Dextran and Polymer Polyethylene Glycol (PEG) Coating Reduce Both 5 and 30 nm Iron Oxide Nanoparticle Cytotoxicity in 2D and 3D Cell Culture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
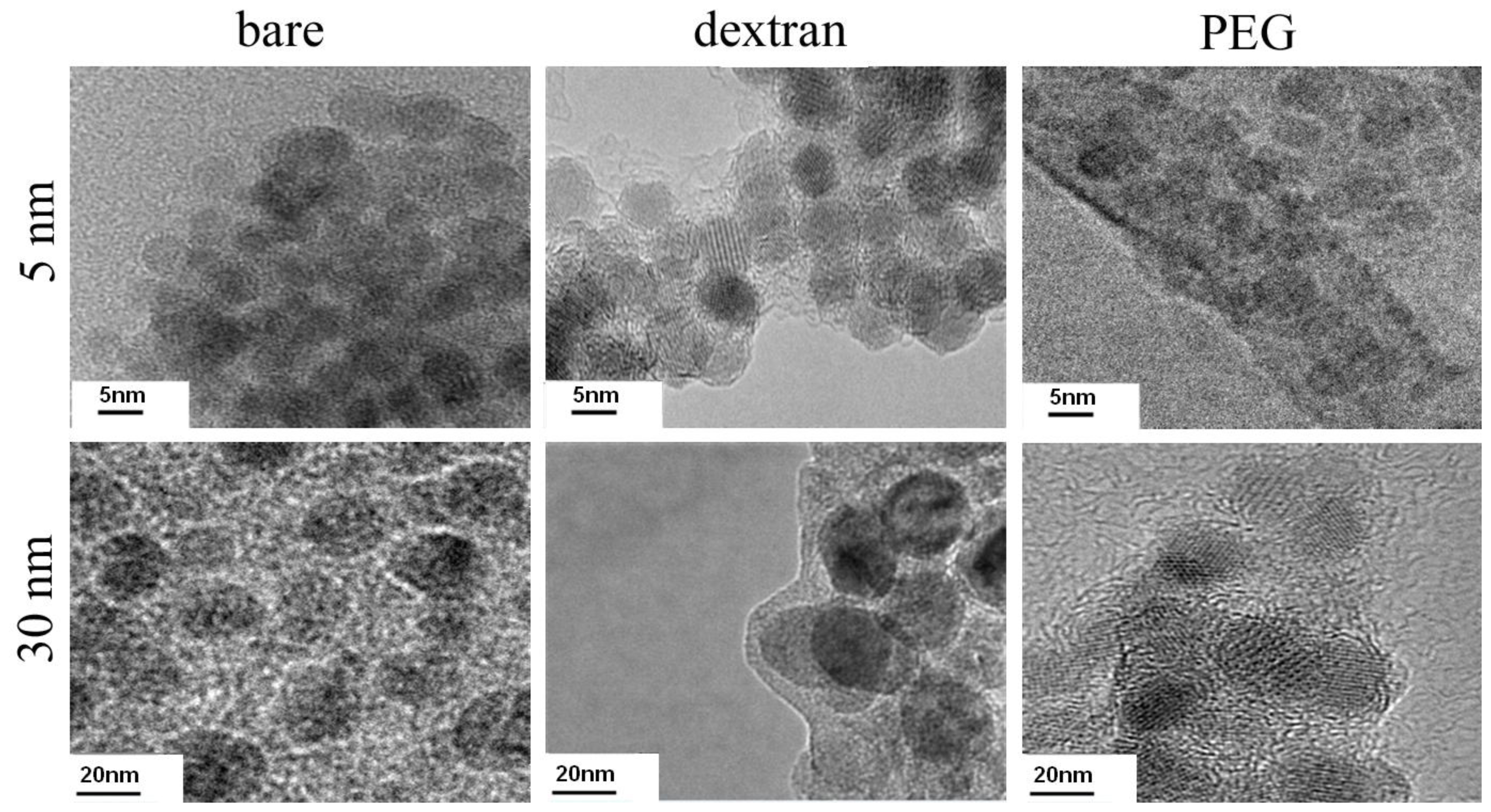
2.1. Nanoparticle Coating
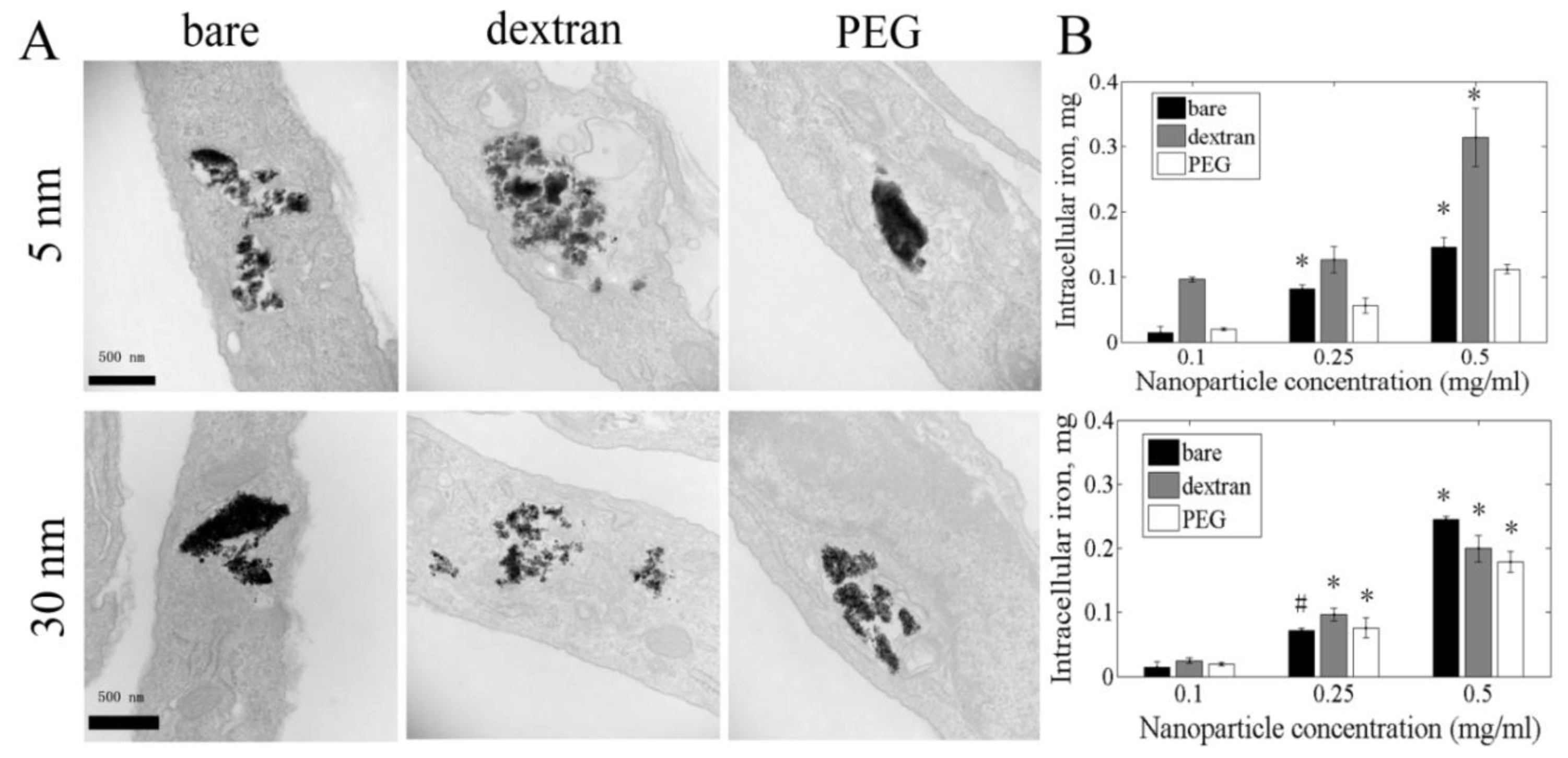
2.2. Cell Uptake of Iron Oxide Nanoparticles
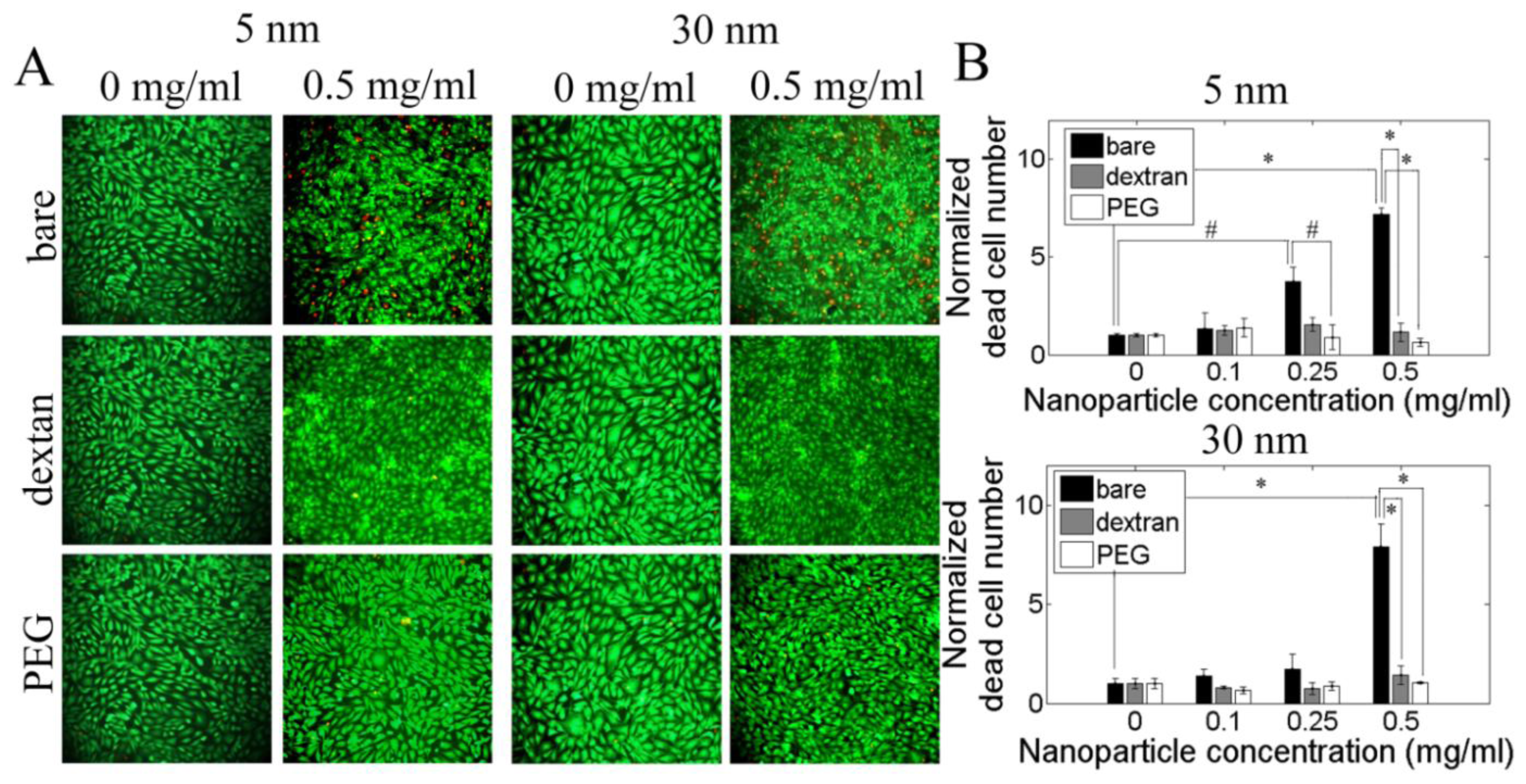
2.3. Cytotoxicity
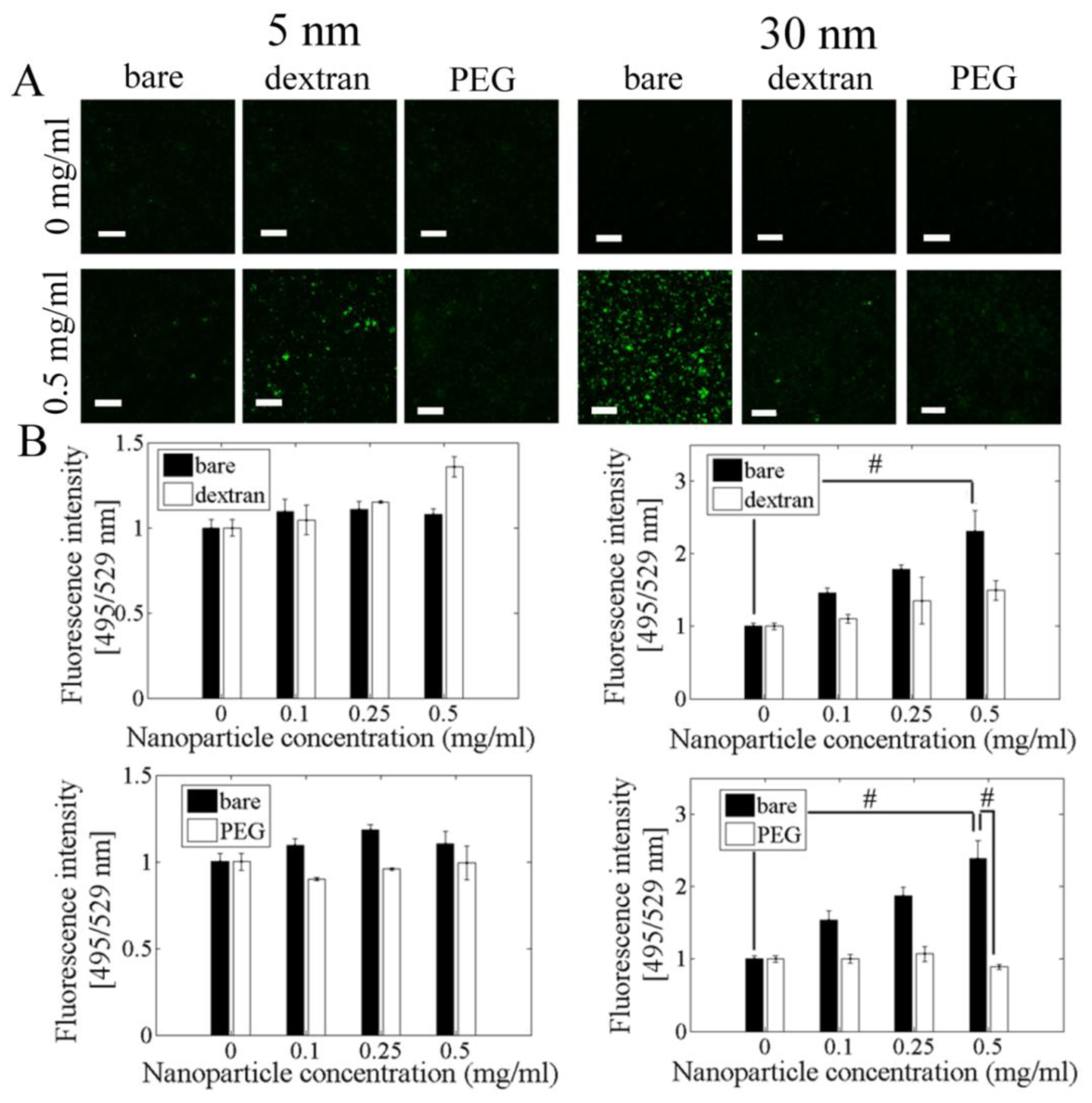
2.4. ROS Formation
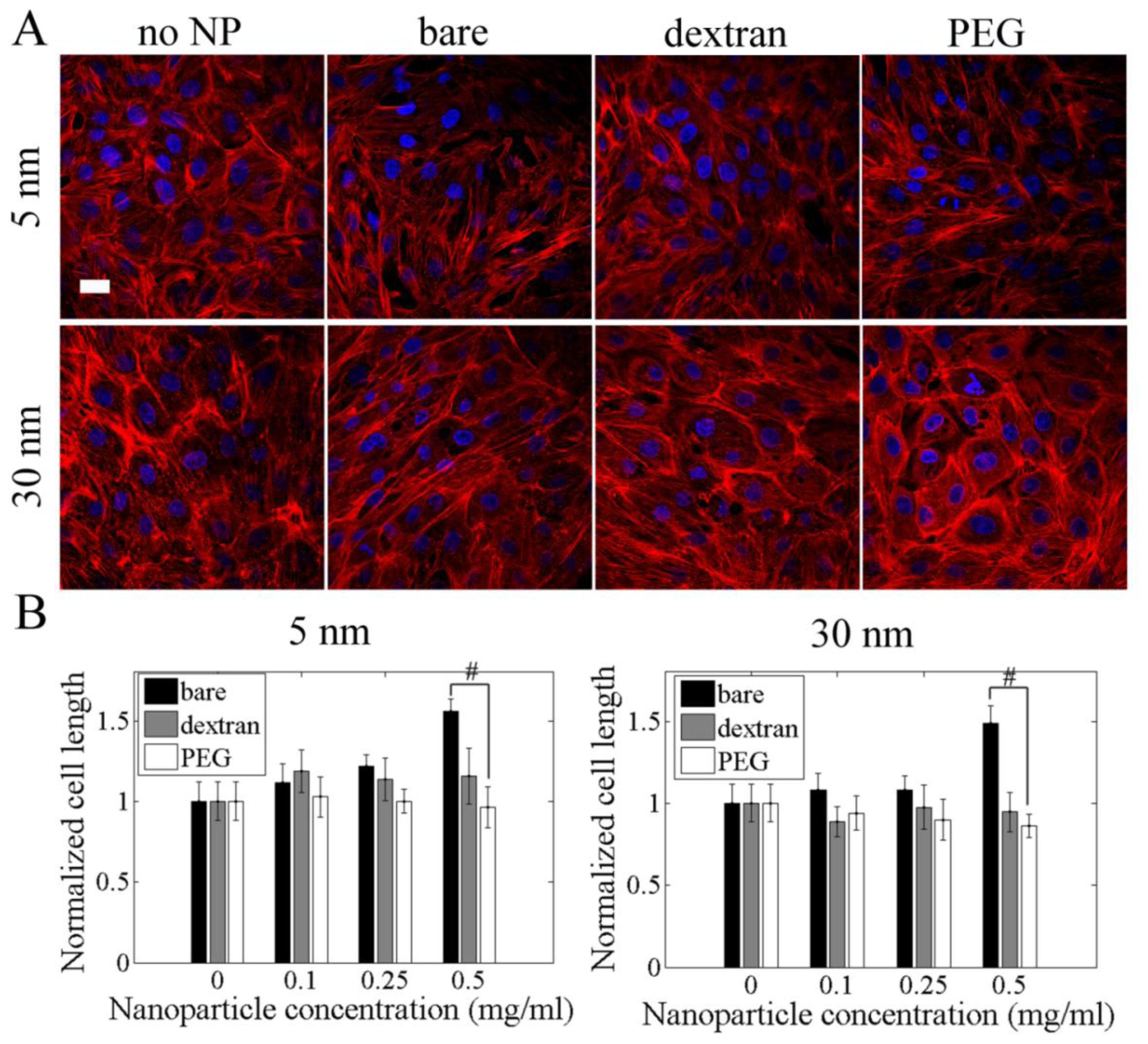
2.5. Cell Length and Actin Cytoskeleton
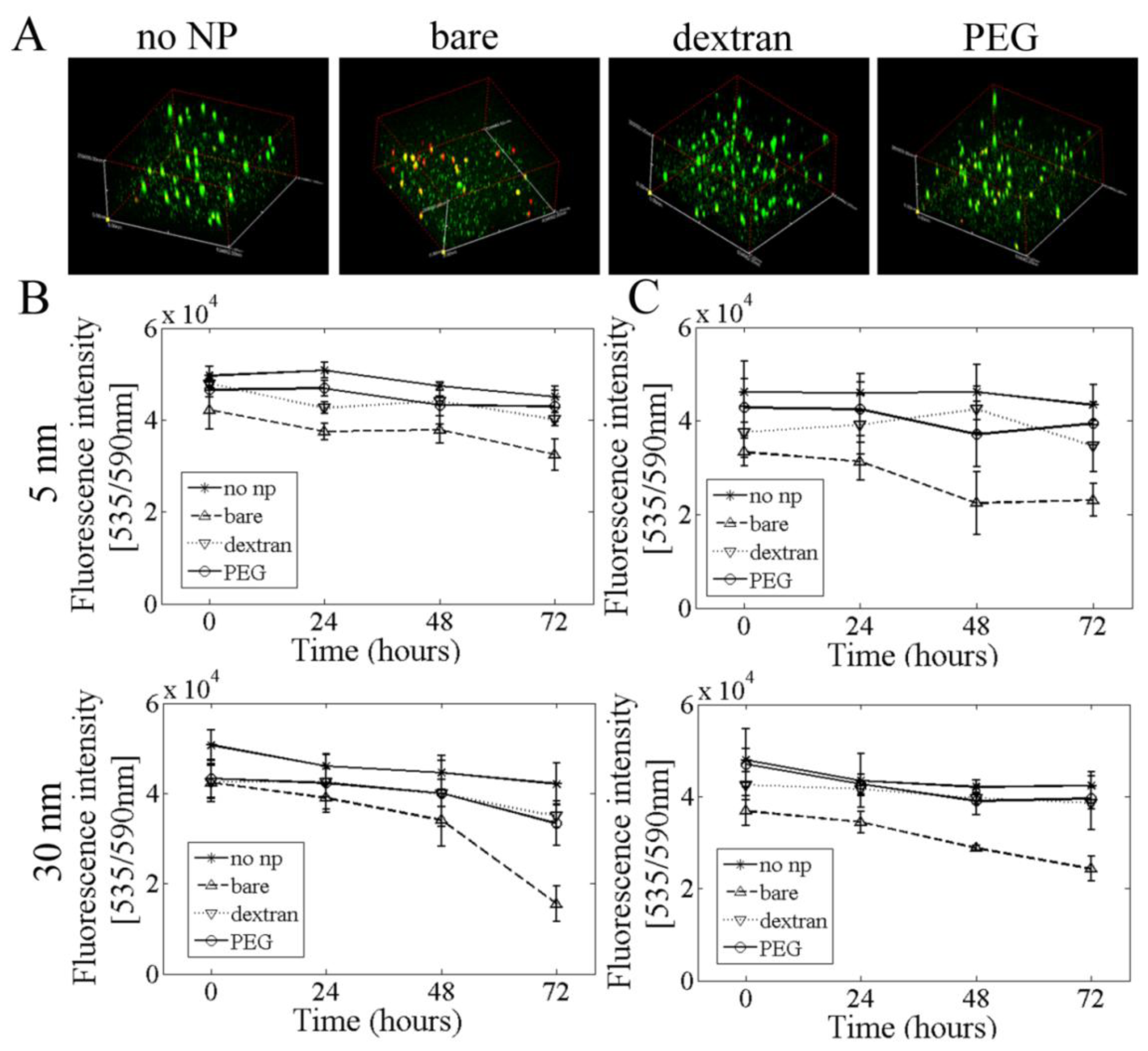
2.6. 3D Cell Culture
2.7. Discussion
3. Experimental Section
3.1. Cell Culture
3.2. Nanoparticle Coating
3.3. Nanoparticle Coating and Cell Uptake by Transmission Electron Microscopy (TEM)
3.4. Cell Viability
3.5. Reactive Oxygen Species (ROS)
3.6. Actin Cytoskeleton
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Chertok, B.; Moffat, B.A.; David, A.E.; Yu, F.; Bergemann, C.; Ross, B.D.; Yang, V.C. Iron oxide nanoparticles as a drug delivery vehicle for MRI monitored magnetic targeting of brain tumors. Biomaterials 2008, 29, 487–496. [Google Scholar]
- Pouliquen, D.; Le Jeune, J.J.; Perdrisot, R.; Ermias, A.; Jallet, P. Iron oxide nanoparticles for use as an MRI contrast agent: Pharmacokinetics and metabolism. Magn. Reson. Imag 1991, 9, 275–283. [Google Scholar]
- Fortin, J.-P.; Wilhelm, C.; Servais, J.; Ménager, C.; Bacri, J.-C.; Gazeau, F. Size-sorted anionic iron oxide nanomagnets as colloidal mediators for magnetic hyperthermia. J. Am. Chem. Soc 2007, 129, 2628–2635. [Google Scholar]
- Babič, M.; Horák, D.; Trchová, M.; Jendelová, P.; Glogarová, K.I.; Lesný, P.; Herynek, V.; Hájek, M.; Syková, E. Poly(l-lysine)-modified iron oxide nanoparticles for stem cell labeling. Bioconjugate Chem 2008, 19, 740–750. [Google Scholar]
- Tartaj, P.; Morales, M.D.P.; Veintemillas-Verdaguer, S.; González-Carreño, T.; Serna, C.J. The preparation of magnetic nanoparticles for applications in biomedicine. J. Phys. Appl. Phys 2003, 36, R182–R197. [Google Scholar]
- Davda, J.; Labhasetwar, V. Characterization of nanoparticle uptake by endothelial cells. Int. J. Pharm 2002, 233, 51–59. [Google Scholar]
- Buyukhatipoglu, K.; Miller, T.A.; Morss Clyne, A. Flame synthesis and in vitro biocompatibility assessment of superparamagnetic iron oxide nanoparticles: Cellular uptake, toxicity and proliferation studies. J. Nanosci. Nanotechnol 2009, 9, 6834–6843. [Google Scholar]
- Soenen, S.J.H.; de Cuyper, M. Assessing iron oxide nanoparticle toxicity in vitro: current status and future prospects. Nanomedicine 2010, 5, 1261–1275. [Google Scholar]
- Mahmoudi, M.; Simchi, A.; Imani, M.; Milani, A.S.; Stroeve, P. An in vitro study of bare and poly(ethylene glycol)-co-fumarate-coated superparamagnetic iron oxide nanoparticles: A new toxicity identification procedure. Nanotechnology 2009, 20. [Google Scholar] [CrossRef]
- Mahmoudi, M.; Laurent, S.; Shokrgozar, M.A.; Hosseinkhani, M. Toxicity evaluations of superparamagnetic iron oxide nanoparticles: Cell “vision” versus physicochemical properties of nanoparticles. ACS Nano 2011, 5, 7263–7276. [Google Scholar]
- Quinlan, T.R.; Marsh, J.P.; Janssen, Y.M.; Borm, P.A.; Mossman, B.T. Oxygen radicals and asbestos-mediated disease. Environ. Health Perspect 1994, 69, 261–274. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Wei, H.; Cai, Q.; Rahn, R.O. Inhibition of UV light- and Fenton Reaction-induced oxidative DNA damage by the soybcan isoflavone genistein. Carcinogenesis 1996, 17, 73–77. [Google Scholar]
- Wu, X.; Tan, Y.; Mao, H.; Zhang, M. Toxic effects of iron oxide nanoparticles on human umbilical vein endothelial cells. Int. J. Nanomedicine 2010, 5, 385–339. [Google Scholar]
- Buyukhatipoglu, K.; Clyne, A.M. Superparamagnetic iron oxide nanoparticles change endothelial cell morphology and mechanics via reactive oxygen species formation. J. Biomed. Mater. Res. Part A 2011, 96A, 186–195. [Google Scholar]
- Gourlay, C.W.; Carpp, L.N.; Timpson, P.; Winder, S.J.; Ayscough, K.R. A role for the actin cytoskeleton in cell death and aging in yeast. J. Cell Biol 2004, 164, 803–809. [Google Scholar]
- Lind, K.; Kresse, M.; Debus, N.P.; Müller, R.H. A novel formulation for superparamagnetic iron oxide (SPIO) particles enhancing MR lymphography: Comparison of physicochemical properties and the in vivo Behaviour. J. Drug Target 2002, 10, 221–230. [Google Scholar]
- Zhang, Y.; Kohler, N.; Zhang, M. Surface modification of superparamagnetic magnetite nanoparticles and their intracellular uptake. Biomaterials 2002, 23, 1553–1561. [Google Scholar]
- Weissleder, R.; Bogdanov, A.; Papisov, M. Drug targeting in magnetic resonance imaging. Magn. Reson. Q 1992, 8, 55–63. [Google Scholar]
- Moore, A.; Marecos, E.; Bogdanov, A.; Weissleder, R. Tumoral distribution of long-circulating dextran-coated iron oxide nanoparticles in a rodent model. Radiology 2000, 214, 568–574. [Google Scholar]
- Thorek, D.; Chen, A.; Czupryna, J.; Tsourkas, A. Superparamagnetic iron oxide nanoparticle probes for molecular imaging. Ann. Biomed. Eng 2006, 34, 23–38. [Google Scholar]
- Mahmoudi, M.; Simchi, A.; Imani, M.; Häfeli, U.O. Superparamagnetic iron oxide nanoparticles with rigid cross-linked polyethylene glycol fumarate coating for application in imaging and drug delivery. J Phys. Chem. C 2009, 113, 8124–8131. [Google Scholar]
- Larsen, E.K.U.; Nielsen, T.; Wittenborn, T.; Birkedal, H.; Vorup-Jensen, T.; Jakobsen, M.H.; Østergaard, L.; Horsman, M.R.; Besenbacher, F.; Howard, K.A.; Kjems, J. Size-dependent accumulation of pegylated silane-coated magnetic iron oxide nanoparticles in murine tumors. ACS Nano 2009, 3, 1947–1951. [Google Scholar]
- Chouly, C.; Pouliquen, D.; Lucet, I.; Jeune, J.J.; Jallet, P. Development of superparamagnetic nanoparticles for MRI: Effect of particle size, charge and surface nature on biodistribution. J. Microencapsul 1996, 13, 245–255. [Google Scholar]
- Matuszewski, L.; Persigehl, T.; Wall, A.; Schwindt, W.; Tombach, B.; Fobker, M.; Poremba, C.; Ebert, W.; Heindel, W.; Bremer, C. Cell tagging with clinically approved iron oxides: Feasibility and effect of lipofection, particle size, and surface coating on labeling efficiency. Radiology 2005, 235, 155–161. [Google Scholar]
- Yin, H.; Too, H.P.; Chow, G.M. The effects of particle size and surface coating on the cytotoxicity of nickel ferrite. Biomaterials 2005, 26, 5818–5826. [Google Scholar]
- Gorth, D.J.; Rand, D.M.; Webster, T.J. Silver nanoparticle toxicity in Drosophila: size does matter. Int. J. Nanomedicine 2011, 6, 343–350. [Google Scholar]
- Kim, T.-H.; Kim, M.; Park, H.-S.; Shin, U.S.; Gong, M.-S.; Kim, H.-W. Size-dependent cellular toxicity of silver nanoparticles. J. Biomed. Mater. Res. Part A 2012, 100A, 1033–1043. [Google Scholar]
- Gupta, A.K.; Gupta, M. Synthesis and surface engineering of iron oxide nanoparticles for biomedical applications. Biomaterials 2005, 26, 3995–4021. [Google Scholar]
- Bellin, M.-F.; Beigelman, C.; Precetti-Morel, S. Iron oxide-enhanced MR lymphography: initial experience. Eur. J. Radiol 2000, 34, 257–264. [Google Scholar]
- Hong, R.Y.; Feng, B.; Chen, L.L.; Liu, G.H.; Li, H.Z.; Zheng, Y.; Wei, D.G. Synthesis, characterization and MRI application of dextran-coated Fe3O4 magnetic nanoparticles. Biochem. Eng. J 2008, 42, 290–300. [Google Scholar]
- Huang, J.; Bu, L.; Xie, J.; Chen, K.; Cheng, Z.; Li, X.; Chen, X. Effects of nanoparticle size on cellular uptake and liver mri with polyvinylpyrrolidone-coated iron oxide nanoparticles. ACS Nano 2010, 4, 7151–7160. [Google Scholar]
- Limbach, L.K.; Li, Y.; Grass, R.N.; Brunner, T.J.; Hintermann, M.A.; Muller, M.; Gunther, D.; Stark, W.J. Oxide nanoparticle uptake in human lung fibroblasts: Effects of particle size, agglomeration, and diffusion at low concentrations. Environ. Sci. Technol 2005, 39, 9370–9376. [Google Scholar]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar]
- Frei, B. Reactive oxygen species and antioxidant vitamins: Mechanisms of action. Am. J. Med 1994, 97, S5–S13. [Google Scholar]
- Kehrer, J.; Smith, C. Free Radicals in Biology: Sources, Reactivities and Roles in the Etiology of Human Diseases. In Natural Antioxidants in Human Health and Disease; Academic Press: San Diego, CA USA, 1994; pp. 25–62. [Google Scholar]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar]
- Halliwell, B. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: Oxford; UK, 2007. [Google Scholar]
- Schmidt, A.; Hall, M.N. Signaling to the actin cytoskeleton. Ann. Rev. Cell Dev. Biol 1998, 14, 305–308. [Google Scholar]
- Lu, L.; Oswald, S.J.; Ngu, H.; Yin, F.C.P. Mechanical properties of actin stress fibers in living cells. Biophy. J 2008, 95, 6060–6071. [Google Scholar]
- Prasain, N.; Stevens, T. The actin cytoskeleton in endothelial cell phenotypes. Microvas. Res 2009, 77, 53–63. [Google Scholar]
- Ayscough, K. Actin regulation in endocytosis. J. Cell Sci 2006, 119, 4589–4598. [Google Scholar]
- Apopa, P.; Qian, Y.; Shao, R.; Guo, N.; Schwegler-Berry, D.; Pacurari, M.; Porter, D.; Shi, X.; Vallyathan, V.; Castranova, V.; Flynn, D. Iron oxide nanoparticles induce human microvascular endothelial cell permeability through reactive oxygen species production and microtubule remodeling. Part. Fibre Toxical 2009, 6. [Google Scholar] [CrossRef]
- Rodriguez, O.C.; Schaefer, A.W.; Mandato, C.A.; Forscher, P.; Bement, W.M.; Waterman-Storer, C.M. Conserved microtubule-actin interactions in cell movement and morphogenesis. Nat. Cell Biol 2003, 5, 599–609. [Google Scholar]
- Osch, G.J.V.; Veen, S.W.V.D.; Burger, E.H.; Verwoerd-Verhoef, H.L. Chondrogenic potential of in vitro multiplied rabbit perichondrium cells cultured in alginate beads in defined medium. Tissue Eng 2000, 6, 321–330. [Google Scholar]
- Buyukhatipoglu, K.; Chang, R.; Sun, W.; Clyne, A.M. Bioprinted nanoparticles for tissue engineering applications. Tissue Eng. Part C Methods 2010, 16, 631–642. [Google Scholar]
- Wu, X.; Tan, Y.; Mao, H.; Zhang, M. Toxic effects of iron oxide nanoparticles on human umbilical vein endothelial cells. Int. J. Nanomedicine 2010, 5, 385–399. [Google Scholar]
- Soenen, S.J.H.; Himmelreich, U.; Nuytten, N.; De Cuyper, M. Cytotoxic effects of iron oxide nanoparticles and implications for safety in cell labelling. Biomaterials 2011, 32, 195–205. [Google Scholar]
- Soenen, S.J.; De Meyer, S.F.; Dresselaers, T.; Velde, G.V.; Pareyn, I.M.; Braeckmans, K.; De Cuyper, M.; Himmelreich, U.; Vanhoorelbeke, K.I. MRI assessment of blood outgrowth endothelial cell homing using cationic magnetoliposomes. Biomaterials 2011, 32, 4140–4150. [Google Scholar]
- Hong, S.C.; Lee, J.H.; Lee, J.; Kim, H.Y.; Park, J.Y.; Cho, J.; Lee, J.; Han, D.W. Subtle cytotoxicity and genotoxicity differences in superparamagnetic iron oxide nanoparticles coated with various functional groups. Int. J. Nanomedicine 2011, 6, 3219–3231. [Google Scholar]
- Carmen Bautista, M.; Bomati-Miguel, O.; del Puerto Morales, M.; Serna, C.J.; Veintemillas-Verdaguer, S. Surface characterisation of dextran-coated iron oxide nanoparticles prepared by laser pyrolysis and coprecipitation. J. Magn. Magn. Mater 2005, 293, 20–27. [Google Scholar]
- Zhang, J.; Rana, S.; Srivastava, R.S.; Misra, R.D.K. On the chemical synthesis and drug delivery response of folate receptor-activated, polyethylene glycol-functionalized magnetite nanoparticles. Acta Biomater 2008, 4, 40–48. [Google Scholar]
- White, M.J.; DiCaprio, M.J.; Greenberg, D.A. Assessment of neuronal viability with Alamar blue in cortical and granule cell cultures. J. Neurosci. Meth. 1996, 70, 195–200. [Google Scholar]






© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yu, M.; Huang, S.; Yu, K.J.; Clyne, A.M. Dextran and Polymer Polyethylene Glycol (PEG) Coating Reduce Both 5 and 30 nm Iron Oxide Nanoparticle Cytotoxicity in 2D and 3D Cell Culture. Int. J. Mol. Sci. 2012, 13, 5554-5570. https://doi.org/10.3390/ijms13055554
Yu M, Huang S, Yu KJ, Clyne AM. Dextran and Polymer Polyethylene Glycol (PEG) Coating Reduce Both 5 and 30 nm Iron Oxide Nanoparticle Cytotoxicity in 2D and 3D Cell Culture. International Journal of Molecular Sciences. 2012; 13(5):5554-5570. https://doi.org/10.3390/ijms13055554
Chicago/Turabian StyleYu, Miao, Shaohui Huang, Kevin Jun Yu, and Alisa Morss Clyne. 2012. "Dextran and Polymer Polyethylene Glycol (PEG) Coating Reduce Both 5 and 30 nm Iron Oxide Nanoparticle Cytotoxicity in 2D and 3D Cell Culture" International Journal of Molecular Sciences 13, no. 5: 5554-5570. https://doi.org/10.3390/ijms13055554
APA StyleYu, M., Huang, S., Yu, K. J., & Clyne, A. M. (2012). Dextran and Polymer Polyethylene Glycol (PEG) Coating Reduce Both 5 and 30 nm Iron Oxide Nanoparticle Cytotoxicity in 2D and 3D Cell Culture. International Journal of Molecular Sciences, 13(5), 5554-5570. https://doi.org/10.3390/ijms13055554