A Germin-Like Protein Gene (CchGLP) of Capsicum chinense Jacq. Is Induced during Incompatible Interactions and Displays Mn-Superoxide Dismutase Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
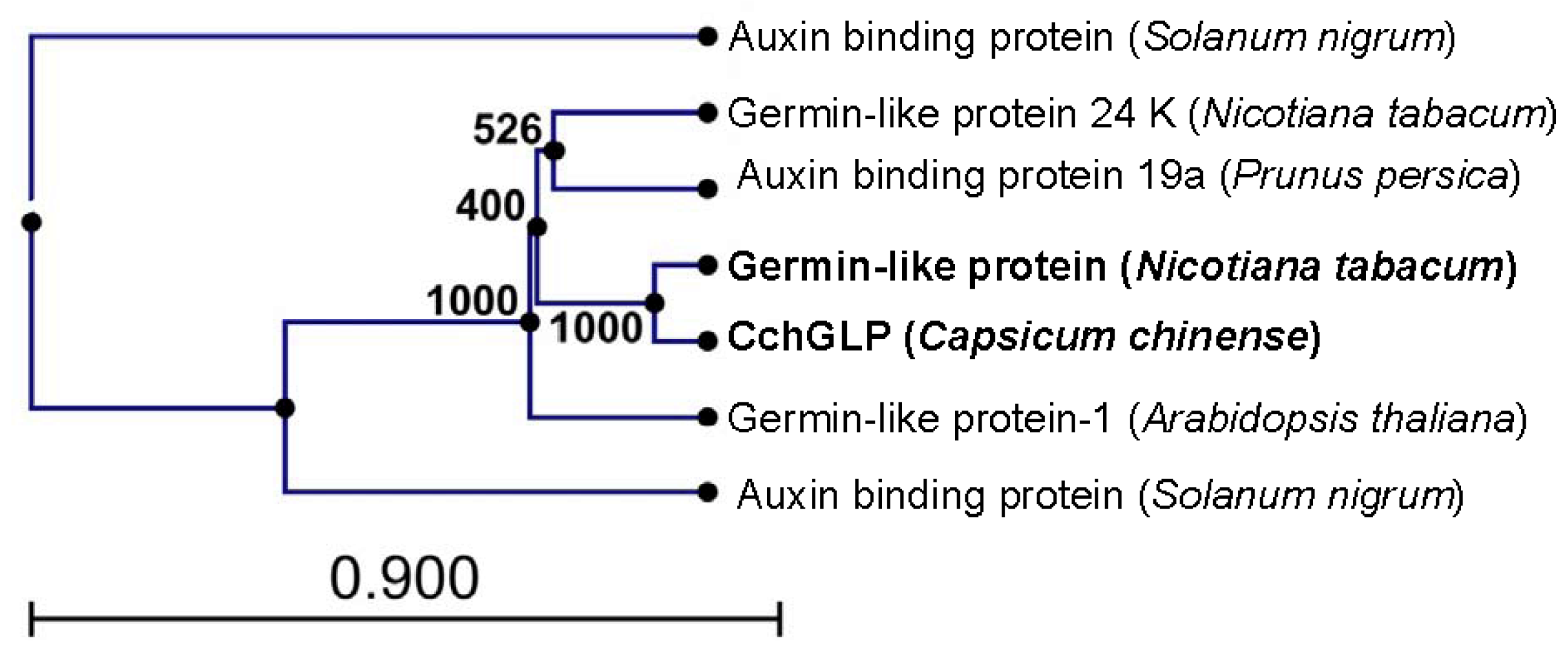
2.1. Characterization of the CchGLP Sequence
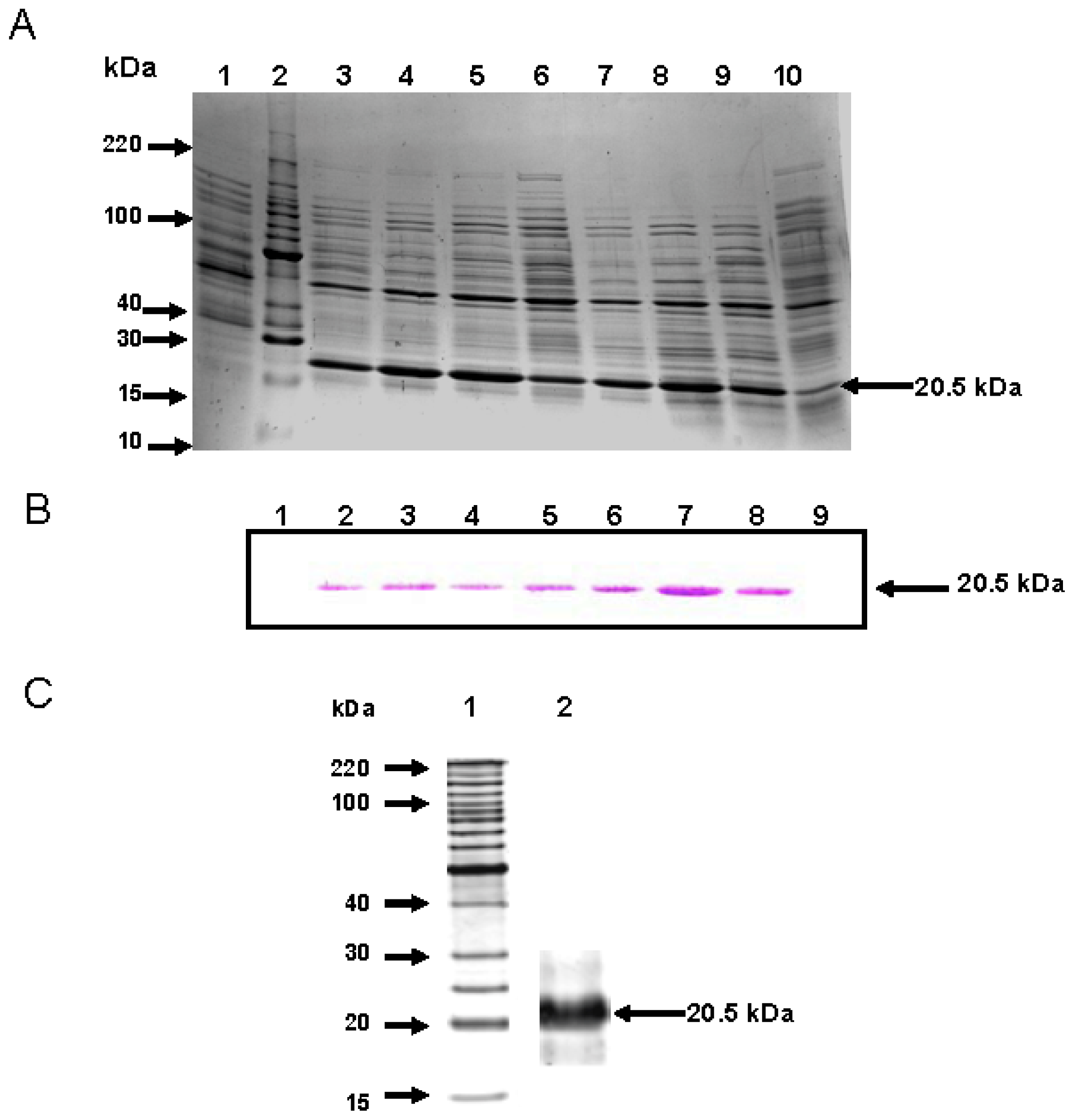
2.2. CchGLP Protein Purification
2.3. In Vitro Enzymatic Activity of CchGLP
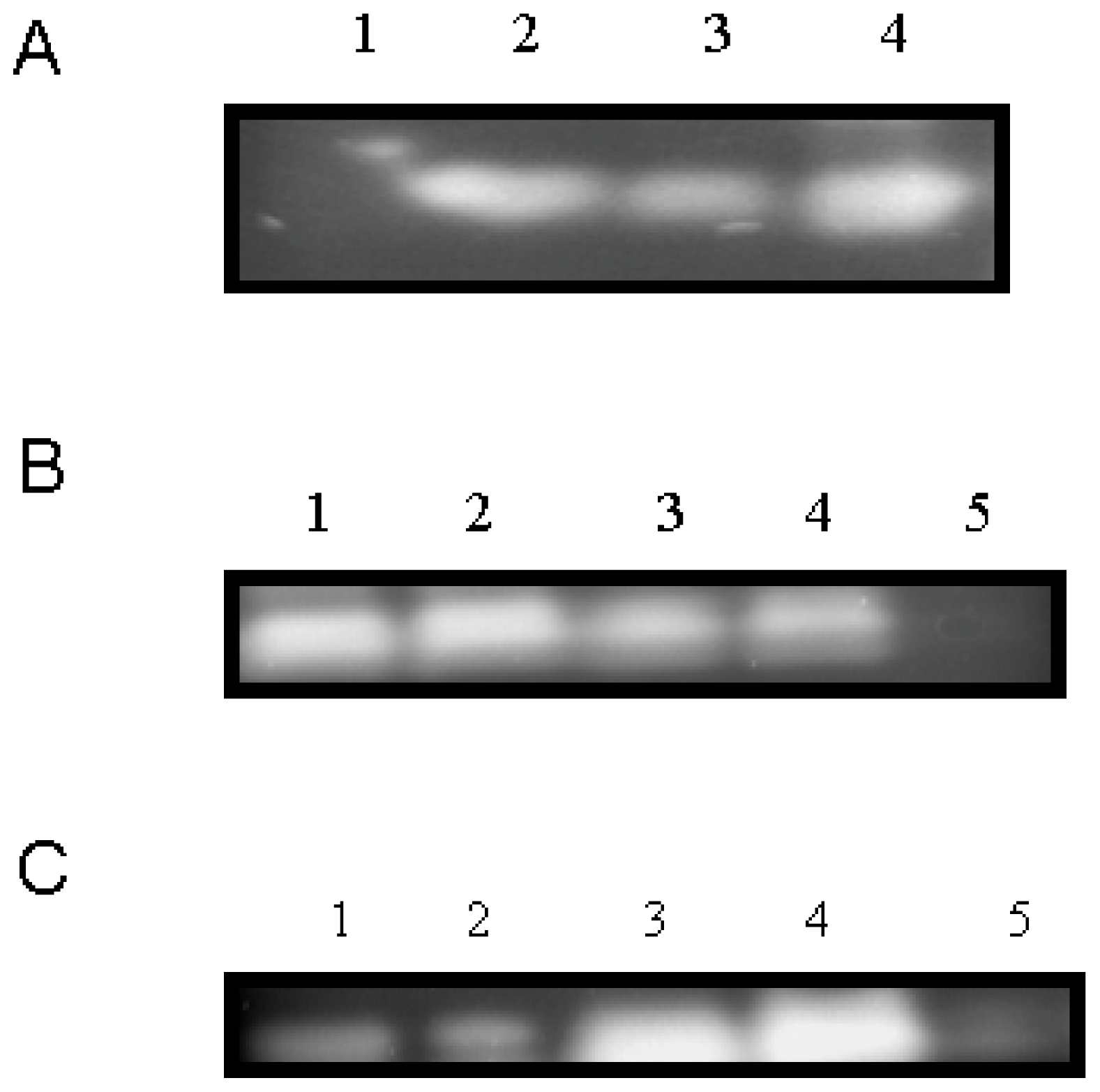
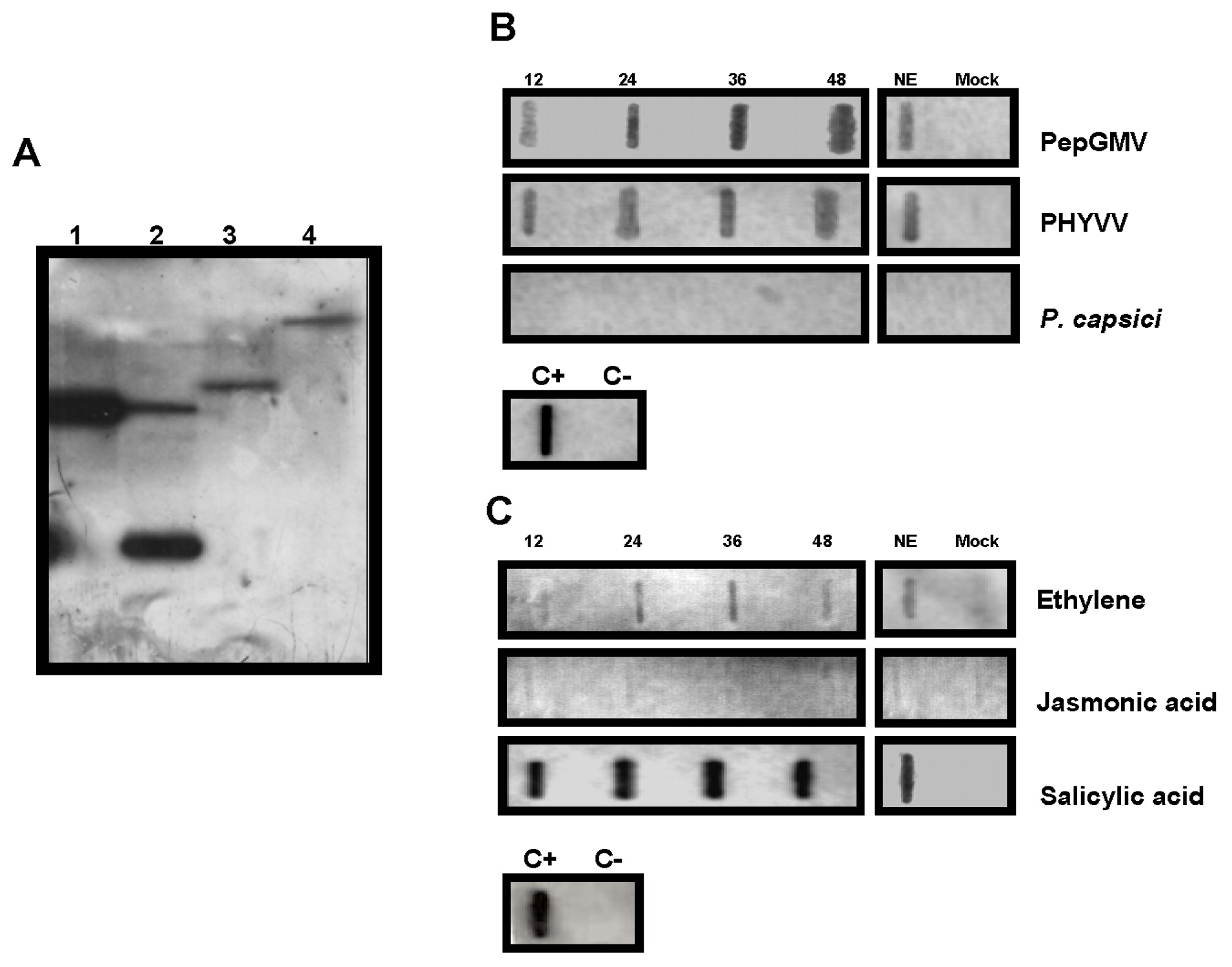
2.4. Molecular Characterization and Gene Expression of CchGLP during Plant-Microbe Interactions
3. Experimental Section
3.1. CchGLP Cloning for Protein Expression in E. coli
3.2. CchGLP Production in E. coli
3.3. Determination of Enzymatic Activity
3.4. Virus and Oomycete Inoculation and Plant Growth Regulators Applications
3.5. Hybridization Analysis
3.6. Analysis of Genomic Sequence of CchGLP Structural Region
3.7. Bioinformatic Analysis
4. Conclusions
Acknowledgments
References
- Schweizer, P.; Christoffel, A.; Dudler, R. Transient expression of members of the germin-like gene family in epidermal cells of wheat confers disease resistance. Plant J 1999, 20, 541–552. [Google Scholar]
- Park, C.-J.; An, J.-M.; Shin, Y.-Ch.; Kim, K.-J.; Lee, B.-J.; Paek, K.-H. Molecular characterization of pepper germin-like protein as the novel PR-16 family of pathogenesis-related proteins isolated during the resistance response to viral and bacterial infection. Planta 2004, 219, 797–806. [Google Scholar]
- Gucciardo, S.; Wisniewski, J.-P.; Brewin, N.; Bornemann, S. A germin-like protein with superoxide dismutase activity in pea nodules with high protein sequence identity to a putative rhicadhesin receptor. J. Exp. Bot 2007, 58, 1161–1171. [Google Scholar]
- Dunwell, J.M.; Khuri, S.; Gane, P. Microbial relatives of the seed storage proteins of higher plants: Conservation of structure and diversification of function during evolution of the cupin superfamily. Microbiol. Mol. Biol. Rev 2000, 64, 153–179. [Google Scholar]
- Carter, C.; Thornburg, R. Tobacco nectarin I-purification and characterization as a germin-like, manganese superoxide dismutase implicated in the defense of floral reproductive tissues. J. Biol. Chem 2000, 275, 36726–36733. [Google Scholar]
- Zimmerman, G.; Bäumlein, H.; Mock, H.P.; Himmelbach, A.; Schweizer, P. The multigene family encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant Physiol 2006, 142, 181–192. [Google Scholar]
- Dunwell, J.; Gibbings, J.G.; Mahmood, T.; Saqlan Naqvi, S.M. Germin and Germin-like Proteins: Evolution, Structure, and Function. Crit. Rev. Plant Sci 2008, 27, 342–375. [Google Scholar]
- Davidson, R.; Reeves, P.A.; Manosalva, P.; Leach, J.E. Germins: A diverse protein family important for crop improvement. Plant Sci 2009, 177, 499–510. [Google Scholar]
- Nakata, M.; Watanabe, Y.; Sakurai, Y.; Hashimoto, Y.; Matsuzaki, M.; Takahashi, Y.; Satoh, T. Germin-like protein gene family of a moss, Physcomitrella patents, phylogenetically falls into two characteristic new clades. Plant Mol. Biol 2004, 56, 381–395. [Google Scholar]
- Rodríguez-López, M.; Baroja-Fernandez, E.; Zandueta-Criado, A.; Moreno-Bruna, B.; Munoz, F.J.; Akazawa, T.; Pozueta-Romero, J. Two isoforms of a nucleotide-sugar pyrophosphatase/phosphodiesterase from barley leaves (Hordeum vulgare L.) are distinct oligomers of HvGLP1, a germin-like protein. FEBS Lett 2001, 490, 44–48. [Google Scholar]
- Barrera-Pacheco, A.; Joaquín-Ramos, A.; Torres-Pacheco, I.; Gonzalez-Chavira, M.; Perez-Perez, C.; Guevara-Olvera, L.; Guevara-Gonzalez, R.G. Analysis of transcriptional expression induced in Capsicum chinense BG-3821 under conditions of biotic and abiotic stress. Agrociencia 2008, 42, 95–106. [Google Scholar]
- Anaya-Lopez, J.L.; Perez-Mora, E.; Torres-Pacheco, I.; Muñoz-Sanchez, C.I.; Guevara-Olvera, L.; Gonzalez-Chavira, M.; Ochoa-Alejo, N.; Rivera-Bustamante, R.F.; Guevara-Gonzalez, R.G. Inducible gene expression by pepper huasteco virus in Capsicum chinense plants with resistance to geminivirus infections. Can. J. Plant Pathol 2005, 27, 276–282. [Google Scholar]
- van Loon, L.C.; Rep, M.; Peterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol 2006, 44, 135–162. [Google Scholar]
- Lu, M.; Han, Y.-P.; Gao, J.-G.; Wang, X.-J.; Li, W.-B. Identification and análisis of the germin-like gene family in soybean. BMC Genomics 2010, 11, 620–635. [Google Scholar]
- Pan, H.Y.; Whittaker, M.M.; Bouveret, R.; Berna, A.; Bernier, F.; Whittaker, J.W. Characterization of wheat germin (oxalate oxidase) expressed by Pichia pastoris. Biochem. Biophys. Res. Commun 2007, 356, 925–929. [Google Scholar]
- Cassland, P.; Larsson, S.; Nilverbrant, N.O.; Jönsson, L.J. Heterologous expression of barley and wheat oxalate oxidase in an E. coli txB gor double mutant. J. Biotechnol 2004, 109, 53–62. [Google Scholar]
- Woo, E.-J.; Dunwell, J.M.; Goodenough, P.W.; Marvier, A.C.; Pickersgill, R.W. Germin is a manganese containing homohexamer with oxalate oxidase and superoxide dismutase activities. Nat. Struct. Biol 2000, 7, 1036–1040. [Google Scholar]
- Manosalva, P.M.; Davidson, R.M.; Liu, B.; Zhu, X.; Hulbert, S.H.; Leung, H.; Leach, J.E. A germin-like protein gene family functions as a complex quantitative trait locus conferring broad-spectrum disease resistance in rice. Plant Physiol 2009, 149, 286–296. [Google Scholar]
- Cheng, H.-Y.; Song, S.-Q. Species and organ diversity in the effects of hydrogen peroxide on superoxide dismutase activity in vitro. J. Integr. Plant Biol 2006, 48, 672–678. [Google Scholar]
- Federico, M.; Iniguez-Luy, F.; Skadsen, R.; Kaeppler, H. Spatial and temporal divergence of expression in duplicated barley germin-like protein-encoding genes. Genetics 2006, 174, 179–190. [Google Scholar]
- Lou, Y.; Baldwin, I.T. Silencing of a germin-like gene in Nicotiana attenuata improves performance of native herbivores. Plant Physiol 2006, 140, 1126–1136. [Google Scholar]
- Godfrey, D.; Able, A.; Dry, I. Induction of a grapevine germin-like protein (VvGLP3) gene is closely linked to the site of Erysiphe necator infection: A possible role in defense? Mol. Plant-Microbe Interact 2007, 20, 1112–1125. [Google Scholar]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol 2002, 5, 325–331. [Google Scholar]
- Mewis, I.; Appel, H.M.; Hom, A.; Raina, R.; Schultz, J.C. Major signaling pathways modulate Arabidopsis glucosinolate accumulation and response to both phloem-feeding and chewing insects. Plant Physiol 2005, 138, 1149–1162. [Google Scholar]
- Burns, S.M.; Hull, S.I. Comparison of serum resistance by defined lipopolysaccharide mutants and an acapsular mutant of uropathogenic Escherichia coli O75:K5. Infect. Immun 1998, 66, 4244–4253. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; pp. 122–135. [Google Scholar]
- Pantoja-Hernandez, M.A.; Muñoz-Sanchez, C.I.; Guevara-Gonzalez, R.G.; Botello-Alvarez, E.; Gonzalez-Chavira, M.; Torres-Pacheco, I.; Guevara-Olvera, L. Expression of ornithine decarboxylase of Coccidioides immitis in three Escherichia coli strains carrying the lamdba DE3 lysogen and an E. coli EWH319 strain odc− null mutant. Biotechnol. Lett 2004, 26, 75–78. [Google Scholar]
- Dumas, B.; Freysinet, G.; Pallett, K. Tissue-specific expression of germin-like oxalate oxidase during development and fungal infection of barley seedlings. Plant Physiol 1995, 107, 1091–1096. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem 1971, 44, 276–287. [Google Scholar]
- Godínez-Hernández, Y.; Anaya-López, J.L.; Díaz-Plaza, R.; González-Chavira, M.M.; Torres-Pacheco, I.; Rivera-Bustamante, R.F.; Guevara-González, R.G. Characterization of resistance to pepper huasteco geminivirus in chili Peppers from Yucatán, México. Hortscience 2001, 36, 139–142. [Google Scholar]
- Proteomics/post-translational_modification. Available online: http://expasy.org/tools/proteomics/post-translational_modification Accessed on 15 January 2011.





© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
León-Galván, F.; Joaquín-Ramos, A.d.J.; Torres-Pacheco, I.; De la Rosa, A.P.B.; Guevara-Olvera, L.; González-Chavira, M.M.; Ocampo-Velazquez, R.V.; Rico-García, E.; Guevara-González, R.G. A Germin-Like Protein Gene (CchGLP) of Capsicum chinense Jacq. Is Induced during Incompatible Interactions and Displays Mn-Superoxide Dismutase Activity. Int. J. Mol. Sci. 2011, 12, 7301-7313. https://doi.org/10.3390/ijms12117301
León-Galván F, Joaquín-Ramos AdJ, Torres-Pacheco I, De la Rosa APB, Guevara-Olvera L, González-Chavira MM, Ocampo-Velazquez RV, Rico-García E, Guevara-González RG. A Germin-Like Protein Gene (CchGLP) of Capsicum chinense Jacq. Is Induced during Incompatible Interactions and Displays Mn-Superoxide Dismutase Activity. International Journal of Molecular Sciences. 2011; 12(11):7301-7313. https://doi.org/10.3390/ijms12117301
Chicago/Turabian StyleLeón-Galván, Fabiola, Ahuizolt de Jesús Joaquín-Ramos, Irineo Torres-Pacheco, Ana P. Barba De la Rosa, Lorenzo Guevara-Olvera, Mario M. González-Chavira, Rosalía V. Ocampo-Velazquez, Enrique Rico-García, and Ramón Gerardo Guevara-González. 2011. "A Germin-Like Protein Gene (CchGLP) of Capsicum chinense Jacq. Is Induced during Incompatible Interactions and Displays Mn-Superoxide Dismutase Activity" International Journal of Molecular Sciences 12, no. 11: 7301-7313. https://doi.org/10.3390/ijms12117301
APA StyleLeón-Galván, F., Joaquín-Ramos, A. d. J., Torres-Pacheco, I., De la Rosa, A. P. B., Guevara-Olvera, L., González-Chavira, M. M., Ocampo-Velazquez, R. V., Rico-García, E., & Guevara-González, R. G. (2011). A Germin-Like Protein Gene (CchGLP) of Capsicum chinense Jacq. Is Induced during Incompatible Interactions and Displays Mn-Superoxide Dismutase Activity. International Journal of Molecular Sciences, 12(11), 7301-7313. https://doi.org/10.3390/ijms12117301