A Review of Molecular Mechanisms of the Anti-Leukemic Effects of Phenolic Compounds in Honey

Abstract
:1. Background
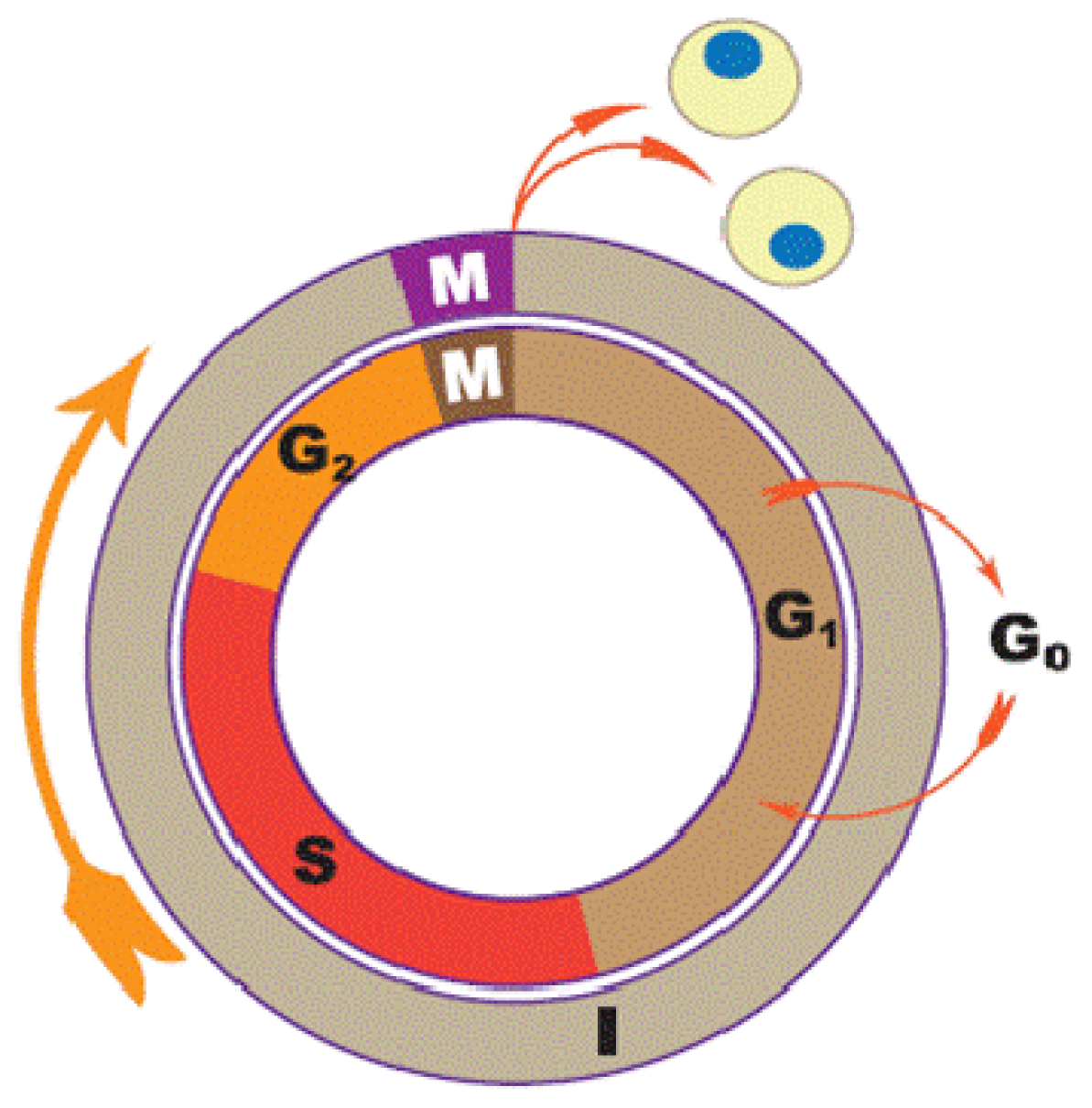
1.1. Cell Cycle
1.2. Apoptosis
1.3. Honey
1.4. Major Phenolic Compounds Present in Honey
2. Molecular Mechanisms of Anti-Leukemic Activities of Individual Polyphenols in Honey on Different Cell Lines
2.1. Quercetin
2.2. Kaempherol
2.3. Galangin
2.4. Apigenin
2.5. Acacetin
2.6. Fisetin, Myricetin and Wogonin
2.7. Caffeic Acid Phenylethyl Ester (CAPE)
2.8. Chrysin
2.9. Luteolin
2.10. Hesperidin
2.11. Dicaffeoylquinic Acids and Caffeoylquinic Acid Derivatives
2.12. Genkwanin
2.13. Rosmarinic Acid and Derivatives
2.14. Ellagic Acid (EA)
2.15. Other Phenolic Compounds
3. Conclusions
Acknowledgments
References
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2008, 127, 2893–2917. [Google Scholar]
- Smith, A.; Howell, D.; Patmore, R.; Jack, A.; Roman, E. Incidence of haematological malignancy by sub-type: A report from the haematological malignancy research network. Br. J. Cancer 2011, 105, 1684–1692. [Google Scholar]
- Fresco, P.; Borges, F.; Diniz, C.; Marques, M. New insights on the anticancer properties of dietary polyphenols. Med. Res. Rev 2006, 26, 747–766. [Google Scholar]
- Morse, M.A.; Stoner, G.D. Cancer chemoprevention: Principles and prospects. Carcinog. Oxf 1993, 14, 1737. [Google Scholar]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr 2000, 130, 2073–2085. [Google Scholar]
- Bogdanov, S. The Book of Honey. Available online: http://www.bee-hexagon.net/en/protected-sid-UHJvcG9saXNCb29rMi5wZGY%3D.htm accessed on 26 June 2012.
- Gómez-Caravaca, A.M.; Gómez-Romero, M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Advances in the analysis of phenolic compounds in products derived from bees. J. Pharm. Biomed. Anal 2006, 41, 1220–1234. [Google Scholar]
- Viuda-Martos, M.; Ruiz-Navajas, Y.; Fernández-López, J.; Pérez-Álvarez, J.A. Functional properties of honey, propolis, and royal jelly. J. Food Sci 2008, 73, R117–R124. [Google Scholar]
- Erejuwa, O.; Sulaiman, S.; Wahab, M.; Sirajudeen, K.; Salleh, M.; Gurtu, S. Antioxidant protection of Malaysian tualang honey in pancreas of normal and streptozotocin-induced diabetic rats. Ann. Endocrinol 2010, 71, 291–296. [Google Scholar]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Salam, S.K.N.; Salleh, M.S.M.; Gurtu, S. Comparison of antioxidant effects of honey, glibenclamide, metformin, and their combinations in the kidneys of streptozotocin-induced diabetic rats. Int. J. Mol. Sci 2011, 12, 829–843. [Google Scholar]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Antioxidant protective effect of glibenclamide and metformin in combination with honey in pancreas of streptozotocin-induced diabetic rats. Int. J. Mol. Sci 2011, 11, 2056–2066. [Google Scholar]
- Jaganathan, S.K.; Mandal, M. Antiproliferative effects of honey and of its polyphenols: A review. J. Biomed. Biotechnol. 2009. [Google Scholar] [CrossRef]
- Le Marchand, L. Cancer preventive effects of flavonoids—A review. Biomed. Pharmacother 2002, 56, 296–301. [Google Scholar]
- Middleton, E., Jr; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Ren, W.; Qiao, Z.; Wang, H.; Zhu, L.; Zhang, L. Flavonoids: Promising anticancer agents. Med. Res. Rev 2003, 23, 519–534. [Google Scholar]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Differential responses to blood pressure and oxidative stress in streptozotocin-induced diabetic wistar-Kyoto rats and spontaneously hypertensive rats: Effects of antioxidant (honey) treatment. Int. J. Mol. Sci 2011, 12, 1888–1907. [Google Scholar]
- Kelkel, M.; Jacob, C.; Dicato, M.; Diederich, M. Potential of the dietary antioxidants resveratrol and curcumin in prevention and treatment of hematologic malignancies. Molecules 2010, 15, 7035–7074. [Google Scholar]
- Meeran, S.M.; Katiyar, S.K. Cell cycle control as a basis for cancer chemoprevention through dietary agents. Front. Biosci 2008, 13, 2191–2202. [Google Scholar]
- Schwartz, G.K.; Shah, M.A. Targeting the cell cycle: A new approach to cancer therapy. J. Clin. Oncol 2005, 23, 9408–9421. [Google Scholar]
- Tyson, J.J.; Csikasz-Nagy, A.; Novak, B. The dynamics of cell cycle regulation. Bioessays 2002, 24, 1095–1109. [Google Scholar]
- Carnero, A. Targeting the cell cycle for cancer therapy. Br. J. Cancer 2002, 87, 129–133. [Google Scholar]
- Nurse, P. A long twentieth century of review the cell cycle and beyond. Cell 2000, 100, 71–78. [Google Scholar]
- Beier, R.; Bürgin, A.; Kiermaier, A.; Fero, M.; Karsunky, H.; Saffrich, R.; Möröy, T.; Ansorge, W.; Roberts, J.; Eilers, M. Induction of cyclin E–cdk2 kinase activity, E2F-dependent transcription and cell growth by Myc are genetically separable events. EMBO J 2000, 19, 5813–5823. [Google Scholar]
- Johnston, K.A.; Polymenis, M.; Wang, S.; Branda, J.; Schmidt, E.V. Novel regulatory factors interacting with the promoter of the gene encoding the mRNA cap binding protein (eIF4E) and their function in growth regulation. Mol. Cell. Biol 1998, 18, 5621–5633. [Google Scholar]
- Kiechle, F.L.; Zhang, X. Apoptosis: Biochemical aspects and clinical implications. Clin. Chim. Acta 2002, 326, 27–45. [Google Scholar]
- Rich, T.; Allen, R.L.; Wyllie, A.H. Defying death after DNA damage. Nature 2000, 407, 777–783. [Google Scholar]
- Debatin, K.M. Apoptosis pathways in cancer and cancer therapy. Cancer Immunol. Immunother 2004, 53, 153–159. [Google Scholar]
- Schuler, M.; Green, D. Mechanisms of p53-dependent apoptosis. Biochem. Soc. Trans 2001, 29, 684–688. [Google Scholar]
- Schuler, M.; Bossy-Wetzel, E.; Goldstein, J.C.; Fitzgerald, P.; Green, D.R. P53 induces apoptosis by caspase activation through mitochondrial cytochrome c release. J. Biol. Chem 2000, 275, 7337–7342. [Google Scholar]
- Hengst, L.; Göpfert, U.; Lashuel, H.A.; Reed, S.I. Complete inhibition of Cdk/cyclin by one molecule of p21Cip1. Genes Dev 1998, 12, 3882–3888. [Google Scholar]
- Bogdanov, S.; Martin, P. Honey authenticity: A review. Mitt. Lebensm. Hyg 2002, 93, 232–254. [Google Scholar]
- Bogdanov, S. Honey Composition. The Honey Book. Available online: http://www.bee-hexagon.net/en/protected-sid-NUNvbXBvc2l0aW9uSG9uZXkucGRm.htm accessed on 28 February 2012.
- Gheldof, N.; Wang, X.H.; Engeseth, N.J. Identification and quantification of antioxidant components of honeys from various floral sources. J. Agric. Food Chem 2002, 50, 5870–5877. [Google Scholar]
- Haydak, M.; Palmer, L.; Tanquary, M.; Vivino, A. Vitamin content of honeys. J. Nutr 1942, 23, 581. [Google Scholar]
- Kishore, R.K.; Halim, A.S.; Syazana, M.; Sirajudeen, K. Tualang honey has higher phenolic content and greater radical scavenging activity compared with other honey sources. Nutr. Res 2011, 4, 322–325. [Google Scholar]
- Mohamed, M.; Sirajudeen, K.; Swamy, M.; Yaacob, N.S.; Sulaiman, S.A. Studies on the antioxidant properties of Tualang honey of Malaysia. Afr. J. Tradit. Complement. Altern. Med 2010, 7, 59–63. [Google Scholar]
- Perez, R.A.; Iglesias, M.T.; Pueyo, E.; Gonzalez, M.; De Lorenzo, C. Amino acid composition and antioxidant capacity of Spanish honeys. J. Agric. Food Chem 2007, 55, 360–365. [Google Scholar]
- White, J.; Doner, L.W. Honey composition and properties. Beekeep. USA 1980, 335, 82–91. [Google Scholar]
- Fauzi, A.N.; Norazmi, M.N.; Yaacob, N.S. Tualang honey induces apoptosis and disrupts the mitochondrial membrane potential of human breast and cervical cancer cell lines. Food Chem. Toxicol 2011, 49, 871–878. [Google Scholar]
- Ghashm, A.A.; Othman, N.H.; Khattak, M.N.; Ismail, N.M.; Saini, R. Antiproliferative effect of Tualang honey on oral squamous cell carcinoma and osteosarcoma cell lines. BMC Complement. Altern. Med 2010, 10, 49. [Google Scholar]
- Gribel, N.; PashinskiÄ, V. The antitumor properties of honey. Vopr. Onkol 1990, 36, 704–709. [Google Scholar]
- Orsolić, N.; Knezević, A.; Sver, L.; Terzić, S.; Hackenberger, B.K.; Basić, I. Influence of honey bee products on transplantable murine tumours. Vet. Comp. Oncol 2003, 1, 216–226. [Google Scholar]
- Swellam, T.; Miyanaga, N.; Onozawa, M.; Hattori, K.; Kawai, K.; Shimazui, T.; Akaza, H. Antineoplastic activity of honey in an experimental bladder cancer implantation model: In vivo and in vitro studies. Int. J. Urol 2003, 10, 213–219. [Google Scholar]
- Wen, C.T.P.; Hussein, S.Z.; Abdullah, S.; Karim, N.A.; Makpol, S.; Yusof, Y.A.M. Gelam and nenas honeys inhibit proliferation of HT 29 colon cancer cells by inducing DNA damage and apoptosis while suppressing inflammation. Asian Pac. J. Cancer Prev 2012, 13, 1605–1610. [Google Scholar]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci 2004, 74, 2157–2184. [Google Scholar]
- Ferreres, F.; Tomáas-Barberáan, F.A.; Gil, M.A.I.; Tomáas-Lorente, F. An HPLC technique for flavonoid analysis in honey. J. Sci. Food Agric 1991, 56, 49–56. [Google Scholar]
- Yao, L.; Jiang, Y.; Singanusong, R.; Datta, N.; Raymont, K. Phenolic acids and abscisic acid in Australian Eucalyptus honeys and their potential for floral authentication. Food Chem 2004, 86, 169–177. [Google Scholar]
- Yaoa, L.; Jiang, Y.; Singanusong, R.; Datta, N.; Raymont, K. Phenolic acids in Australian Melaleuca, Guioa, Lophostemon, Banksia and Helianthus honeys and their potential for floral authentication. Food Res. Int 2005, 38, 651–658. [Google Scholar]
- Andrade, P.; Ferreres, F.; Gil, M.I.; Tomás-Barberán, F.A. Determination of phenolic compounds in honeys with different floral origin by capillary zone electrophoresis. Food Chem 1997, 60, 79–84. [Google Scholar]
- Robinson, J.P.; Narayanan, P.K.; Carter, W.O. Functional measurements using HL-60 cells. Methods Cell Biol 1994, 42, 423–436. [Google Scholar]
- Trayner, I.D.; Bustorff, T.; Etches, A.E.; Mufti, G.J.; Foss, Y.; Farzaneh, F. Changes in antigen expression on differentiating HL60 cells treated with dimethylsulphoxide, all-trans retinoic acid, α1,25-dihydroxyvitamin D3 or 12-O-tetradecanoyl phorbol-13-acetate. Leuk. Res 1998, 22, 537–547. [Google Scholar]
- O’Gorman, D.M.; McKenna, S.L.; McGahon, A.J.; Knox, K.A.; Cotter, T.G. Sensitisation of HL60 human leukaemic cells to cytotoxic drug-induced apoptosis by inhibition of PI3-kinase survival signals. Leukemia 2000, 14, 602–611. [Google Scholar]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F.; Gallo, R. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. Blood 1979, 54, 713–733. [Google Scholar]
- Klein, E.; Vánky, F.; Ben-Bassat, H.; Neumann, H.; Ralph, P.; Zeuthen, J.; Polliack, A. Properties of the K562 cell line, derived from a patient with chronic myeloid leukemia. Int. J. Cancer 1976, 18, 421–431. [Google Scholar]
- Harris, P.; Ralph, P. Human leukemic models of myelomonocytic development: A review of the HL-60 and U937 cell lines. J. Leukoc. Biol 1985, 37, 407–422. [Google Scholar]
- Hurwitz, R.; Hozier, J.; Lebien, T.; Minowada, J.; Gajl-Peczalska, K.; Kubonishi, I.; Kersey, J. Characterization of a leukemic cell line of the pre-B phenotype. Int. J. Cancer 1979, 23, 174–180. [Google Scholar]
- Broggini, M.; Marchini, S.; Galliera, E.; Borsotti, P.; Taraboletti, G.; Erba, E.; Sironi, M.; Jimeno, J.; Faircloth, G.; Giavazzi, R. Aplidine, a new anticancer agent of marine origin, inhibits vascular endothelial growth factor (VEGF) secretion and blocks VEGF-VEGFR-1 (flt-1) autocrine loop in human leukemia cells MOLT-4. Leukemia 2003, 17, 52–59. [Google Scholar]
- Goto, H.; Yanagimachi, M.; Goto, S.; Takeuchi, M.; Kato, H.; Yokosuka, T.; Kajiwara, R.; Yokota, S. Methylated chrysin reduced cell proliferation, but antagonized cytotoxicity of other anticancer drugs in acute lymphoblastic leukemia. Anti-Cancer Drugs 2012, 23, 417–425. [Google Scholar]
- Goto, H.; Yanagimachi, M.; Kajiwara, R.; Kuroki, F.; Yokota, S. Lack of mitochondrial depolarization by oxidative stress is associated with resistance to buthionine sulfoximine in acute lymphoblastic leukemia cells. Leuk. Res 2007, 31, 1293–1301. [Google Scholar]
- Kang, T.; Liang, N. Studies on the inhibitory effects of quercetin on the growth of HL-60 leukemia cells. Biochem. Pharmacol 1997, 54, 1013–1018. [Google Scholar]
- Csokay, B.; Prajda, N.; Weber, G.; Olah, E. Molecular mechanisms in the antiproliferative action of quercetin. Life Sci 1997, 60, 2157–2163. [Google Scholar]
- Lee, T.J.; Kim, O.H.; Kim, Y.H.; Lim, J.H.; Kim, S.; Park, J.W.; Kwon, T.K. Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Cancer Lett 2006, 240, 234–242. [Google Scholar]
- Spagnuolo, C.; Russo, M.; Bilotto, S.; Tedesco, I.; Laratta, B.; Russo, G.L. Dietary polyphenols in cancer prevention: The example of the flavonoid quercetin in leukemia. Ann. NY Acad. Sci 2012, 1259, 95–103. [Google Scholar]
- Bestwick, C.S.; Milne, L.; Duthie, S.J. Kaempferol induced inhibition of HL-60 cell growth results from a heterogeneous response, dominated by cell cycle alterations. Chem. Biol. Interact 2007, 170, 76–85. [Google Scholar]
- Bestwick, C.S.; Milne, L. Influence of galangin on HL-60 cell proliferation and survival. Cancer Lett 2006, 243, 80–89. [Google Scholar]
- Wang, I.K.; Lin-Shiau, S.Y.; Lin, J.K. Induction of apoptosis by apigenin and related flavonoids through cytochrome c release and activation of caspase-9 and caspase-3 in leukaemia HL-60 cells. Eur. J. Cancer 1999, 35, 1517–1525. [Google Scholar]
- Gonzalez-Mejia, M.; Voss, O.; Murnan, E.; Doseff, A. Apigenin-Induced apoptosis of leukemia cells is mediated by a bimodal and differentially regulated residue-specific phosphorylation of heat-shock protein-27. Cell Death Dis 2010, 1, e64. [Google Scholar]
- Budhraja, A.; Gao, N.; Zhang, Z.; Son, Y.O.; Cheng, S.; Wang, X.; Ding, S.; Hitron, A.; Chen, G.; Luo, J. Apigenin induces apoptosis in human leukemia cells and exhibits anti-leukemic activity in vivo. Mol. Cancer Ther 2012, 11, 132–142. [Google Scholar]
- Watanabe, K.; Kanno, S.; Tomizawa, A.; Yomogida, S.; Ishikawa, M. Acacetin induces apoptosis in human T cell leukemia Jurkat cells via activation of a caspase cascade. Oncol. Rep 2012, 27, 204–209. [Google Scholar]
- Lee, W.-R.; Shen, S.-C.; Lin, H.-Y.; Hou, W.-C.; Yang, L.-L.; Chen, Y.-C. Wogonin and fisetin induce apoptosis in human promyeloleukemic cells, accompanied by a decrease of reactive oxygen species, and activation of caspase 3 and Ca2+-dependent endonuclease. Biochem. Pharmacol 2002, 63, 225–236. [Google Scholar]
- Ding, J.; Polier, G.; Köhler, R.; Giaisi, M.; Krammer, P.H.; Li-Weber, M. Wogonin and related natural flavones overcome tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) protein resistance of tumors by down-regulation of c-FLIP protein and up-regulation of TRAIL receptor 2 expression. J. Biol. Chem 2012, 287, 641–649. [Google Scholar]
- Chen, J.H.; Shao, Y.; Huang, M.T.; Chin, C.K.; Ho, C.T. Inhibitory effect of caffeic acid phenethyl ester on human leukemia HL-60 cells. Cancer Lett 1996, 108, 211–214. [Google Scholar]
- Chen, Y.J.; Shiao, M.S.; Hsu, M.L.; Tsai, T.H.; Wang, S.Y. Effect of caffeic acid phenethyl ester, an antioxidant from propolis, on inducing apoptosis in human leukemic HL-60 cells. J. Agric. Food Chem 2001, 49, 5615–5619. [Google Scholar]
- Jin, U.H.; Song, K.H.; Motomura, M.; Suzuki, I.; Gu, Y.H.; Kang, Y.J.; Moon, T.C.; Kim, C.H. Caffeic acid phenethyl ester induces mitochondria-mediated apoptosis in human myeloid leukemia U937 cells. Mol. Cell. Biochem 2008, 310, 43–48. [Google Scholar]
- Woo, K.J.; Jeong, Y.J.; Park, J.W.; Kwon, T.K. Chrysin-Induced apoptosis is mediated through caspase activation and Akt inactivation in U937 leukemia cells. Biochem. Biophys. Res. Commun 2004, 325, 1215–1222. [Google Scholar]
- Wudtiwai, B.; Sripanidkulchai, B.; Kongtawelert, P.; Banjerdpongchai, R. Methoxyflavone derivatives modulate the effect of TRAIL-induced apoptosis in human leukemic cell lines. J. Hematol. Oncol 2011, 4, 52. [Google Scholar]
- Ko, W.; Kang, T.; Lee, S.; Kim, Y.; Lee, B. Effects of luteolin on the inhibition of proliferation and induction of apoptosis in human myeloid leukaemia cells. Phytother. Res 2002, 16, 295–298. [Google Scholar]
- Cheng, A.C.; Huang, T.C.; Lai, C.S.; Pan, M.H. Induction of apoptosis by luteolin through cleavage of Bcl-2 family in human leukemia HL-60 cells. Eur. J. Pharmacol 2005, 509, 1–10. [Google Scholar]
- Ghorbani, A.; Nazari, M.; Jeddi-Tehrani, M.; Zand, H. The citrus flavonoid hesperidin induces p53 and inhibits NF-κB activation in order to trigger apoptosis in NALM-6 cells: Involvement of PPARγ-dependent mechanism. Eur. J. Nutr 2012, 51, 1–8. [Google Scholar]
- Mishima, S.; Inoh, Y.; Narita, Y.; Ohta, S.; Sakamoto, T.; Araki, Y.; Suzuki, K.M.; Akao, Y.; Nozawa, Y. Identification of caffeoylquinic acid derivatives from Brazilian propolis as constituents involved in induction of granulocytic differentiation of HL-60 cells. Bioorgan. Med. Chem 2005, 13, 5814–5818. [Google Scholar]
- Sghaier, M.B.; Skandrani, I.; Nasr, N.; Franca, M.G.D.; Chekir-Ghedira, L.; Ghedira, K. Flavonoids and sesquiterpenes from Tecurium ramosissimum promote antiproliferation of human cancer cells and enhance antioxidant activity: A structure-activity relationship study. Environ. Toxicol. Pharmacol 2011, 32, 336–348. [Google Scholar]
- Valdés, A.; Simó, C.; Ibáñez, C.; Rocamora-Reverte, L.; Ferragut, J.A.; García-Cañas, V.; Cifuentes, A. Effect of dietary polyphenols on K562 leukemia cells: A foodomics approach. Electrophoresis 2012, 33, 2314–2327. [Google Scholar]
- Hagiwara, Y.; Kasukabe, T.; Kaneko, Y.; Niitsu, N.; Okabe-Kado, J. Ellagic acid, a natural polyphenolic compound, induces apoptosis and potentiates retinoic acid-induced differentiation of human leukemia HL-60 cells. Int. J. Hematol 2010, 92, 136–143. [Google Scholar]
- Del Pozo-Insfran, D.; Percival, S.S.; Talcott, S.T. Açai polyphenolics in their glycoside and aglycone forms induce apoptosis of HL-60 leukemia cells. J. Agric. Food Chem 2006, 54, 1222–1229. [Google Scholar]
- Ninomiya, M.; Nishida, K.; Tanaka, K.; Watanabe, K.; Koketsu, M. Structure-Activity relationship studies of 5,7-dihydroxyflavones as naturally occurring inhibitors of cell proliferation in human leukemia HL-60 cells. J. Natur. Med. 2012. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Category | Compound |
|---|---|
| Flavonoles | Quercetin, kaempferol, Galangin, Fisetin, Myricetin |
| Flavanones | Pinocembrin, Naringin, Naringenin, Hesperidin Pinobanksin |
| Flavones | Apigenin, Acacetin, Chrysin, Luteolin Genkwanin, wogonin, tricetin |
| Phenolic acids | Caffeic acid, chlorogenic acid, cinnamic acid, p-coumaric acid, vanillic acid, ferulic acid, p-hydroxybenzoic acid, gallic acid, syringic acid abscisic acid *, rosmarinic acid and derivatives |
| Coumarins | Coumarin |
| Tannins | Ellagic acid |
| Phenolic compound | Cell line | Mechanism of action (s) |
|---|---|---|
| Quercetin | HL-60 | Cell cycle arrest, inhibition of growth and proliferation |
| K562 | Induction of apoptosis and differentiation | |
| U937 | Induction of apoptosis cell cycle arrest | |
| Kaempferol | HL-60 | Cell cycle arrest and growth inhibition |
| Galangin | HL-60 | Antiproliferation and induction of apoptosis |
| Apigenin | HL-60 | Induction of apoptosis |
| THP | Induction of apoptosis | |
| U937 | Induction of apoptosis | |
| Acacetin | Jurkat cells | Growth inhibition and induction of apoptosis |
| Fisetin, myricetin and wogonin | HL-60 | Induction of apoptosis |
| CAPE | HL-60 | Growth inhibition and induction of apoptosis |
| U937 | Induction of apoptosis | |
| Chrysin | U937 | Induction of apoptosis |
| YCUB series | Growth inhibition and induction of apoptosis | |
| Luteolin | HL-60 | Growth inhibition and induction of apoptosis |
| Hesperidin | NALM-6 | Growth inhibition and induction of apoptosis |
| Dicaffeoylquinic acids and caffeoylquinic acid derivatives | HL-60 | Growth inhibition, induction of apoptosis and Differentiation |
| Genkwanin | K562 | Cytotoxicity |
| Rosmarinic acid and derivatives | K562 | Growth inhibition and induction of apoptosis |
| Other phenolic acids | HL-60 | Growth inhibition and induction of apoptosis |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Abubakar, M.B.; Abdullah, W.Z.; Sulaiman, S.A.; Suen, A.B. A Review of Molecular Mechanisms of the Anti-Leukemic Effects of Phenolic Compounds in Honey. Int. J. Mol. Sci. 2012, 13, 15054-15073. https://doi.org/10.3390/ijms131115054
Abubakar MB, Abdullah WZ, Sulaiman SA, Suen AB. A Review of Molecular Mechanisms of the Anti-Leukemic Effects of Phenolic Compounds in Honey. International Journal of Molecular Sciences. 2012; 13(11):15054-15073. https://doi.org/10.3390/ijms131115054
Chicago/Turabian StyleAbubakar, Murtala B., Wan Zaidah Abdullah, Siti Amrah Sulaiman, and Ang Boon Suen. 2012. "A Review of Molecular Mechanisms of the Anti-Leukemic Effects of Phenolic Compounds in Honey" International Journal of Molecular Sciences 13, no. 11: 15054-15073. https://doi.org/10.3390/ijms131115054
APA StyleAbubakar, M. B., Abdullah, W. Z., Sulaiman, S. A., & Suen, A. B. (2012). A Review of Molecular Mechanisms of the Anti-Leukemic Effects of Phenolic Compounds in Honey. International Journal of Molecular Sciences, 13(11), 15054-15073. https://doi.org/10.3390/ijms131115054