Membrane-Based Inverse Transition Cycling: An Improved Means for Purifying Plant-Derived Recombinant Protein-Elastin-Like Polypeptide Fusions

Abstract
:1. Introduction
2. Results and Discussion
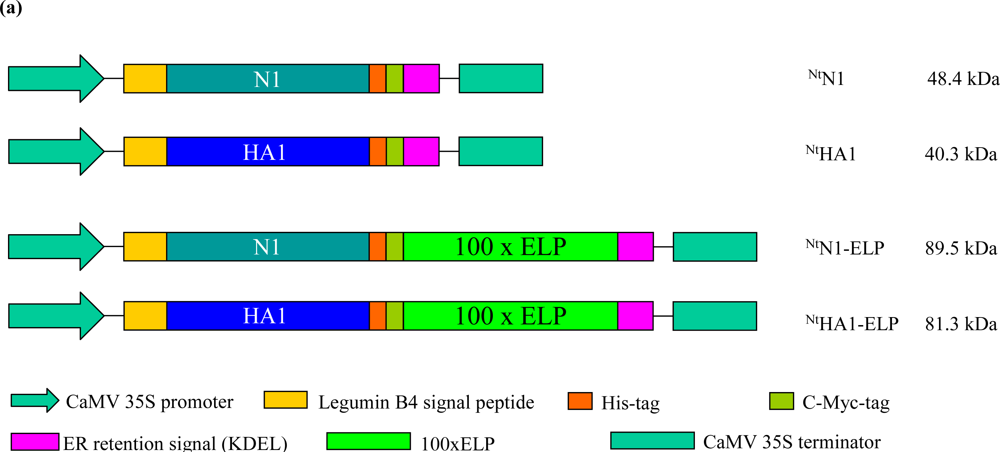
2.1. Generation of Transgenic Tobacco Plants Expressing Hemagglutinin and NA Sequences
2.2. Purification of NtHA1-ELP, NtN1-Elp Using Centrifugation-Based ITC (cITC)
2.3. Purification Using Membrane-Based ITC (mITC)
3. Experimental Section
3.1. Plant Transformation Vectors
3.2. Plant Transformation
3.3. ITC Purification of ELP Fusion Proteins
3.4. SDS-PAGE and Western Blotting
3.5. Determination of Purification Efficiency
3.6. NtN1-ELP Activity Assay
3.7. Competitive ELISA
4. Conclusions
Acknowledgments
References
- Curtiss, RI; Cardineau, CA. Oral immunisation by transgenic plants. U.S. Patent 5,679,880. 1990. [Google Scholar]
- Hiatt, A; Caffferkey, R; Bowdish, K. Production of antibodies in transgenic plants. Nature 1989, 342, 76–78. [Google Scholar]
- Ma, JK; Drake, PM; Christou, P. The production of recombinant pharmaceutical proteins in plants. Nat. Rev. Genet 2003, 4, 794–805. [Google Scholar]
- Ma, JK; Barros, E; Bock, R; Christou, P; Dale, PJ; Dix, PJ; Fischer, R; Irwin, J; Mahoney, R; Pezzotti, M; et al. Molecular farming for new drugs and vaccines. Current perspectives on the production of pharmaceuticals in transgenic plants. EMBO Rep 2005, 6, 593–599. [Google Scholar]
- Stoger, E; Ma, JK; Fischer, R; Christou, P. Sowing the seeds of success: Pharmaceutical proteins from plants. Curr. Opin. Biotechnol 2005, 16, 167–173. [Google Scholar]
- Streatfield, SJ; Howard, JA. Plant production systems for vaccines. Expert Rev. Vaccines 2003, 2, 763–775. [Google Scholar]
- Warzecha, H; Mason, HS. Benefits and risks of antibody and vaccine production in transgenic plants. J. Plant Physiol 2003, 160, 755–764. [Google Scholar]
- Yusibov, V; Rabindran, S. Recent progress in the development of plant derived vaccines. Expert Rev. Vaccines 2008, 7, 1173–1183. [Google Scholar]
- Hassan, S; van Dolleweerd, CJ; Ioakeimidis, F; Keshavarz-Moore, E; Ma, JKC. Considerations for extraction of monoclonal antibodies targeted to different subcellular compartments in transgenic tobacco plants. Plant Biotechnol. J 2008, 6, 733–748. [Google Scholar]
- Hood, EE; Woodard, SL; Horn, ME. Monoclonal antibody manufacturing in transgenic plants-myths and realities. Curr. Opin. Biotechnol 2002, 13, 630–635. [Google Scholar]
- Hussack, G; Grohs, BM; Almquist, KC; McLean, MD; Ghosh, R; Hall, JC. Purification of plant-derived antibodies through direct immobilization of affinity ligands on cellulose. J. Agric. Food Chem 2010, 58, 3451–3459. [Google Scholar]
- Floss, DM; Conrad, U. Plant Molecular Pharming–Veterinary Applications. In Springer Encycl. Sustainability Sci. Technol.; 2011; in press. [Google Scholar]
- Floss, DM; Schallau, K; Rose-John, S; Conrad, U; Scheller, J. Elastin-like polypeptides revolutionize recombinant protein expression and their biomedical application. Trends Biotechnol 2010, 28, 37–45. [Google Scholar]
- Conley, AJ; Joensuu, JJ; Richman, A; Menassa, R. Protein body-inducing fusions for high-level production and purification of recombinant proteins in plants. Plant Biotechnol. J 2011, 9, 419–433. [Google Scholar]
- Floss, DM; Mockey, M; Zanello, G; Brosson, D; Diogon, M; Frutos, R; Bruel, T; Rodrigues, V; Garzon, E; Chevaleyre, C; et al. Expression and Immunogenicity of the Mycobacterial Ag85B/ESAT-6 antigens produced in transgenic plants by elastin-like peptide fusion strategy. J. Biomed. Biotechnol 2010, 2010, 1–15. [Google Scholar]
- Floss, DM; Sack, M; Stadlmann, J; Rademacher, T; Scheller, J; Stöger, E; Fischer, R; Conrad, U. Biochemical and functional characterization of anti-HIV antibody–ELP fusion proteins from transgenic plants. Plant Biotechnol. J 2008, 6, 379–391. [Google Scholar]
- Floss, DM; Sack, M; Arcalis, E; Stadlmann, J; Quendler, H; Rademacher, T; Stoger, E; Scheller, JR; Fischer, R; Conrad, U. Influence of elastin-like peptide fusions on the quantity and quality of a tobacco-derived human immunodeficiency virus-neutralizing antibody. Plant Biotechnol. J 2009, 7, 899–913. [Google Scholar]
- Scheller, J; Leps, M; Conrad, U. Forcing single-chain variable fragment production in tobacco seeds by fusion to elastin-like polypeptides. Plant Biotechnol. J 2006, 4, 243–249. [Google Scholar]
- Conrad, U; Plagmann, I; Malchow, S; Sack, M; Floss, DM; Kruglov, AA; Nedospasov, SA; Rose-John, S; Scheller, J. ELPylated anti-human TNF therapeutic single-domain antibodies for prevention of lethal septic shock. Plant Biotechnol. J 2011, 9, 22–31. [Google Scholar]
- Meyer, DE; Chilkoti, A. Purification of recombinant proteins by fusion with thermally-responsive polypeptides. Nat. Biotechnol 1999, 17, 1112–1115. [Google Scholar]
- Ge, X; Trabbic-Carlson, K; Chilkoti, A; Filipe, CDM. Purification of an elastin-like fusion protein by microfiltration. Biotechnol. Bioeng 2006, 95, 424–432. [Google Scholar]
- Capua, I; Alexander, DJ. Avian influenza: recent developments. Avian Pathol 2004, 33, 393–404. [Google Scholar]
- Capua, I; Alexander, DJ. Animal and human health implications of avian influenza infections. Biosci. Rep 2007, 27, 359–372. [Google Scholar]
- Shoji, Y; Chichester, JA; Bi, H; Musiychuk, K; de la Rosa, P; Goldschmidt, L; Horsey, A; Ugulava, N; Palmer, GA; Mett, V. Plant-expressed HA as a seasonal influenza vaccine candidate. Vaccine 2008, 26, 2930–2934. [Google Scholar]
- Shoji, Y; Farrance, CE; Bi, H; Shamloul, M; Green, B; Manceva, S; Rhee, A; Ugulava, N; Roy, G; Musiychuk, K. Immunogenicity of hemagglutinin from A/Bar-headed Goose/Qinghai/1A/05 and A/Anhui/1/05 strains of H5N1 influenza viruses produced in Nicotiana benthamiana plants. Vaccine 2009, 27, 3467–3470. [Google Scholar]
- Mett, V; Musiychuk, K; Bi, H; Farrance, CE; Horsey, A; Ugulava, N; Shoji, Y; de la Rosa, P; Palmer, GA; Rabindran, S; et al. A plant-produced influenza subunit vaccine protects ferrets against virus challenge. Influenza Other Res. Viruses 2008, 2, 33–40. [Google Scholar]
- Shoji, Y; Bi, H; Musiychuk, K; Rhee, A; Horsey, A; Roy, G; Green, B; Shamloul, M; Farrance, CE; Taggart, B. Plant-derived hemagglutinin protects ferrets against challenge infection with the A/Indonesia/05/05 strain of avian influenza. Vaccine 2009, 27, 1087–1092. [Google Scholar]
- Kalthoff, D; Giritch, A; Geisler, K; Bettmann, U; Klimyuk, V; Hehnen, HR; Gleba, Y; Beer, M. Immunization with plant-expressed hemagglutinin protects chickens from lethal highly pathogenic avian influenza virus H5N1 challenge infection. J. Virol 2010, 84, 12002–12010. [Google Scholar]
- Spitsin, S; Andrianov, V; Pogrebnyak, N; Smirnov, Y; Borisjuk, N; Portocarrero, C; Veguilla, V; Koprowski, H; Golovkin, M. Immunological assessment of plant-derived avian flu H5/HA1 variants. Vaccine 2009, 27, 1289–1292. [Google Scholar]
- Sylte, M; Hubby, B; Suarez, D. Influenza neuraminidase antibodies provide partial protection for chickens against high pathogenic avian influenza infection. Vaccine 2007, 25, 3763–3772. [Google Scholar]
- Deroo, T; Jou, WM; Fiers, W. Recombinant neuraminidase vaccine protects against lethal influenza. Vaccine 1996, 14, 561–569. [Google Scholar]
- Skehel, JJ; Wiley, DC. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem 2000, 69, 531–569. [Google Scholar]
- Wandelt, CI; Khan, MR; Craig, S; Schroeder, HE; Spencer, D; Higgins, TJ. Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants. Plant J 1992, 2, 181–192. [Google Scholar]
- Conrad, U; Fiedler, U. Compartment-specific accumulation of recombinant immunoglobulins in plant cells: An essential tool for antibody production and immunomodulation of physiological functions and pathogen activity. Plant Mol. Biol 1998, 38, 101–109. [Google Scholar]
- Scheller, J; Henggeler, D; Viviani, A; Conrad, U. Purification of spider silk-elastin from transgenic plants and application for human chondrocyte proliferation. Transgenic Res 2004, 13, 51–57. [Google Scholar]
- Conley, AJ; Joensuu, JJ; Jevnikar, AM; Menassa, R; Brandle, JE. Optimization of elastin-like polypeptide fusions for expression and purification of recombinant proteins in plants. Biotechnol. Bioeng 2009, 103, 562–573. [Google Scholar]
- Xiang, C; Han, P; Lutziger, I; Wang, K; Oliver, D. A mini binary vector series for plant transformation. Plant Mol. Biol 1999, 40, 711–717. [Google Scholar]
- Deblaere, R; Bytebier, B; de Greve, H; Deboeck, F; Schell, J; van Montagu, M; Leemans, J. Efficient octopine Ti plasmid-derived vectors for Agrobacterium-mediated gene transfer to plants. Nucleic Acids Res 1985, 13, 4777–4788. [Google Scholar]
- Horsch, RB; Fry, JE; Hoffmann, NL; Eichholtz, D; Rogers, SG; Fraley, RT. A Simple and General Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Potier, M; Mameli, L; Belisle, M; Dallaire, L; Melancon, SB. Fluorometric assay of neuraminidase with a sodium (4-methylumbelliferyl-alpha-d-N-acetylneuraminate) substrate. Anal. Biochem 1979, 94, 287–296. [Google Scholar]
- Yongkiettrakul, S; Boonyapakron, K; Jongkaewwattana, A; Wanitchang, A; Leartsakulpanich, U; Chitnumsub, P; Eurwilaichitr, L; Yuthavong, Y. Avian influenza A/H5N1 neuraminidase expressed in yeast with a functional head domain. J. Virol. Methods 2009, 156, 44–51. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transgene | Number of Regenerated Plants | Number of Transgene Expressing Plants | TransgeneExpression (%TSP) |
|---|---|---|---|
| NtN1 | 42 | 23 | 0.004% |
| NtN1-ELP | 37 | 23 | 0.2% |
| NtHA1 | 36 | 22 | 0.01% |
| NtHA1-ELP | 88 | 67 | 0.3% |
| Recombinant proteins | Recovery (%) | |
|---|---|---|
| mITC | cITC | |
| NtHA1-ELP | 95* | 98* |
| NtN1-ELP | 92.5**–94* | 33.7**–40* |
| Ntanti-hTNFα-VHH-ELP | 98* | 11* [19] |
| Status of NtN1-ELP | Total protein (mg) | Total activity (U) | Recovery rate (%) | Specific activity (U mg−1) |
|---|---|---|---|---|
| Raw extract | 3521.9 | 736.5 | 100 | 0.21 |
| Purified NtN1-ELP by mITC | 20.3 | 681.1 | 92.5 | 33.6 |
| Purified NtN1-ELP by cITC | 31.2 | 248.5 | 33.7 | 7.96 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Phan, H.T.; Conrad, U. Membrane-Based Inverse Transition Cycling: An Improved Means for Purifying Plant-Derived Recombinant Protein-Elastin-Like Polypeptide Fusions. Int. J. Mol. Sci. 2011, 12, 2808-2821. https://doi.org/10.3390/ijms12052808
Phan HT, Conrad U. Membrane-Based Inverse Transition Cycling: An Improved Means for Purifying Plant-Derived Recombinant Protein-Elastin-Like Polypeptide Fusions. International Journal of Molecular Sciences. 2011; 12(5):2808-2821. https://doi.org/10.3390/ijms12052808
Chicago/Turabian StylePhan, Hoang Trong, and Udo Conrad. 2011. "Membrane-Based Inverse Transition Cycling: An Improved Means for Purifying Plant-Derived Recombinant Protein-Elastin-Like Polypeptide Fusions" International Journal of Molecular Sciences 12, no. 5: 2808-2821. https://doi.org/10.3390/ijms12052808
APA StylePhan, H. T., & Conrad, U. (2011). Membrane-Based Inverse Transition Cycling: An Improved Means for Purifying Plant-Derived Recombinant Protein-Elastin-Like Polypeptide Fusions. International Journal of Molecular Sciences, 12(5), 2808-2821. https://doi.org/10.3390/ijms12052808