Scholar] [CrossRef]
- Beighton, P.; De Paepe, A.; Steinmann, B.; Tsipouras, P.; Wenstrup, R.J. Ehlers-Danlos syndromes: Revised nosology, Villefranche, 1997. Ehlers-Danlos National Foundation (USA) and Ehlers-Danlos Support Group (UK). Am. J. Med. Genet. 1998, 77, 31–37. [Google Scholar] [CrossRef]
- Steinmann, B.; Royce, P.M.; Superti-Furga, A. Connective Tissue and Its Heritable Disorders: Molecular, Genetic, and Medical Aspects; Wiley-Liss, Inc.: Hoboken, NJ, USA, 2002. [Google Scholar] [CrossRef]
- Yeowell, H.N.; Walker, L.C. Mutations in the lysyl hydroxylase 1 gene that result in enzyme deficiency and the clinical phenotype of Ehlers-Danlos syndrome type VI. Mol. Genet. Metab. 2000, 71, 212–224. [Google Scholar] [CrossRef]
- Ha-Vinh, R.; Alanay, Y.; Bank, R.A.; Campos-Xavier, A.B.; Zankl, A.; Superti-Furga, A.; Bonafe, L. Phenotypic and molecular characterization of Bruck syndrome (osteogenesis imperfecta with contractures of the large joints) caused by a recessive mutation in PLOD2. Am. J. Med. Genet. A 2004, 131, 115–120. [Google Scholar] [CrossRef]
- van der Slot, A.J.; Zuurmond, A.M.; Bardoel, A.F.; Wijmenga, C.; Pruijs, H.E.; Sillence, D.O.; Brinckmann, J.; Abraham, D.J.; Black, C.M.; Verzijl, N.; et al. Identification of PLOD2 as telopeptide lysyl hydroxylase, an important enzyme in fibrosis. J. Biol. Chem. 2003, 278, 40967–40972. [Google Scholar] [CrossRef]
- Puig-Hervas, M.T.; Temtamy, S.; Aglan, M.; Valencia, M.; Martinez-Glez, V.; Ballesta-Martinez, M.J.; Lopez-Gonzalez, V.; Ashour, A.M.; Amr, K.; Pulido, V.; et al. Mutations in PLOD2 cause autosomal-recessive connective tissue disorders within the Bruck syndrome--osteogenesis imperfecta phenotypic spectrum. Hum. Mutat. 2012, 33, 1444–1449. [Google Scholar] [CrossRef]
- Luce, L.; Casale, M.; Waldron, S. A Rare Case of Bruck Syndrome Type 2 in Siblings with Broad Phenotypic Variability. Ochsner J. 2020, 20, 204–208. [Google Scholar] [CrossRef]
- Uitto, J.; Richard, G. Progress in epidermolysis bullosa: From eponyms to molecular genetic classification. Clin. Dermatol. 2005, 23, 33–40. [Google Scholar] [CrossRef]
- Savolainen, E.R.; Kero, M.; Pihlajaniemi, T.; Kivirikko, K.I. Deficiency of galactosylhydroxylysyl glucosyltransferase, an enzyme of collagen synthesis, in a family with dominant epidermolysis bullosa simplex. N. Engl. J. Med. 1981, 304, 197–204. [Google Scholar] [CrossRef]
- Snead, M.P.; Yates, J.R. Clinical and Molecular genetics of Stickler syndrome. J. Med. Genet. 1999, 36, 353–359. [Google Scholar] [CrossRef]
- Soh, Z.; Richards, A.J.; McNinch, A.; Alexander, P.; Martin, H.; Snead, M.P. Dominant Stickler Syndrome. Genes 2022, 13, 1089. [Google Scholar] [CrossRef]
- Boothe, M.; Morris, R.; Robin, N. Stickler Syndrome: A Review of Clinical Manifestations and the Genetics Evaluation. J. Pers. Med. 2020, 10, 105. [Google Scholar] [CrossRef]
- Bank, R.A.; Verzijl, N.; Lafeber, F.P.; Tekoppele, J.M. Putative role of lysyl hydroxylation and pyridinoline cross-linking during adolescence in the occurrence of osteoarthritis at old age. Osteoarthr. Cartil. 2002, 10, 127–134. [Google Scholar] [CrossRef]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef]
- Bank, R.A.; TeKoppele, J.M.; Oostingh, G.; Hazleman, B.L.; Riley, G.P. Lysylhydroxylation and non-reducible crosslinking of human supraspinatus tendon collagen: Changes with age and in chronic rotator cuff tendinitis. Ann. Rheum. Dis. 1999, 58, 35–41. [Google Scholar] [CrossRef]
- Xu, Y.; Murrell, G.A. The basic science of tendinopathy. Clin. Orthop. Relat. Res. 2008, 466, 1528–1538. [Google Scholar] [CrossRef]
- de Mos, M.; van El, B.; DeGroot, J.; Jahr, H.; van Schie, H.T.; van Arkel, E.R.; Tol, H.; Heijboer, R.; van Osch, G.J.; Verhaar, J.A. Achilles tendinosis: Changes in biochemical composition and collagen turnover rate. Am. J. Sports Med. 2007, 35, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Terajima, M.; Yang, Y.; Sun, L.; Ahn, Y.H.; Pankova, D.; Puperi, D.S.; Watanabe, T.; Kim, M.P.; Blackmon, S.H.; et al. Lysyl hydroxylase 2 induces a collagen cross-link switch in tumor stroma. J. Clin. Investig. 2015, 125, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Eisinger-Mathason, T.S.; Zhang, M.; Qiu, Q.; Skuli, N.; Nakazawa, M.S.; Karakasheva, T.; Mucaj, V.; Shay, J.E.; Stangenberg, L.; Sadri, N.; et al. Hypoxia-dependent modification of collagen networks promotes sarcoma metastasis. Cancer Discov. 2013, 3, 1190–1205. [Google Scholar] [CrossRef] [PubMed]
- Gilkes, D.M.; Bajpai, S.; Wong, C.C.; Chaturvedi, P.; Hubbi, M.E.; Wirtz, D.; Semenza, G.L. Procollagen lysyl hydroxylase 2 is essential for hypoxia-induced breast cancer metastasis. Mol. Cancer Res. 2013, 11, 456–466. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tvaroška, I. Glycosylation Modulates the Structure and Functions of Collagen: A Review. Molecules 2024, 29, 1417. https://doi.org/10.3390/molecules29071417
Tvaroška I. Glycosylation Modulates the Structure and Functions of Collagen: A Review. Molecules. 2024; 29(7):1417. https://doi.org/10.3390/molecules29071417
Chicago/Turabian StyleTvaroška, Igor. 2024. "Glycosylation Modulates the Structure and Functions of Collagen: A Review" Molecules 29, no. 7: 1417. https://doi.org/10.3390/molecules29071417
APA StyleTvaroška, I. (2024). Glycosylation Modulates the Structure and Functions of Collagen: A Review. Molecules, 29(7), 1417. https://doi.org/10.3390/molecules29071417