Antioxidant Protective Effect of Honey in Cigarette Smoke-Induced Testicular Damage in Rats

Abstract
:1. Introduction
2. Results and Discussion
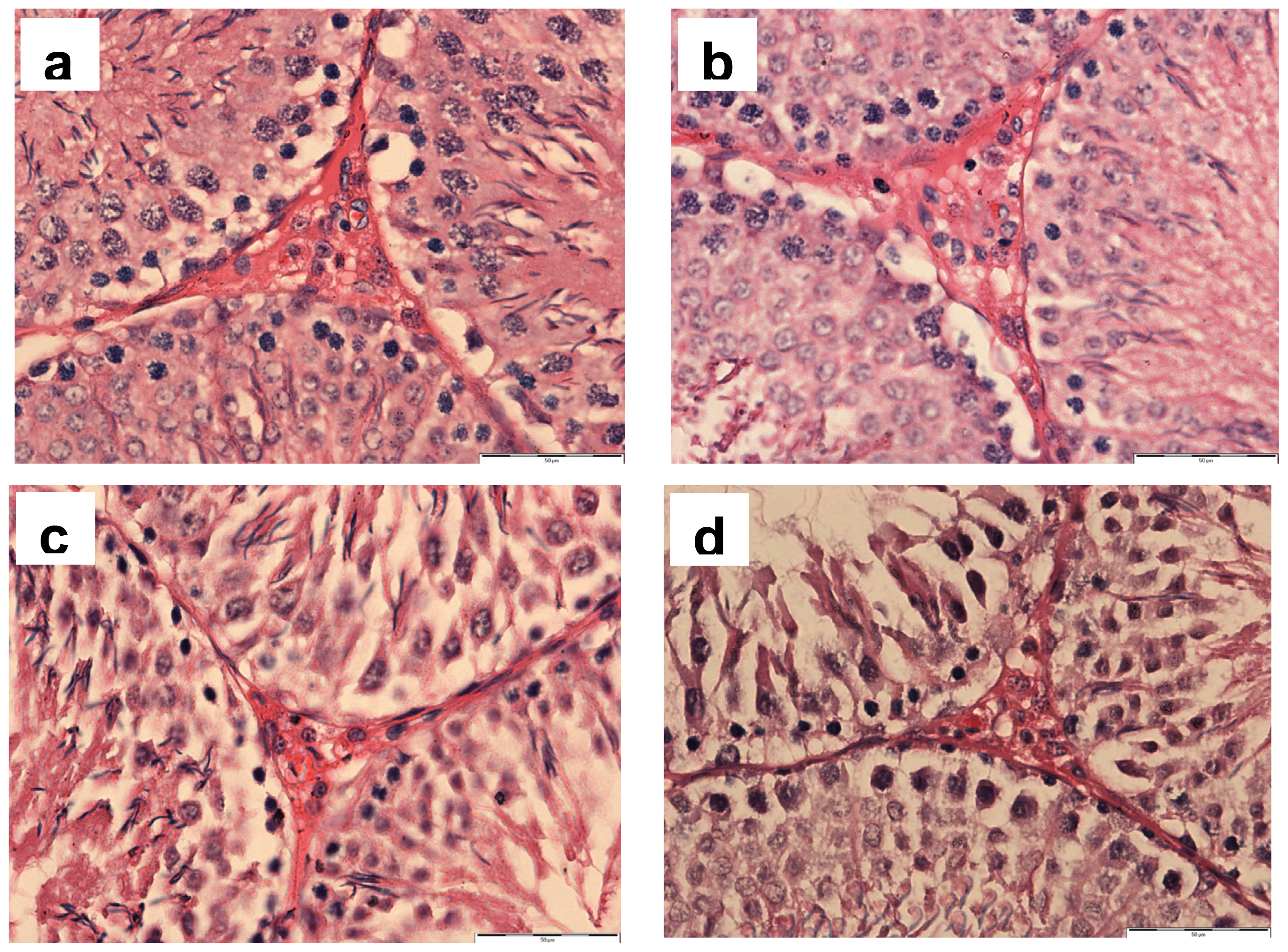
2.1. Histopathology of Testis
2.2. Biochemical Analyses
3. Experimental Section
3.1. Chemicals
3.2. Animals
3.3. Experimental Design
3.4. Histopathological Examination
3.5. Biochemical Analysis
3.5.1. Oxidative Stress Markers Assays in Rat Testis
3.5.2. Determination of the Antioxidant Capacities of Honey
3.6. Statistical Analyses
4. Conclusions
Acknowledgements
- Conflict of InterestThe authors declare that there are no conflicts of interest.
References
- Richthoff, J; Elzanaty, S; Rylander, L; Hagmar, L; Giwercman, A. Association between tobacco exposure and reproductive parameters in adolescent males. Int J Androl 2008, 31, 31–39. [Google Scholar]
- Gaur, DS; Talekar, M; Pathak, VP. Effect of cigarette smoking on semen quality of infertile men. Singapore Med J 2007, 48, 119–123. [Google Scholar]
- Evans, HJ; Fletcher, J; Torrance, M; Hargreave, TB. Sperm abnormalities and cigarette smoking. Lancet 1981, 1, 627–629. [Google Scholar]
- Potts, RJ; Newbury, CJ; Smith, G; Notarianni, LJ; Jefferies, TM. Sperm chromatin damage associated with male smoking. Mutat Res 1999, 423, 103–111. [Google Scholar]
- Sepaniak, S; Forges, T; Gerard, H; Foliguet, B; Bene, MC; Monnier-Barbarino, P. The influence of cigarette smoking on human sperm quality and DNA fragmentation. Toxicology 2006, 223, 54–60. [Google Scholar]
- Yardimci, S; Atan, A; Delibasi, T; Sunguroglu, K; Guven, MC. Long-term effects of cigarettesmoke exposure on plasma testosterone, luteinizing hormone and follicle-stimulating hormone levels in male rats. Br J Urol 1997, 79, 66–69. [Google Scholar]
- Rajpurkar, A; Li, H; Dhabuwala, CB. Morphometric analysis of rat testis following chronic exposure to cigarette smoke. J Environ Pathol Toxicol Oncol 2000, 19, 363–368. [Google Scholar]
- Ahmadnia, H; Ghanbari, M; Moradi, MR; Khaje-Dalouee, M. Effect of cigarette smoke on spermatogenesis in rats. Urol J 2007, 4, 159–163. [Google Scholar]
- Rajpurkar, A; Dhabuwala, CB; Jiang, Y; Li, H. Chronic cigarette smoking induces an oxidantantioxidant imbalance in the testis. J Environ Pathol Toxicol Oncol 2000, 19, 369–373. [Google Scholar]
- Kücük, M; Kolaylı, S; Karaođlu, S; Ulusoy, E; Baltacı, C; Candan, F. Biological activities and chemical composition of three honeys of different types from Anatolia. Food Chem 2007, 100, 526–534. [Google Scholar]
- Gheldof, N; Wang, XH; Engeseth, NJ. Identification and quantification of antioxidant components of honeys from various floral sources. J Agric Food Chem 2002, 50, 5870–5877. [Google Scholar]
- Mato, I; Huidobro, JF; Simal-Lozano, J; Sancho, MT. Significance of nonaromatic organic acids in honey. J Food Prot 2003, 66, 2371–2376. [Google Scholar]
- Al-Waili, NS. Effects of daily consumption of honey solution on hematological indices and blood levels of minerals and enzymes in normal individuals. J Med Food 2003, 6, 135–140. [Google Scholar]
- Yao, L; Jiang, Y; D’Arcy, B; Singanusong, R; Datta, N; Caffin, N; Raymont, K. Quantitative high-performance liquid chromatography analyses of flavonoids in Australian Eucalyptus honeys. J Agric Food Chem 2004, 52, 210–214. [Google Scholar]
- Michalkiewicz, A; Biesaga, M; Pyrzynska, K. Solid-phase extraction procedure for determination of phenolic acids and some flavonols in honey. J Chromatogr A 2008, 1187, 18–24. [Google Scholar]
- Tan, HT; Rahman, RA; Gan, SH; Halim, AS; Hassan, SA; Sulaiman, SA; Kirnpal-Kaur, B. The antibacterial properties of Malaysian tualang honey against wound and enteric microorganisms in comparison to manuka honey. BMC Complement Altern Med 2009, 9, 34. [Google Scholar]
- Molan, PC. The Antibacterial Activity of Honey. 1. The Nature of the Antibacterial Activity. Bee World 1992, 73, 5–28. [Google Scholar]
- Prakash, A; Medhi, B; Avti, PK; Saikia, UN; Pandhi, P; Khanduja, KL. Effect of different doses of Manuka honey in experimentally induced inflammatory bowel disease in rats. Phytother Res 2008, 22, 1511–1519. [Google Scholar]
- Bilsel, Y; Bugra, D; Yamaner, S; Bulut, T; Cevikbas, U; Turkoglu, U. Could honey have a place in colitis therapy? Effects of honey, prednisolone, and disulfiram on inflammation, nitric oxide, and free radical formation. Dig Surg 2002, 19, 306–312. [Google Scholar]
- Berretta, G; Granata, P; Ferrero, M; Orioli, M; Facino, RM. Standardization of antioxidant properties of honey by combination of spectrophotometric/fluorometric assays and chemometrics. Anal Chim Acta 2005, 533, 185–191. [Google Scholar]
- Perez, E; Rodriguez-Malaver, AJ; Vit, P. Antioxidant capacity of Venezuelan honey in wistar rat homogenates. J Med Food 2006, 9, 510–516. [Google Scholar]
- Mohamed, M; Sirajudeen, KNS; Swamy, M; Yaacob, NS; Sulaiman, SA. Studies on the antioxidant properties of tualang honey of Malaysia. Afr J Trad CAM 2010, 7, 59–63. [Google Scholar]
- Mahaneem, M; Sulaiman, SA; Jaafar, H; Sirajudeen, KNS. Effect of honey on testicular functions in rats exposed to cigarette smoke. J ApiProd ApiMed Sci 2011, 3, 12–17. [Google Scholar]
- Ozyurt, H; Pekmez, H; Parlaktas, BS; Kus, I; Ozyurt, B; Sarsilmaz, M. Oxidative stress in testicular tissues of rats exposed to cigarette smoke and protective effects of caffeic acid phenethyl ester. Asian J Androl 2006, 8, 189–193. [Google Scholar]
- Church, DF; Pryor, WA. Free-radical chemistry of cigarette smoke and its toxicological implications. Environ Health Perspect 1985, 64, 111–126. [Google Scholar]
- Bartalis, J; Zhao, YL; Flora, JW; Paine, JB; Wooten, JB. Carbon-centered radicals in cigarette smoke: acyl and alkylaminocarbonyl radicals. Anal Chem 2009, 81, 631–641. [Google Scholar]
- Alvarez, JG; Storey, BT. Differential incorporation of fatty acids into and peroxidative loss of fatty acids from phospholipids of human spermatozoa. Mol Reprod Dev 1995, 42, 334–346. [Google Scholar]
- Smith, C; Marks, AD; Liebermann, ML. Oxygen Toxicity and Free Radical Injury. In Mark’s Basic Medical Biochemistry: A Clinical Approach, 2nd ed; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2005; pp. 439–457. [Google Scholar]
- Somogyi, A; Rosta, K; Pusztai, P; Tulassay, Z; Nagy, G. Antioxidant measurements. Physiol Meas 2007, 28, R41–R55. [Google Scholar]
- Kohen, R; Nyska, A. Oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol Pathol 2002, 30, 620–650. [Google Scholar]
- Tian, L; Shi, MM; Forman, HJ. Increased transcription of the regulatory subunit of gammaglutamylcysteine synthetase in rat lung epithelial L2 cells exposed to oxidative stress or glutathione depletion. Arch Biochem Biophys 1997, 342, 126–133. [Google Scholar]
- Lu, SC. Regulation of glutathione synthesis. Mol Aspects Med 2009, 30, 42–59. [Google Scholar]
- Moskaug, JO; Carlsen, H; Myhrstad, MC; Blomhoff, R. Polyphenols and glutathione synthesis regulation. Am J Clin Nutr 2005, 81, 277S–283S. [Google Scholar]
- Beretta, G; Orioli, M; Facino, RM. Antioxidant and radical scavenging activity of honey in endothelial cell cultures. Planta Med 2007, 73, 1182–1189. [Google Scholar]
- Cesar-Neto, JB; Duarte, PM; Sallum, EA; Barbieri, D; Moreno, H, Jr; Nociti, FH, Jr. A comparative study on the effect of nicotine administration and cigarette smoke inhalation on bone healing around titanium implants. J Periodontol 2003, 74, 1454–1459. [Google Scholar]
- Faqi, AS; Johnson, WD; Morrissey, RL; McCormick, DL. Reproductive toxicity assessment of chronic dietary exposure to soy isoflavones in male rats. Reprod Toxicol 2004, 18, 605–611. [Google Scholar]
- D’Souza, UJ. Effect of tamoxifen on spermatogenesis and tubular morphology in rats. Asian J Androl 2004, 6, 223–226. [Google Scholar]
- Watanabe, N; Kamei, S; Ohkubo, A; Yamanaka, M; Ohsawa, S; Makino, K; Tokuda, K. Urinary protein as measured with a pyrogallol red-molybdate complex, manually and in a Hitachi 726 automated analyzer. Clin Chem 1986, 32, 1551–1554. [Google Scholar]
- Ohkawa, H; Ohishi, N; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 1979, 95, 351–358. [Google Scholar]
- Koracevic, D; Koracevic, G; Djordjevic, V; Andrejevic, S; Cosic, V. Method for the measurement of antioxidant activity in human fluids. J Clin Pathol 2001, 54, 356–361. [Google Scholar]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin Chim Acta 1991, 196, 143–151. [Google Scholar]
- Luchese, C; Pinton, S; Nogueira, CW. Brain and lungs of rats are differently affected by cigarette smoke exposure: antioxidant effect of an organoselenium compound. Pharmacol Res 2009, 59, 194–201. [Google Scholar]
- Habig, WH; Pabst, MJ; Jakoby, WB. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J Biol Chem 1974, 249, 7130–7139. [Google Scholar]
- Beretta, G; Granata, P; Ferrero, M; Orioli, M; Facino, RM. Standardization of antioxidant properties of honey by combination of spectrophotometric/fluorometric assays and chemometrics. Anal Chim Acta 2005, 533, 185–191. [Google Scholar]
- Benzie, IF; Strain, JJ. Ferric reducing/antioxidant power assay: direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol 1999, 299, 15–27. [Google Scholar]
- Chen, L; Mehta, A; Berenbaum, M; Zangerl, AR; Engeseth, NJ. Honeys from different floral sources as inhibitors of enzymatic browning in fruit and vegetable homogenates. J Agric Food Chem 2000, 48, 4997–5000. [Google Scholar]
- Aljadi, AM; Kamaruddin, MY. Evaluation of the phenolic contents and antioxidant capacities of two Malaysian floral honeys. Food Chem 2004, 85, 513–518. [Google Scholar]



{kind=link}
{kind=link}
| Parameters | Control Group | H Group | CS Group | H + CS Group | p (KW test) |
|---|---|---|---|---|---|
| Seminiferous tubule diameter (μm) | 276.32 (255.22–286.97) | 260.74 (256.94–275.27) | 246.24 (237.19–256.50)a,b | 267.46 (258.72–283.21)c | <0.05 |
| Seminiferous epithelial height (μm) | 81.80 (76.05–90.26) | 78.60 (76.39–83.17) | 71.01 (66.95–74.82)a,b | 77.47 (73.19–80.79)c | <0.05 |
| Seminiferous tubule with germ cell loss (%) | 0.00 (0.00–1.00) | 0.00 (0.00–0.63) | 7.50 (4.88–15.50)a,b | 2.25 (2.25–5.13)a,b,c | <0.001 |
| Leydig cell count | 206.5 (197.8–219.0) | 195.5 (178.8–219.0) | 154.5 (138.5–162.5)a,b | 175.0 (165.3–190.3)a,c | <0.01 |
| Parameters | Control group | H group | CS group | H + CS group | p |
|---|---|---|---|---|---|
| TBARS (nmol MDA Eq/mg protein)# | 0.29 (0.26–0.41) | 0.34 (0.19–0.49) | 0.68 (0.54–0.87)a,b | 0.32 (0.27–0.41)c | <0.01£ |
| TAS (μmol uric acid Eq/mg protein) @ | 0.23 ± 0.02 | 0.26 ± 0.02 a | 0.14 ± 0.03 a,b | 0.19 ± 0.03 b,c | <0.001 § |
| SOD activity (unit/mg protein) # | 1.30 (1.21–1.43) | 1.23 (0.89–1.31) | 0.82 (0.68–0.94) a,b | 1.09 (0.95–1.51) c | <0.01 £ |
| CAT activity (unit/mg protein) # | 20.54 (17.58–26.52) | 25.25 (22.51–28.44) | 16.30 (14.38–19.21) a,b | 20.21 (19.06–22.64)b,c | <0.01 £ |
| Total glutathione (nmol GSH Eq/mg protein) @ | 1.45 ± 0.14 | 1.46 ± 0.34 | 1.40 ± 0.33 | 1.93 ± 0.50 a,c | <0.05 $ |
| GPx activity (unit × 103/mg protein) # | 166.56 (161.45–174.22) | 168.41 (153.41–195.44) | 183.69 (179.83–195.67) a | 163.26 (156.38–170.99) c | <0.05 £ |
| GR activity (unit/mg protein) @ | 20.64 ± 1.39 | 20.50 ± 2.63 | 20.29 ± 1.53 | 21.31 ± 1.17 | NS § |
| GST activity (unit × 103/mg protein) # | 0.75 (0.67–0.80) | 0.73 (0.68–0.89) | 0.66 (0.63–0.71) | 0.70 (0.68–0.74) | NS £ |
| Parameters | |
|---|---|
| Total phenolic content | 219.53 ± 8.98 mg of gallic acid Eq/kg of honey |
| Antioxidant activity (FRAP assay) | 369.79 ± 21.46 μmol of Fe2+ Eq/L |
| Free radical scavenging activity (DPPH assay) | 47.25 ± 0.62% inhibition of DPPH radical |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mohamed, M.; Sulaiman, S.A.; Jaafar, H.; Sirajudeen, K.N.S. Antioxidant Protective Effect of Honey in Cigarette Smoke-Induced Testicular Damage in Rats. Int. J. Mol. Sci. 2011, 12, 5508-5521. https://doi.org/10.3390/ijms12095508
Mohamed M, Sulaiman SA, Jaafar H, Sirajudeen KNS. Antioxidant Protective Effect of Honey in Cigarette Smoke-Induced Testicular Damage in Rats. International Journal of Molecular Sciences. 2011; 12(9):5508-5521. https://doi.org/10.3390/ijms12095508
Chicago/Turabian StyleMohamed, Mahaneem, Siti Amrah Sulaiman, Hasnan Jaafar, and Kuttulebbai Nainamohamed Salam Sirajudeen. 2011. "Antioxidant Protective Effect of Honey in Cigarette Smoke-Induced Testicular Damage in Rats" International Journal of Molecular Sciences 12, no. 9: 5508-5521. https://doi.org/10.3390/ijms12095508
APA StyleMohamed, M., Sulaiman, S. A., Jaafar, H., & Sirajudeen, K. N. S. (2011). Antioxidant Protective Effect of Honey in Cigarette Smoke-Induced Testicular Damage in Rats. International Journal of Molecular Sciences, 12(9), 5508-5521. https://doi.org/10.3390/ijms12095508