
Effects of Active Ingredients in Alcoholic Beverages and Their De-Alcoholized Counterparts on High-Fat Diet Bees: A Comparative Study


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Phenols, Monosaccharides, Amino Acids, and Organic Acids in Alcoholic Beverage Samples and in Dietary Samples from Different Groups of Bees
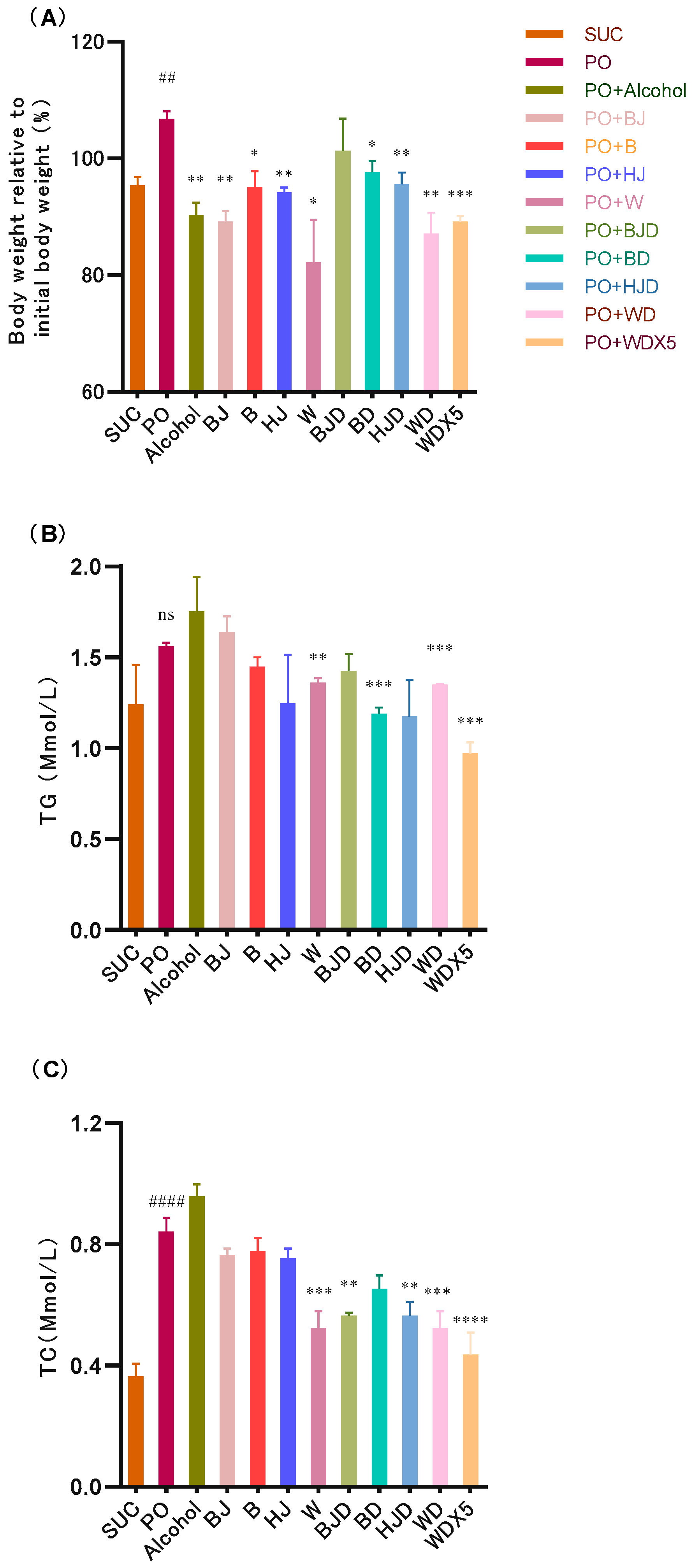
2.2. Alcohol Tends to Exacerbate Metabolic Disorders in High-Fat-Diet-Induced Bees
2.3. Alcoholic Beverages and Their Components Alleviate Metabolic Disorders in High-Fat-Diet-Induced Bees
2.4. Consuming Non-Alcoholic Alcoholic Beverages Can Help Avoid the Adverse Effects of Alcohol While Improving Metabolic Disruptions
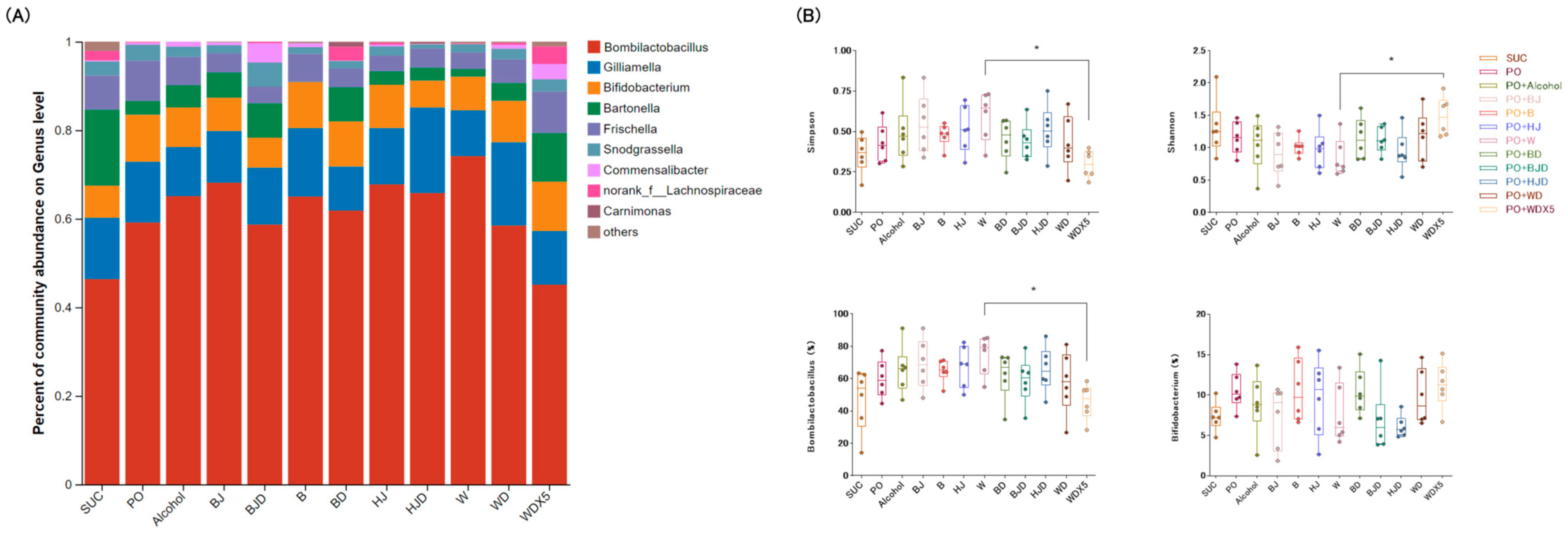
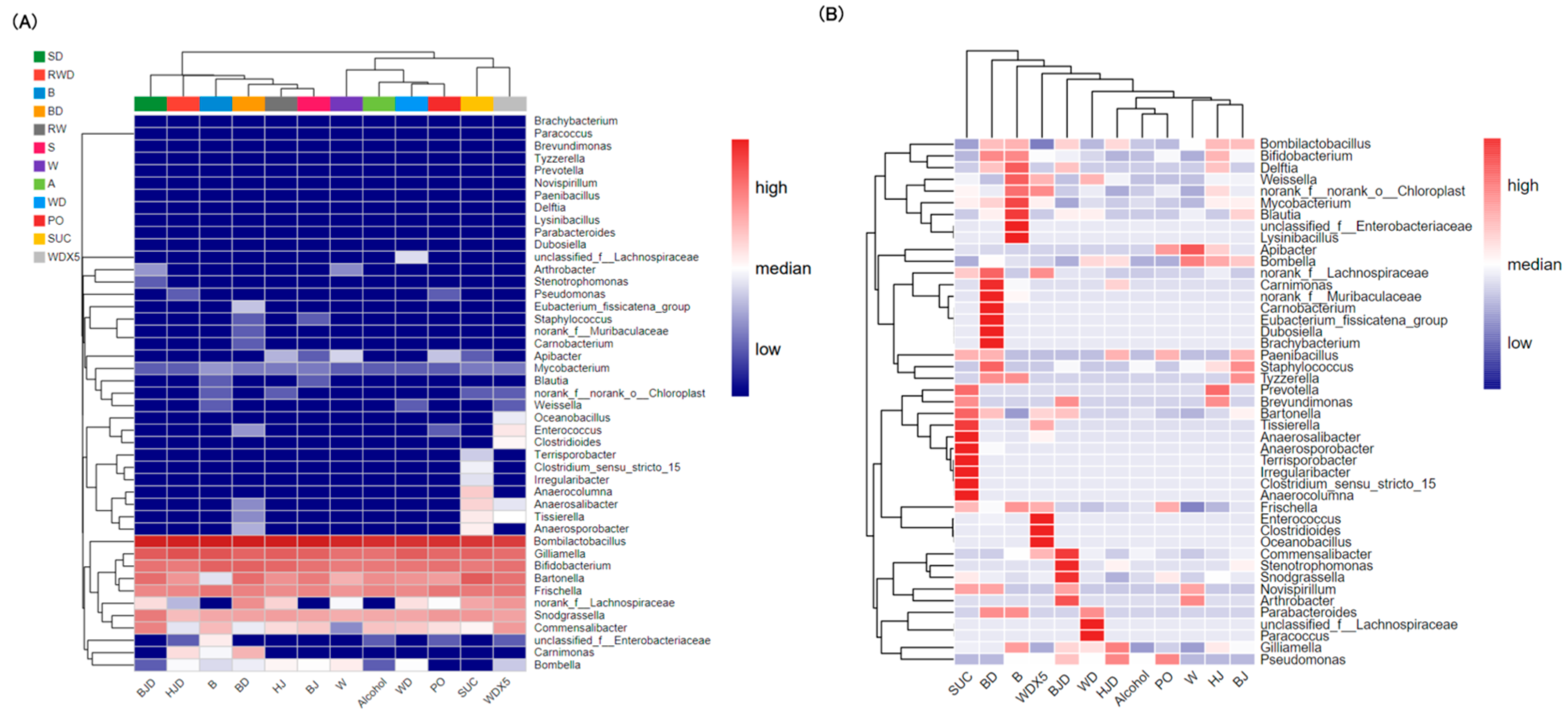
2.5. Wine Balances the Proportions of Gut Microbiota
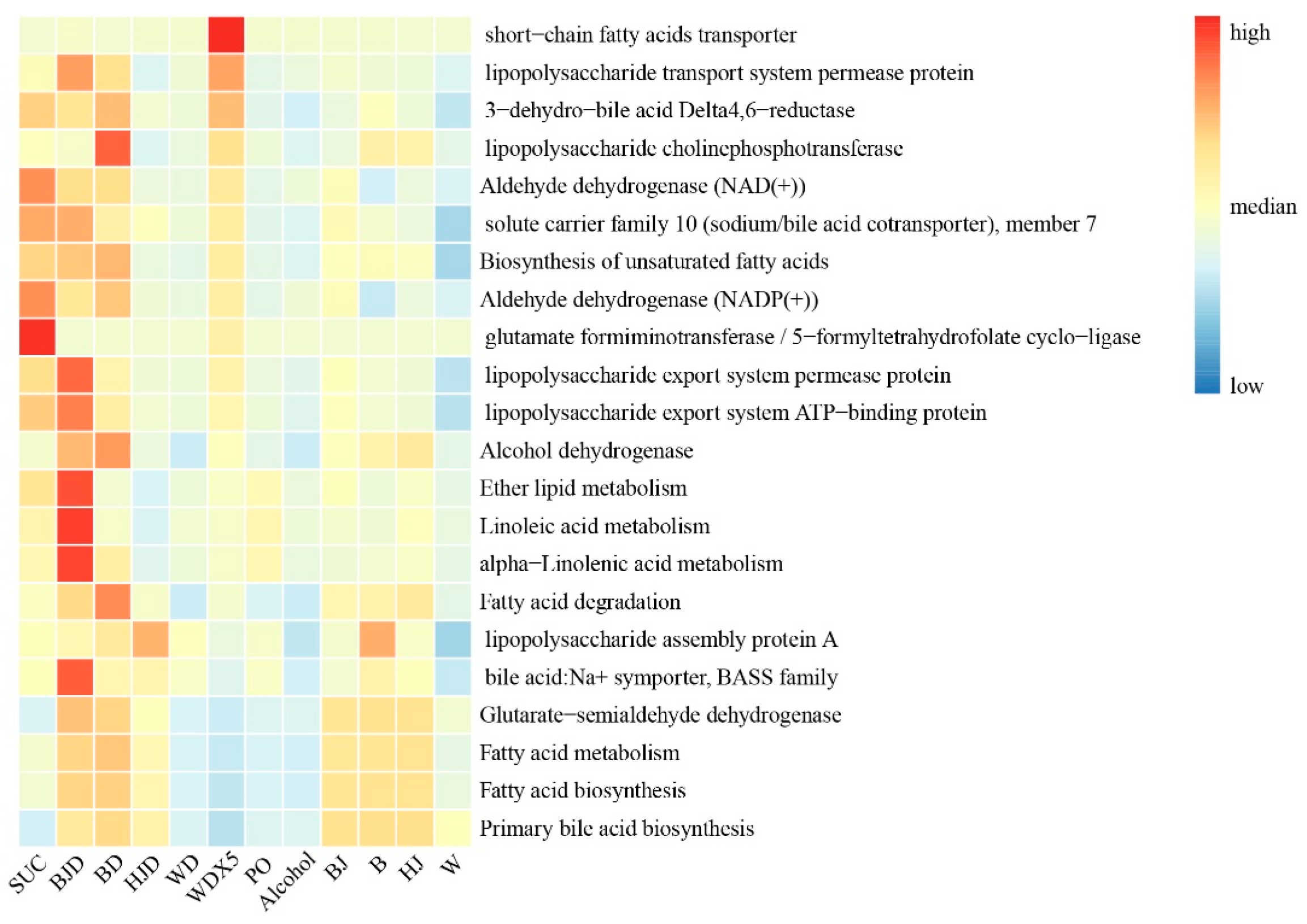
2.6. WDX5 Improved Metabolic Function
3. Discussion
4. Materials and Methods
4.1. De-Alcoholization of Alcoholic Beverages
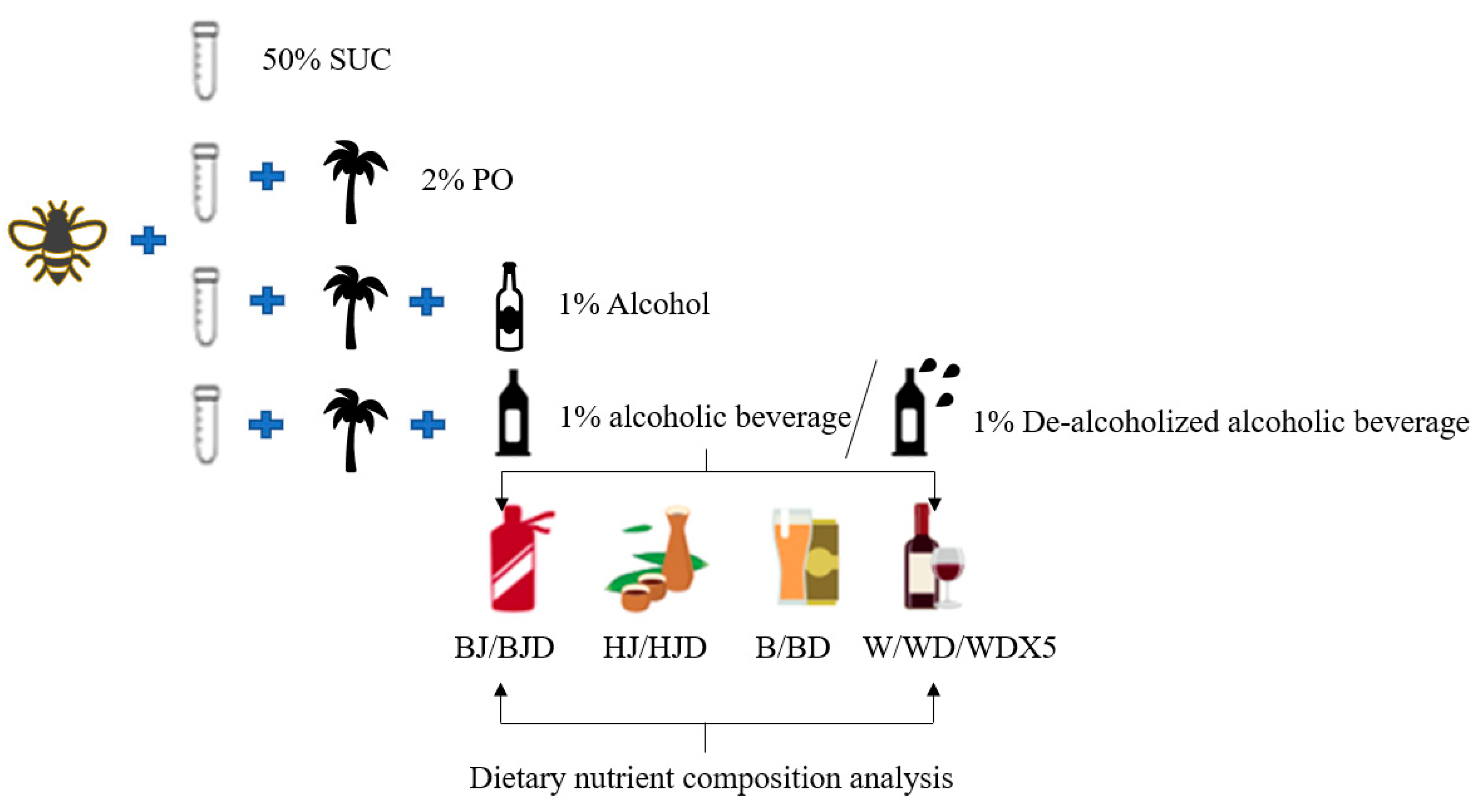
4.2. Alcoholic Beverages and Their De-Alcoholized Counterparts and Bee Diet Composition Analysis
4.3. Animal Experiments
4.4. Determination of Hemolymph TG and TC in Bees
4.5. Gut Microbiota Analysis by 16SrRNA Sequencing
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ngowi, E.E.; Wang, Y.; Khattak, S.; Khan, N.H.; Mahmoud, S.S.M.; Helmy, Y.A.S.H.; Jiang, Q.; Li, T.; Duan, S.; Ji, X.; et al. Impact of the factors shaping gut microbiota on obesity. J. Appl. Microbiol. 2021, 131, 2131–2147. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. Gut microbiota—At the intersection of everything? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Houtman, T.A.; Eckermann, H.A.; Smidt, H.; de Weerth, C. Gut microbiota and BMI throughout childhood: The role of firmicutes, bacteroidetes, and short-chain fatty acid producers. Sci. Rep. 2022, 12, 3140. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.S.; Comelli, E.M. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr. Diabetes 2014, 4, e121. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Alvarez-Quintero, R.; Velasquez-Mejia, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher fecal Short-Chain fatty acid levels are associated with gut microbiome dysbiosis, obesity, hypertension and cardiometabolic disease risk factors. Nutrients 2019, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Nkosi, B.V.; Padayachee, T.; Gront, D.; Nelson, D.R.; Syed, K. Contrasting health effects of bacteroidetes and firmicutes lies in their genomes: Analysis of p450s, ferredoxins, and secondary metabolite clusters. Int. J. Mol. Sci. 2022, 23, 5057. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Kamareddine, L.; Najjar, H.; Sohail, M.U.; Abdulkader, H.; Al-Asmakh, M. The Microbiota and Gut-Related Disorders: Insights from Animal Models. Cells 2020, 9, 2401. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M.V. Honey bees as a model for vision, perception, and cognition. Annu. Rev. Entomol. 2010, 55, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X.; Zhang, Z.; Lang, H.; Zheng, H. Honey bee as a model organism to study gut microbiota and diseases. Drug Discov. Today Dis. Models 2018, 28, 35–42. [Google Scholar] [CrossRef]
- Zhang, Z.; Mu, X.; Cao, Q.; Shi, Y.; Hu, X.; Zheng, H. Honeybee gut microbiota modulates host behaviors and neuro logical processes. bioRxiv 2020. [CrossRef]
- Zheng, H.; Steele, M.I.; Leonard, S.P.; Motta, E.V.S.; Moran, N.A. Honey bees as models for gut microbiota research. Lab Anim. 2018, 47, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Simple animal models for microbiome research. Nat. Rev. Microbiol. 2019, 17, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Lang, H.; Wang, H.; Wang, H.; Zhong, Z.; Xie, X.; Zhang, W.; Guo, J.; Meng, L.; Hu, X.; Zhang, X.; et al. Engineered symbiotic bacteria interfering Nosema redox system inhibit microsporidia parasitism in honeybees. Nat. Commun. 2023, 14, 2778. [Google Scholar] [CrossRef] [PubMed]
- Ellegaard, K.M.; Tamarit, D.; Javelind, E.; Olofsson, T.C.; Andersson, S.G.; Vásquez, A. Extensive intra-phylotype diversity in lactobacilli and bifidobacteria from the honeybee gut. BMC Genom. 2015, 16, 284. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Arteel, G.E. Effect of ethanol on lipid metabolism. J. Hepatol. 2019, 70, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Suliga, E.; Kozieł, D.; Ciesla, E.; Rebak, D.; Głuszek-Osuch, M.; Naszydłowska, E.; Głuszek, S. The consumption of alcoholic beverages and the prevalence of cardiovascular diseases in men and women: A Cross-Sectional study. Nutrients 2019, 11, 1318. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.; Urpi-Sarda, M.; Martinez-Gonzalez, M.A.; Andres-Lacueva, C.; Mitjavila, M.T. Dealcoholised beers reduce atherosclerosis and expression of adhesion molecules in apoE-deficient mice. Brit. J. Nutr. 2011, 105, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Bassus, S.; Mahnel, R.; Scholz, T.; Wegert, W.; Westrup, D.; Kirchmaier, C.M. Effect of Dealcoholized Beer (Bitburger Drive®) Consumption on Hemostasis in Humans. Alcohol. Clin. Exp. Res. 2004, 28, 786–791. [Google Scholar] [CrossRef] [PubMed]
- DE Curtis, A.; Murzilli, S.; DI Castelnuovo, A.; Rotilio, D.; Donati, M.B.; DE Gaetano, G.; Iacoviello, L. Alcohol-free red wine prevents arterial thrombosis in dietary-induced hypercholesterolemic rats: Experimental support for the ‘French paradox’. J. Thromb. Haemost. 2005, 3, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Arranz, S.; Chiva-Blanch, G.; Valderas-Martínez, P.; Medina-Remón, A.; Lamuela-Raventós, R.M.; Estruch, R. Wine, Beer, Alcohol and Polyphenols on Cardiovascular Disease and Cancer. Nutrients 2012, 4, 759–781. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xu, E.; Long, J.; Pan, X.; Xu, X.; Jin, Z.; Jiao, A. Comparison between ATR-IR, Raman, concatenated ATR-IR and Raman spectroscopy for the determination of total antioxidant capacity and total phenolic content of Chinese rice wine. Food Chem. 2016, 194, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Oladokun, O.; Tarrega, A.; James, S.; Smart, K.; Hort, J.; Cook, D. The impact of hop bitter acid and polyphenol profiles on the perceived bitterness of beer. Food Chem. 2016, 205, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qi, Y.; Zheng, H. Dietary polyphenol, gut microbiota, and health benefits. Antioxidants 2022, 11, 1212. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J.; Yin, N.; Wele, P.; Li, F.; Dave, S.; Lin, J.; Xiao, H.; Wu, X. Resveratrol in disease prevention and health promotion: A role of the gut microbiome. Crit. Rev. Food Sci. 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Fang, L.; Pan, Y.; Jiang, L. Effect of grape seed proanthocyanidins on fat metabolism and adipocytokines in obese rats. Metabolites 2023, 13, 568. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morato, J.; Matthan, N.R.; Liu, J.; de la Torre, R.; Chen, C.Y.O. Cranberries attenuate animal-based diet-induced changes inmicrobiota composition and functionality: A randomized crossover controlled feeding trial. J. Nutr. Biochem. 2018, 62, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhong, Z.; Chen, X.; Hong, Z.; Lin, W.; Mu, X.; Hu, X.; Zheng, H. High-Fat diets with differential fatty acids induce obesity and perturb gut microbiota in honey bee. Int. J. Mol. Sci. 2021, 22, 834. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, Y.; Zhou, Y.; Li, Y.; Zhou, T.; Zheng, J.; Zhang, J.; Li, S.; Xu, D.; Li, H. Effects of beverages on alcohol metabolism: Potential health benefits and harmful impacts. Int. J. Mol. Sci. 2016, 17, 354. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Nishida, A.; Kwong, W.K.; Koch, H.; Engel, P.; Steele, M.I.; Moran, N.A. Metabolism of toxic sugars by strains of the bee gut symbiont Gilliamella apicola. mBio 2016, 7, 01326-16. [Google Scholar] [CrossRef] [PubMed]
- Aires, V.; Labbé, J.; Deckert, V.; Pais De Barros, J.; Boidot, R.; Haumont, M.; Maquart, G.; Le Guern, N.; Masson, D.; Prost-Camus, E.; et al. Healthy adiposity and extended lifespan in obese mice fed a diet supplemented with a polyphenol-rich plant extract. Sci. Rep. 2019, 9, 9134. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhai, Y.; Zhou, W.; Qiao, Y.; Guan, L.; Liu, H.; Jiang, J.; Peng, L. Intestinal flora mediates antiobesity effect of rutin in High-Fat-Diet mice. Mol. Nutr. Food Res. 2022, 66, 2100948. [Google Scholar] [CrossRef] [PubMed]
- Nakadate, K.; Kawakami, K.; Yamazaki, N. Anti-Obesity and Anti-Inflammatory synergistic effects of green tea catechins and citrus β-Cryptoxanthin ingestion in obese mice. Int. J. Mol. Sci. 2023, 24, 7054. [Google Scholar] [CrossRef] [PubMed]
- Kešnerová, L.; Emery, O.; Troilo, M.; Liberti, J.; Erkosar, B.; Engel, P. Gut microbiota structure differs between honeybees in winter and summer. ISME J. 2020, 14, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Moreira Júnior, R.E.; de Carvalho, L.M.; Dos Reis, D.C.; Cassali, G.D.; Faria, A.M.C.; Maioli, T.U.; Brunialti-Godard, A.L. Diet-induced obesity leads to alterations in behavior and gut microbiota composition in mice. J. Nutr. Biochem. 2021, 92, 108622. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Halaka, D.A.; Hayouka, Z.; Tirosh, O. High-Fat Diet Induced Alteration of Mice Microbiota and the Functional Ability to Utilize Fructooligosaccharide for Ethanol Production. Front. Cell. Infect. Microbiol. 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuno, M.I.; Boto-Ordonez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Diaz, F.C.; Andres-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 12. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Kandimalla, R.; Bhattacharya, A.; Wahengbam, R.; Dehingia, M.; Kalita, M.C.; Talukdar, N.C.; Talukdar, R.; Khan, M.R. Multi-Omics analysis demonstrates the critical role of Non-Ethanolic components of alcoholic beverages in the host microbiome and metabolome: A human- and Animal-Based study. Microorganisms 2023, 11, 1501. [Google Scholar] [CrossRef] [PubMed]
- Zorraquín, I.; Sánchez-Hernández, E.; Ayuda-Durán, B.; Silva, M.; González-Paramás, A.M.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. Current and future experimental approaches in the study of grape and wine polyphenols interacting gut microbiota. J. Sci. Food Agric. 2020, 100, 3789–3802. [Google Scholar] [CrossRef] [PubMed]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, 73, 24. [Google Scholar] [CrossRef]
- Hybl, M.; Mraz, P.; Sipos, J.; Hostickova, I.; Bohata, A.; Curn, V.; Kopec, T. Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis mellifera) Intoxicated by Pesticide Thiacloprid. Insects 2021, 12, 572. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xu, J.; Sheng, Y.; Liu, J.; Li, H.; Guo, M.; Xu, W.; Luo, Y.; Huang, K.; He, X. Pleurotus ostreatus ameliorates obesity by modulating the gut microbiota in obese mice induced by High-Fat diet. Nutrients 2022, 14, 1868. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Dai, K.; Song, Y.; Zhang, D.; Wei, Y.; Jiang, S.; Xu, F.; Wang, H.; Zhang, X.; Shao, X. Thinned peach polyphenols alleviate obesity in high fat mice by affecting gut microbiota. Food Res. Int. 2022, 157, 111255. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | W | WD | B | BD | HJ | HJD | BJ | BJD |
|---|---|---|---|---|---|---|---|---|
| Gallic acid | 273.0 ± 0.9 | 301.0 ± 0.2 | — | — | — | — | — | — |
| Caffeic acid | 5.0 ± 0.2 | 6 ± 0.4 | — | — | — | — | — | — |
| Catechin | 72.0. ± 0.8 | 63 ± 0.7 | — | — | — | — | — | — |
| L-Epicatechin | 6.0 ± 0.1 | 5.0 ± 0.1 | — | — | — | — | — | — |
| Quercetin | 95.0 ± 0.7 | 7.0 ± 0.2 | — | — | — | — | — | — |
| Chlorogenic acid | 0.6 ± 0.1 | 1.0 ± 0.1 | — | — | — | — | — | — |
| (-)-Gallocatechin | — | — | 10.0 ± 0.2 | 1.0 ± 0.1 | — | — | ||
| Rutin | 46.0 ± 0.2 | 64.0 ± 0.9 | 33.0 ± 0.3 | 27.0 ± 0.7 | — | — | — | — |
| Vanillic acid | 9.0 ± 0.7 | 10.5 ± 0.7 | — | — | — | — | — | — |
| Resveratrol | 24.0 ± 0.2 | 30.0 ± 0.2 | — | — | — | — | — | — |
| Kaempferol | 1.0 ± 0.1 | 0.40 ± 0.01 | — | — | — | — | — | — |
| Ferulic acid | 2.0 ± 0.1 | 2.0 ± 0.1 | 73.0 ± 0.3 | 63.0 ± 0.5 | 47.0 ± 0.5 | 36.0 ± 0.9 | — | — |
| Para-hydroxybenzoic acid | 0.60 ± 0.03 | 0.60 ± 0.09 | — | — | — | — | — | — |
| Erucic acid | 4.0 ± 0.1 | 1.0 ± 0.1 | 10.0 ± 0.2 | 9.0 ± 0.3 | 0.50 ± 0.02 | 0.50 ± 0.02 | — | — |
| 3,4-Dihydroxybenzoic acid | 22 ± 0.34 | 24 ± 0.39 | 0.1 ± 0.03 | 0.1 ± 0.03 | — | — | — | — |
| Total monomeric phenolics | 476.0 ± 0.8 | 518.0 ± 0.9 | 117.0 ± 0.4 | 99.0 ± 0.7 | 48.0 ± 0.2 | 37.0 ± 0.8 | — | — |
| Mannose | 280.0 ± 0.4 | 301.0 ± 0.3 | 9.0 ± 0.6 | 74.0 ± 0.1 | 8.0 ± 0.7 | 301.0 ± 0.1 | — | — |
| Ribose | 82.0 ± 0.4 | 79.0 ± 0.5 | — | 172.0 ± 0.1 | — | 41.0 ± 0.3 | — | — |
| Rhamnose | 243.0 ± 0.4 | 241.0 ± 0.1 | — | — | — | — | — | — |
| Glucuronic acid | 44.0 ± 0.1 | 48.0 ± 0.7 | 399.0 ± 0.3 | 506.0 ± 0.1 | 361.0 ± 0.4 | 560.0 ± 0.1 | — | — |
| Galacturonic acid | 406.0. ± 0.7 | 410.0 ± 0.5 | 8 ± 0.1 | — | — | — | — | — |
| Glucose | 1000.0 ± 0.9 | 1015.0 ± 0.6 | 20,847.0 ± 0.1 | 27,026.0 ± 0.1 | 18,784.0 ± 0.3 | 34,746.0 ± 0.4 | 3.0 ± 0.2 | 6.0 ± 0.1 |
| Galactose | 214.0 ± 0.1 | 219.0 ± 0.7 | — | 40.0 ± 0.4 | — | 100.0 ± 0.6 | — | — |
| Xylose | 135.0 ± 0.7 | 141.0 ± 0.3 | — | 233.0 ± 0.1 | — | 284.0 ± 0.1 | 1.0 ± 0.4 | 1.0. ± 0.5 |
| Arabinose | 214.0 ± 0.1 | 220.0 ± 0.3 | — | 140.0 ± 0.4 | — | 180.0 ± 0.3 | — | — |
| Fucose | 25.0 ± 0.2 | 39.0 ± 0.1 | — | — | — | 166.0 ± 0.5 | — | — |
| Total monosaccharides | 2646.0 ± 0.9 | 2717.0 ± 0.7 | 21,263.0 ± 0.2 | 28,192.0 ± 0.6 | 19,155.0 ± 0.8 | 36,381.0 ± 0.4 | 4.0 ± 0.1 | 7.0 ± 0.5 |
| Aspartic acid | 124.0 ± 0.1 | 52.0 ± 0.3 | 279.0 ± 0.6 | 290.0 ± 0.7 | 132.0 ± 0.9 | 54.0 ± 0.3 | — | — |
| Glutamic acid | 444.0 ± 0.6 | 48.0 ± 0.8 | 1015.0 ± 0.9 | 1050.0 ± 0.7 | 453.0 ± 0.5 | 56.0 ± 0.5 | — | — |
| Hydroxyproline | 4.0 ± 0.1 | 11.0 ± 0.2 | 6.0 ± 0.3 | 6.0 ± 0.3 | 4.0 ± 0.1 | 11.0 ± 0.2 | — | — |
| Serine | 72.0. ± 0.5 | 38.0 ± 0.2 | 265.0 ± 0.4 | 264.0 ± 0.3 | 74.0 ± 0.1 | 36.0 ± 0.2 | — | — |
| Glycine | 67.0 ± 0.1 | 31.0 ± 0.2 | 293.0 ± 0.9 | 293.0. ± 0.9 | 71.0 ± 0.1 | 32.0 ± 0.1 | — | — |
| Histidine | 29.0 ± 0.2 | 3.0 ± 0.1 | 104.0. ± 0.2 | 102.0 ± 0.2 | 30.0 ± 0.3 | 3.0 ± 0.1 | — | — |
| Arginine | 50.0 ± 0.6 | 4.0 ± 0.1 | 252.0 ± 0.4 | 250.0 ± 0.3 | 52.0 ± 0.3 | 4.0 ± 0.5 | — | — |
| Threonine | 34.0. ± 0.3 | 16.0 ± 0.2 | 159.0 ± 0.2 | 164.0 ± 0.4 | 36.0 ± 0.3 | 19.0 ± 0.3 | — | — |
| Alanine | 116.0 ± 0.4 | 39.0. ± 0.3 | 362.0 ± 0.4 | 380.0 ± 0.5 | 124.0 ± 0.3 | 44.0 ± 0.3 | — | — |
| Proline | 252.0 ± 0.4 | 804.0 ± 0.8 | 665.0 ± 0.7 | 676.0 ± 0.7 | 276.0 ± 0.4 | 849.0 ± 0.6 | — | — |
| Tyrosine | 50.0 ± 0.4 | 7.0. ± 0.3 | 187.0 ± 0.5 | 191.0 ± 0.8 | 53.0 ± 0.6 | 8.0 ± 0.1 | ||
| Valine | 67.0 ± 0.4 | 20.0 ± 0.3 | 227.0 ± 0.7 | 232.0 ± 0.6 | 68.0 ± 0.5 | 23.0 ± 0.4 | ||
| Methionine | 8.0 ± 0.1 | 2.0 ± 0.1 | 16.0 ± 0.2 | 210 ± 0.2 | 11.0 ± 0.3 | 2.0 ± 0.3 | ||
| Cysteine | 4.0 ± 0.1 | 13.0 ± 0.2 | 13.0 ± 0.2 | 60.0. ± 0.2 | 5.0 ± 0.2 | 10.0 ± 0.2 | ||
| Isoleucine | 35.0 ± 0.3 | 18.0 ± 0.2 | 169.0 ± 0.4 | 177.0 ± 0.3 | 35.0 ± 0.1 | 20.0 ± 0.4 | ||
| Leucine | 64.0. ± 0.5 | 32.0 ± 0.3 | 310.0 ± 0.6 | 324.0 ± 0.7 | 67.0 ± 0.3 | 49.0 ± 0.5 | ||
| Ortholeucine | — | — | — | — | — | — | ||
| Phenylalanine | 52.0 ± 0.3 | 18.0 ± 0.3 | 251.0 ± 0.7 | 249.0 ± 0.5 | 55.0 ± 0.4 | 19.0 ± 0.3 | ||
| Lysine | 44.0 ± 0.5 | 18.0. ± 0.5 | 108.0 ± 0.7 | 119.0 ± 0.4 | 46.0 ± 0.4 | 22.0. ± 0.3 | — | — |
| Total amino acids | 1518.0. ± 0.9 | 1174.0 ± 0.8 | 4680.0 ± 0.4 | 4793.0 ± 0.4 | 1591.0 ± 0.5 | 1261.0 ± 0.4 | — | — |
| Oxalic acid | 31.0 ± 0.9 | 32.0 ± 0.5 | 3.0 ± 0.3 | 3.0 ± 0.1 | 32.0 ± 0.5 | 31.0 ± 0.3 | 0.40 ± 0.03 | 0.40 ± 0.07 |
| Tartaric acid | 1255.0 ± 0.7 | 1687.0. ± 0.8 | — | — | 29.0 ± 0.7 | 27.0. ± 0.1 | — | — |
| Malic acid | — | — | 58.0 ± 0.5 | 53.0 ± 0.3 | 14.0 ± 0.1 | 9.0 ± 0.01 | — | — |
| Lactic acid | 1719.0 ± 0.05 | 2233.0 ± 0.6 | — | 133.0 ± 0.1 | 156.0 ± 0.9 | 873.0. ± 0.3 | 4763.0 ± 0.4 | 1728.0 ± 0.4 |
| Acetic acid | 156.0 ± 0.5 | — | 2962.0 ± 0.5 | — | — | 58.0 ± 0.6 | — | — |
| Maleic acid | 0.20 ± 0.06 | 0.10 ± 0.07 | 10.0 ± 0.3 | 9.0 ± 0.5 | 24.0 ± 0.5 | 10.0 ± 0.1 | — | 0.30 ± 0.06 |
| Citric acid | 64.0 ± 0.6 | 286.0 ± 0.4 | 1460.0 ± 0.7 | 1311.0 ± 0.7 | 617.0 ± 0.1 | 3857.0 ± 0.4 | 5.0 ± 0.6 | 38.0 ± 0.8 |
| Fumaric acid | — | — | 1.80 ± 0.07 | 1.6 ± 0.4 | 0.40 ± 0.05 | — | — | 0.20 ± 0.02 |
| Succinic acid | 22.0 ± 0.7 | 3363.0 ± 0.1 | 12.0. ± 0.1 | 12.0 ± 0.5 | 2491.0. ± 0.7 | 18.0 ± 0.1 | 4.7 ± 0.1 | 91.0 ± 0.6 |
| Total organic acids | 3550.0 ± 0.7 | 7566.0 ± 0.2 | 1703.0 ± 0.02 | 1524.0 ± 0.5 | 5141.0 ± 0.6 | 8716.0 ± 0.5 | 4773.0 ± 0.2 | 1859.0 ± 0.3 |
| Composition | W | WD | WDX5 | B | BD | HJ | HJD | BJ | BJD | SUC | Alcohol | PO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gallic acid | 44.0 ± 0.7 | 420 ± 0.2 | 9.0 ± 0.1 | — | — | — | — | — | — | — | — | — |
| Caffeic acid | 0.80 ± 0.04 | 0.90 ± 0.01 | 0.20 ± 0.02 | — | — | — | — | — | — | — | — | — |
| Catechin | 12.0 ± 0.3 | 9.0 ± 0.1 | 2.0 ± 0.1 | — | — | — | — | — | — | — | — | — |
| Epicatechin | 0.90 ± 0.09 | 0.70 ± 0.03 | 0.10 ± 0.04 | — | — | — | — | — | — | — | — | — |
| Quercetin | 1.60 ± 0.02 | 1.0 ± 0.1 | 0.20 ± 0.03 | — | — | — | — | — | — | — | — | — |
| Chlorogenic acid | — | — | — | 0.40 ± 0.08 | 0.40 ± 0.01 | — | — | — | — | — | — | — |
| Rutin | 8.0 ± 0.3 | 9.0 ± 0.1 | 2.0 ± 0.2 | 13.0 ± 0.3 | 10.0 ± 0.2 | — | — | — | — | — | — | — |
| Vanillic acid | 1.5 ± 0.2 | 1.5 ± 0.1 | 0.30 ± 0.02 | — | — | — | — | — | — | — | — | — |
| Resveratrol | 4.0. ± 0.1 | 4.0 ± 0.8 | 0.80 ± 0.01 | — | — | — | — | — | — | — | — | — |
| Kaempferol | 0.20 ± 0.01 | 0.10 ± 0.01 | — | — | — | — | — | — | — | — | — | — |
| Ferulic acid | 0.30 ± 0.04 | 0.20 ± 0.04 | — | 28.0 ± 0.9 | 24.0 ± 0.4 | 3.8 ± 0.3 | 2.8 ± 0.2 | — | — | — | — | — |
| Para-hydroxybenzoic acid | 0.10 ± 0.03 | — | — | — | — | — | — | — | — | — | — | — |
| Erucic acid | 0.60 ± 0.04 | 0.20 ± 0.01 | - | 3.7 ± 0.8 | 3.3 ± 0.1 | — | — | — | — | — | — | — |
| 3,4-Dihydroxybenzoic acid | 3.7 ± 0.2 | 3.3 ± 0.8 | 0.60 ± 0.08 | — | — | — | — | — | — | — | — | — |
| Total monomeric phenolics | 78.0 ± 0.7 | 72.0 ± 0.9 | 15.0 ± 0.4 | 45.0 ± 0.6 | 38.0 ± 0.7 | 4.0 ± 0.6 | 3.0 ± 0.2 | — | — | — | — | — |
| Mannose | 10.7 ± 0.2 | 11.7 ± 0.1 | 11.6 ± 0.3 | 28.3 ± 0.6 | 21.0 ± 0.5 | 12.4 ± 0.2 | 15.0 ± 0.7 | 12.4 ± 0.4 | 11.0 ± 0.2 | 6140.2 ± 0.4 | 1313.9 ± 0.2 | 6080.2 ± 0.1 |
| Ribose | 61.4 ± 0.1 | 47.7 ± 0.4 | 40.3 ± 0.9 | 98.2 ± 0.8 | 64.8 ± 0.6 | 30.1 ± 0.5 | 42.4 ± 0.3 | — | — | 799.9 ± 0.6 | 369.0 ± 0.7 | 1247.6 ± 0.7 |
| rhamnose | — | — | — | — | — | — | — | — | — | 2498.4 ± 0.8 | 241.2 ± 0.9 | 10,148.0 ± 0.8 |
| Glucuronic acid | 123.1 ± 0.5 | 86.9 ± 0.4 | 85.0 ± 0.9 | 275.9 ± 0.6 | 161.0 ± 0.8 | 66.0 ± 0.4 | 88.3 ± 0.2 | 66.0 ± 0.1 | 143.8 ± 0.6 | 1186.6 ± 0.4 | 452.5 ± 0.8 | 4374.0 ± 0.3 |
| Galacturonic acid | — | — | — | 110.7 ± 0.65 | — | — | 41.5 ± 0.5 | — | — | 800.7 ± 0.7 | 135.7 ± 0.4 | 4128.7 ± 0.8 |
| Glucose | 6126.4 ± 0.7 | 5335.3 ± 0.8 | 4992.4 ± 0.8 | 13,067.8 ± 0.9 | 10,665.7 ± 0.8 | 4605.5 ± 0.8 | 6281.8 ± 0.6 | 4605.5 ± 0.6 | 7080.2 ± 0.1 | 19,855.6 ± 0.1 | 17,016.4 ± 0.1 | 21,137.7 ± 0.1 |
| Galactose | — | 6.9 ± 0.1 | 5.7 ± 0.1 | — | — | — | — | — | — | 8929.1 ± 0.8 | 2109.5 ± 0.7 | 47,749.9 ± 0.9 |
| Xylose | — | 7.0 ± 0.5 | 0.80 ± 0.15 | 4.8 ± 0.1 | 7.7 ± 0.1 | 14.7 ± 0.1 | — | 14.7 ± 0.4 | — | 228.6 ± 0.5 | 22.6 ± 0.1 | 355.8 ± 0.9 |
| Arabinose | 3.2 ± 0.1 | — | — | 10.7 ± 0.7 | 11.7 ± 0.5 | 5.1 ± 0.1 | — | 5.1 ± 0.7 | — | 5877.5 ± 0.5 | 1512.5 ± 0.1 | 30,350.2 ± 0.9 |
| Fucose | — | — | — | 11.3 ± 0.1 | — | — | — | — | — | 847.5 ± 0.1 | 302.8 ± 0.5 | 1477.4 ± 0.7 |
| Total monosaccharides | 6324.9 ± 0.1 | 5495.6 ± 0.9 | 5135.9 ± 0.4 | 13,607.6 ± 0.7 | 10,931.8 ± 0.1 | 4733.8 ± 0.5 | 6469.0 ± 0.3 | 4703.7 ± 0.6 | 7235.0 ± 0.3 | 47,164.1 ± 0.7 | 23,476.1 ± 0.9 | 12,7049.6 ± 0.5 |
| Aspartic acid | 5.0 ± 0.2 | 4.0 ± 0.1 | 4.0 ± 0.1 | 5.0 ± 0.3 | 4.0 ± 0.4 | — | 4.0 ± 0.5 | — | — | — | — | — |
| Glutamic acid | — | — | — | — | — | — | — | — | — | — | — | — |
| Hydroxyproline | 2.0 ± 0.3 | 1.0. ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 2.0 ± 02 | — | — | — | — | — |
| Serine | — | — | 1.0 ± 0.1 | — | 1.0 ± 0.1 | 1.0 ± 0.1 | — | — | — | — | — | — |
| Glycine | 2.0 ± 0.4 | 2.0 ± 0.3 | 2.0 ± 0.2 | 2.0 ± 0.1 | 2.0 ± 0.05 | 2.0 ± 0.3 | 2.0 ± 0.6 | — | — | — | — | — |
| Histidine | — | 1.0. ± 0.4 | — | 1.0 ± 0.1 | 1.0 ± 0.2 | — | — | — | — | — | — | — |
| Arginine | — | — | — | — | — | — | — | — | — | — | — | — |
| Threonine | 7.0 ± 0.4 | 4.0 ± 0.3 | 5.0 ± 0.8 | 4.0 ± 0.2 | 5.0 ± 0.1 | 3.0. ± 0.2 | 5.0 ± 0.3 | — | — | — | — | — |
| Alanine | — | 1.0. ± 0.1 | — | 1.0 ± 0.2 | 1.0 ± 0.2 | — | 1.0 ± 0.1 | — | — | — | — | — |
| Proline | — | — | — | 1.0 ± 0.08 | 1.0 ± 0.11 | 1.0 ± 0.05 | — | — | — | — | — | — |
| Lysine | 1.0 ± 0.1 | 1.0 ± 0.2 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | — | — | — | — | — |
| Total amino acids | 17.0 ± 0.2 | 14.0 ± 0.5 | 13.0 ± 0.8 | 16.0 ± 0.5 | 16.0 ± 0.5 | 9.0 ± 0.5 | 15.0 ± 0.6 | — | — | — | — | — |
| Oxalic acid | 1.0 ± 0.1 | 0.50 ± 0.01 | 0.2 ± 0.03 | 1.40 ± 0.05 | 0.80 ± 0.08 | 3.30 ± 0.06 | 2.90 ± 0.01 | 0.10 ± 0.01 | — | — | — | — |
| Tartaric acid | 109.0 ± 0.8 | 116.0 ± 0.8 | 24.0 ± 0.1 | 53.0 ± 0.1 | 34.0 ± 0.4 | 5.0 ± 0.3 | 4.0 ± 0.6 | — | — | — | — | — |
| Malic acid | — | — | — | 11.0 ± 0.5 | 3.0 ± 0.3 | 5.0 ± 0.4 | 4.0 ± 0.6 | — | — | — | — | — |
| Lactic acid | 244.0 ± 0.1 | 188.0 ± 0.4 | 53.0 ± 0.4 | 38.0 ± 0.6 | 57.0 ± 0.4 | 347.0 ± 0.9 | 285.0 ± 0.3 | 47.0 ± 0.6 | 34.0 ± 0.5 | — | — | — |
| Acetic acid | 272.0 ± 0.8 | 201.0 ± 0.6 | 45.0 ± 0.9 | 21.0 ± 0.8 | 71.0. ± 0.8 | 64.0 ± 0.2 | 6.0 ± 0.4 | 35.0 ± 0.2 | 20.0 ± 0.6 | — | — | — |
| Maleic acid | — | — | — | — | — | — | — | ±0.01 | ±0.01 | — | — | — |
| Citric acid | 56.0 ± 0.9 | 44.0 ± 0.8 | 19.0 ± 0.2 | 436.0 ± 0.3 | 345.0 ± 0.1 | 178.0 ± 0.1 | 157.0 ± 0.6 | 12.0 ± 0.8 | 7.0 ± 0.8 | — | — | — |
| Fumaric acid | — | — | — | 0.40 ± 0.06 | 0.30 ± 0.02 | 0.30 ± 0.06 | 0.30 ± 0.02 | — | — | — | — | — |
| Succinic acid | 49.0 ± 0.6 | 41.0 ± 0.6 | 8.0 ± 0.9 | 12.0 ± 0.2 | 23.0 ± 0.5 | 22.0 ± 0.5 | 20.0 ± 0.2 | 2.0 ± 0.4 | 1.0 ± 0.3 | — | — | — |
| Total organic acids | 734.0 ± 0.5 | 594.0. ± 0.9 | 152.0 ± 0.2 | 575.0 ± 0.1 | 537.0 ± 0.5 | 626.0 ± 0.8 | 482.0 ± 0.4 | 97.0 ± 0.6 | 64.0 ± 0.4 | — | — | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, G.; Wang, X.; Gao, C.; Kang, X.; Xue, H.; Huang, W.; Zhan, J.; You, Y. Effects of Active Ingredients in Alcoholic Beverages and Their De-Alcoholized Counterparts on High-Fat Diet Bees: A Comparative Study. Molecules 2024, 29, 1693. https://doi.org/10.3390/molecules29081693
Fan G, Wang X, Gao C, Kang X, Xue H, Huang W, Zhan J, You Y. Effects of Active Ingredients in Alcoholic Beverages and Their De-Alcoholized Counterparts on High-Fat Diet Bees: A Comparative Study. Molecules. 2024; 29(8):1693. https://doi.org/10.3390/molecules29081693
Chicago/Turabian StyleFan, Guanghe, Xiaofei Wang, Cuicui Gao, Xiping Kang, Huimin Xue, Weidong Huang, Jicheng Zhan, and Yilin You. 2024. "Effects of Active Ingredients in Alcoholic Beverages and Their De-Alcoholized Counterparts on High-Fat Diet Bees: A Comparative Study" Molecules 29, no. 8: 1693. https://doi.org/10.3390/molecules29081693
APA StyleFan, G., Wang, X., Gao, C., Kang, X., Xue, H., Huang, W., Zhan, J., & You, Y. (2024). Effects of Active Ingredients in Alcoholic Beverages and Their De-Alcoholized Counterparts on High-Fat Diet Bees: A Comparative Study. Molecules, 29(8), 1693. https://doi.org/10.3390/molecules29081693