Local Mechanical Stimulation of Mardin-Darby Canine Kidney Cell Sheets on Temperature-Responsive Hydrogel

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
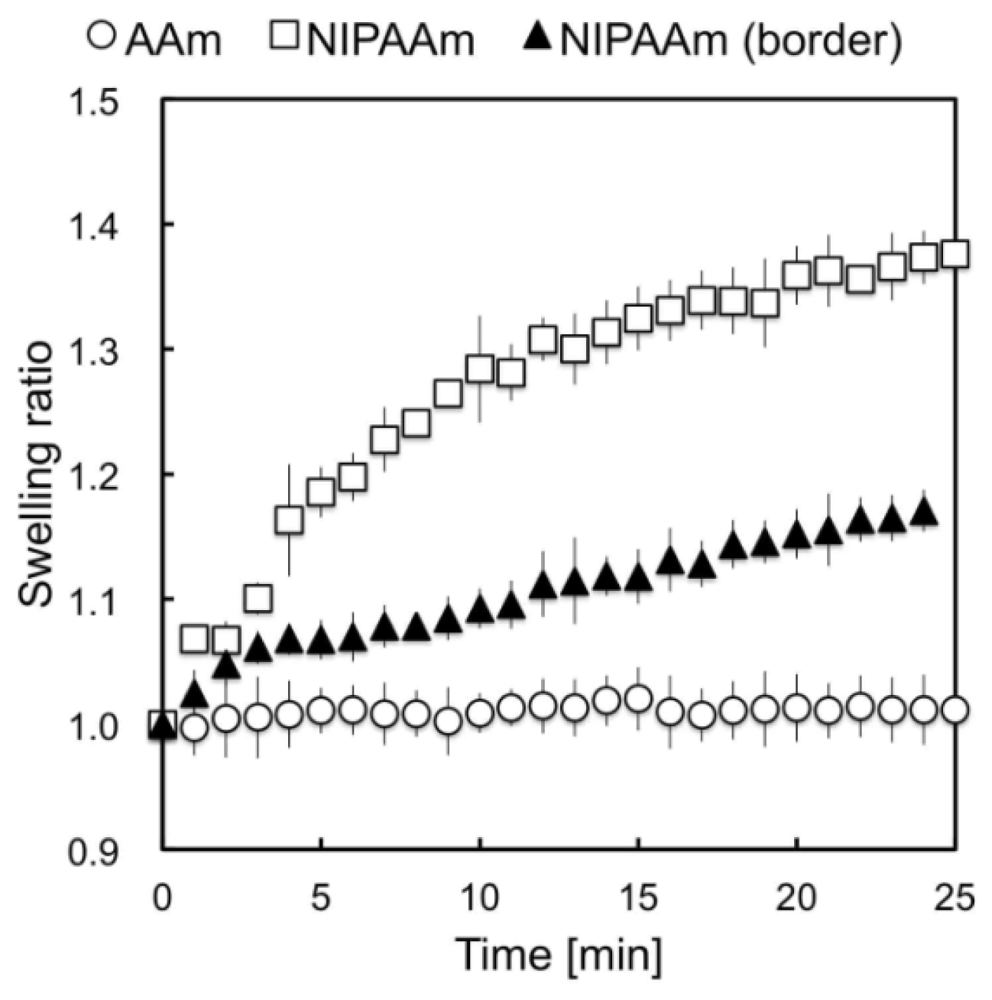
2.1. Preparation of Collagen-Immobilized PNIPAAm/PAAm Combined Gel Culture Substrate
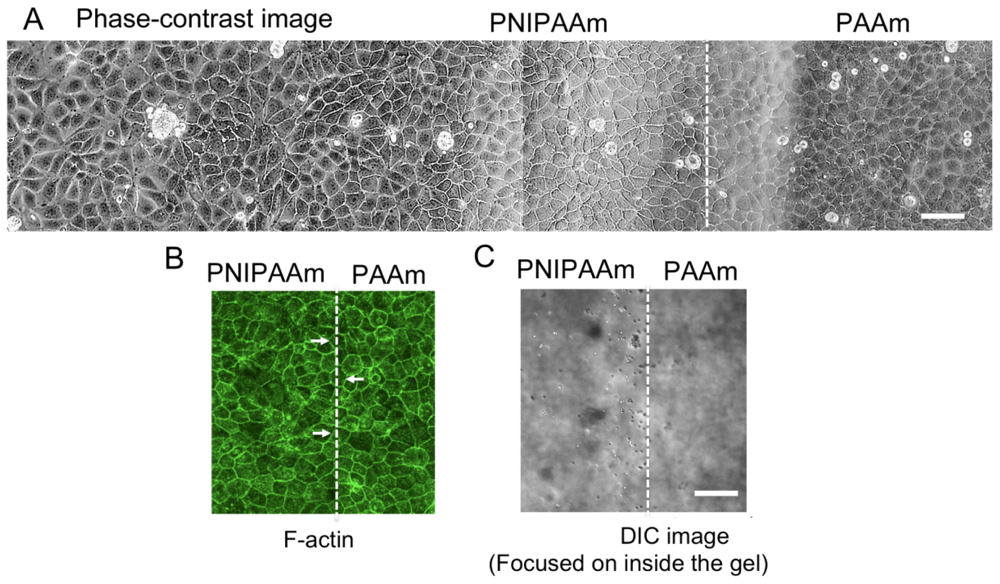
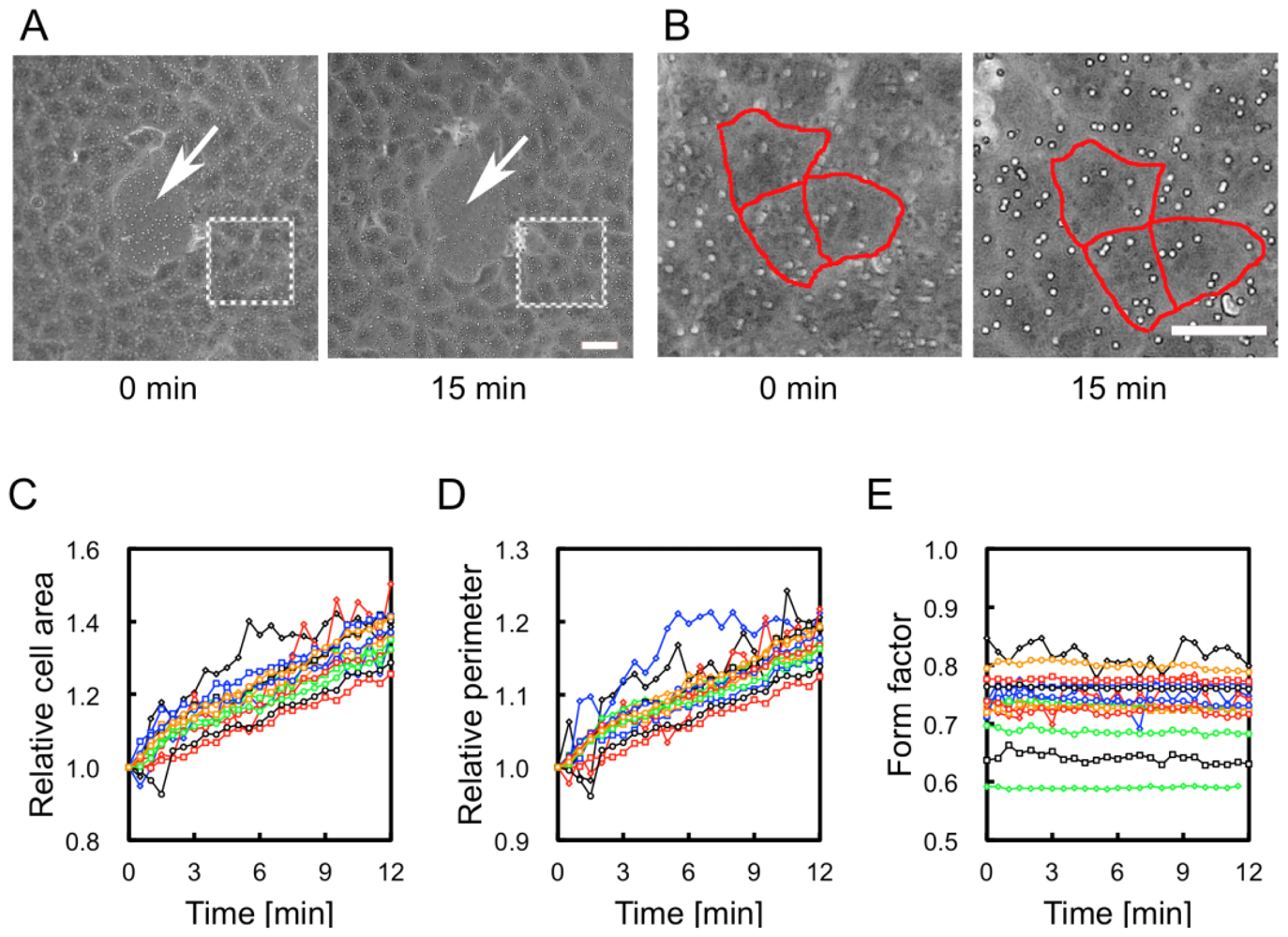
2.2. Observations of Change in Cell Shape on the collagen-Immobilized PNIPAAm Gel Side with Temperature Change
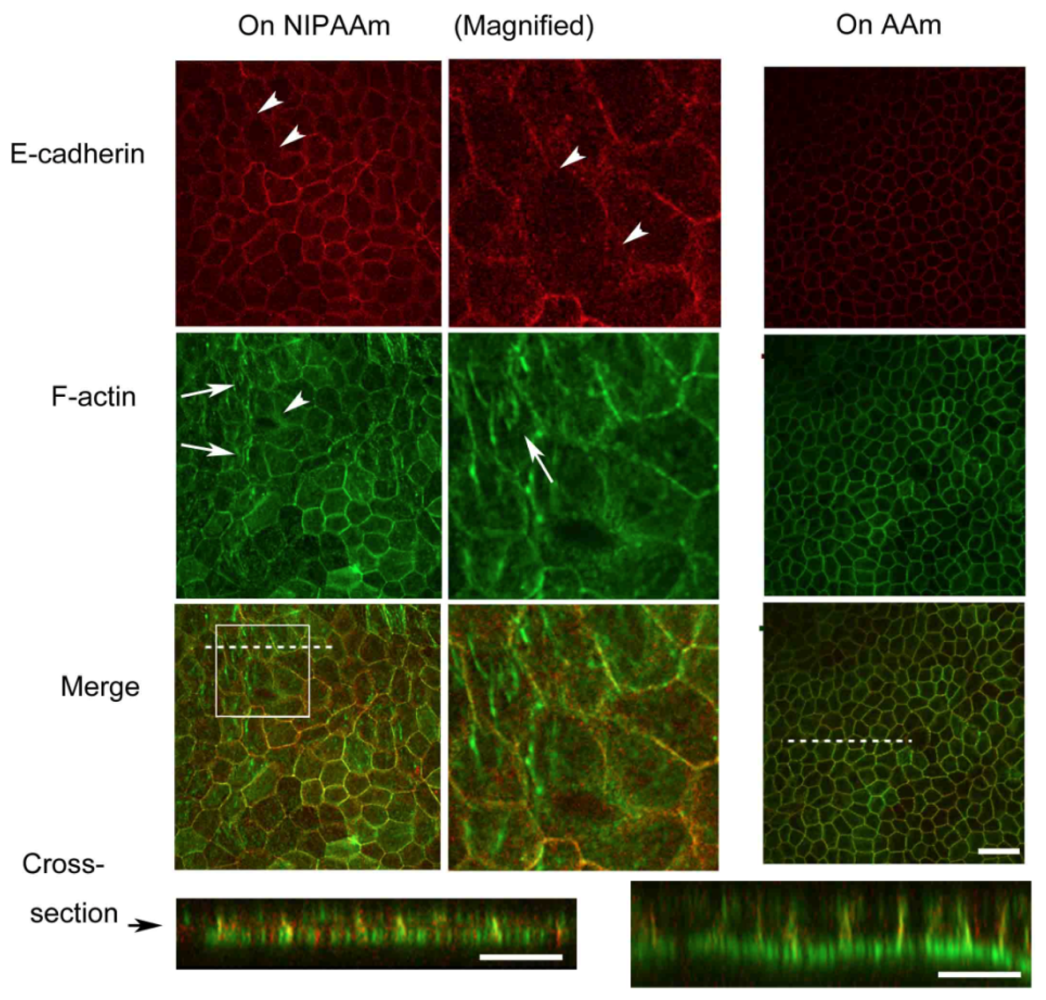
2.3. Observation of Adherens Junctions
2.4. Observation of ERK1/2 Signaling and Initial Propagation
3. Experimental Section
3.1. Cells and Reagents
3.2. Preparation of Collagen-Immobilized PNIPAAm/PAAm Combined Hydrogel
3.3 Measurement of Temperature-Sensitivity of the Collagen-Immobilized PNIPAAm/PAAm Combined Hydrogel
3.4. Observation and Quantification of Changes in Cell Shape
3.5. Immunostaining and Quantification
3.6. Statistical Analysis
4. Conclusion
Supplementary Material
Acknowledgements
References
- Trepat, X.; Fredberg, J.J. Plithotaxis and emergent dynamics in collective cellular migration. Trends Cell Biol 2011, 21, 638–646. [Google Scholar]
- Poujade, M.; Grasland-Mongrain, E.; Hertzog, A.; Jouanneau, J.; Chavrier, P.; Ladoux, B.; Buguin, A.; Silberzan, P. Collective migration of an epithelial monolayer in response to a model wound. Proc. Natl. Acad. Sci. USA 2007, 104, 15988–15993. [Google Scholar]
- Fenteany, G.; Janmey, P.A.; Stossel, T.P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Curr. Biol 2000, 10, 831–838. [Google Scholar]
- Vaezi, A.; Bauer, C.; Vasioukhin, V.; Fuchs, E. Actin cable dynamics and Rho/Rock orchestrate a polarized cytoskeletal architecture in the early steps of assembling a stratified epithelium. Dev. Cell 2002, 3, 367–381. [Google Scholar]
- Matsubayashi, Y.; Ebisuya, M.; Honjoh, S.; Nishida, E. Erk activation propagates in epithelial cell sheets and regulates their migration during wound healing. Curr. Biol 2004, 14, 731–735. [Google Scholar]
- Nikolic, D.L.; Boettiger, A.N.; Bar-Sagi, D.; Carbeck, J.D.; Shvartsman, S.Y. Role of boundary conditions in an experimental model of epithelial wound healing. Am. J. Physiol. Cell Physiol 2006, 291, C68–C75. [Google Scholar]
- Hirokawa, Y.; Tanaka, T. Volume phase-transition in a nonionic gel. J. Chem. Phys 1984, 81, 6379–6380. [Google Scholar]
- Bae, Y.H.; Okano, T.; Kim, S.W. “On-off” thermocontrol of solute transport. I. Temperature dependence of swelling of N-isopropylacrylamide networks modified with hydrophobic components in water. Pharm. Res 1991, 8, 531–537. [Google Scholar]
- Tang, Z.; Akiyama, Y.; Yamato, M.; Okano, T. Comb-type grafted poly(N-isopropylacrylamide) gel modified surfaces for rapid detachment of cell sheet. Biomaterials 2010, 31, 7435–7443. [Google Scholar]
- Yamaki, K.; Harada, I.; Goto, M.; Cho, C.S.; Akaike, T. Regulation of cellular morphology using temperature-responsive hydrogel for integrin-mediated mechanical force stimulation. Biomaterials 2009, 30, 1421–1427. [Google Scholar]
- Wang, J.; Heo, J.; Hua, S.Z. Spatially resolved shear distribution in microfluidic chip for studying force transduction mechanisms in cells. Lab Chip 2010, 10, 235–239. [Google Scholar]
- Sawada, Y.; Nakamura, K.; Doi, K.; Takeda, K.; Tobiume, K.; Saitoh, M.; Morita, K.; Komuro, I.; de Vos, K.; Sheetz, M.; et al. Rap1 is involved in cell stretching modulation of p38 but not ERK or JNK MAP kinase. J. Cell Sci 2001, 114, 1221–1227. [Google Scholar]
- Yonemura, S.; Wada, Y.; Watanabe, T.; Nagafuchi, A.; Shibata, M. Alpha-catenin as a tension transducer that induces adherens junction development. Nat. Cell Biol 2010, 12, 533–542. [Google Scholar]
- ImageJ, version 1.44c; National Institute of Health: Bethesda, MD, USA, 2010.







© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Harada, I.; Yanagisawa, S.; Iwasaki, K.; Cho, C.-S.; Akaike, T. Local Mechanical Stimulation of Mardin-Darby Canine Kidney Cell Sheets on Temperature-Responsive Hydrogel. Int. J. Mol. Sci. 2012, 13, 1095-1108. https://doi.org/10.3390/ijms13011095
Harada I, Yanagisawa S, Iwasaki K, Cho C-S, Akaike T. Local Mechanical Stimulation of Mardin-Darby Canine Kidney Cell Sheets on Temperature-Responsive Hydrogel. International Journal of Molecular Sciences. 2012; 13(1):1095-1108. https://doi.org/10.3390/ijms13011095
Chicago/Turabian StyleHarada, Ichiro, Shunpei Yanagisawa, Katsuhiko Iwasaki, Chong-Su Cho, and Toshihiro Akaike. 2012. "Local Mechanical Stimulation of Mardin-Darby Canine Kidney Cell Sheets on Temperature-Responsive Hydrogel" International Journal of Molecular Sciences 13, no. 1: 1095-1108. https://doi.org/10.3390/ijms13011095
APA StyleHarada, I., Yanagisawa, S., Iwasaki, K., Cho, C. -S., & Akaike, T. (2012). Local Mechanical Stimulation of Mardin-Darby Canine Kidney Cell Sheets on Temperature-Responsive Hydrogel. International Journal of Molecular Sciences, 13(1), 1095-1108. https://doi.org/10.3390/ijms13011095