Galectin-1 Is an Interactive Protein of Selenoprotein M in the Brain


Abstract
:1. Introduction
2. Results
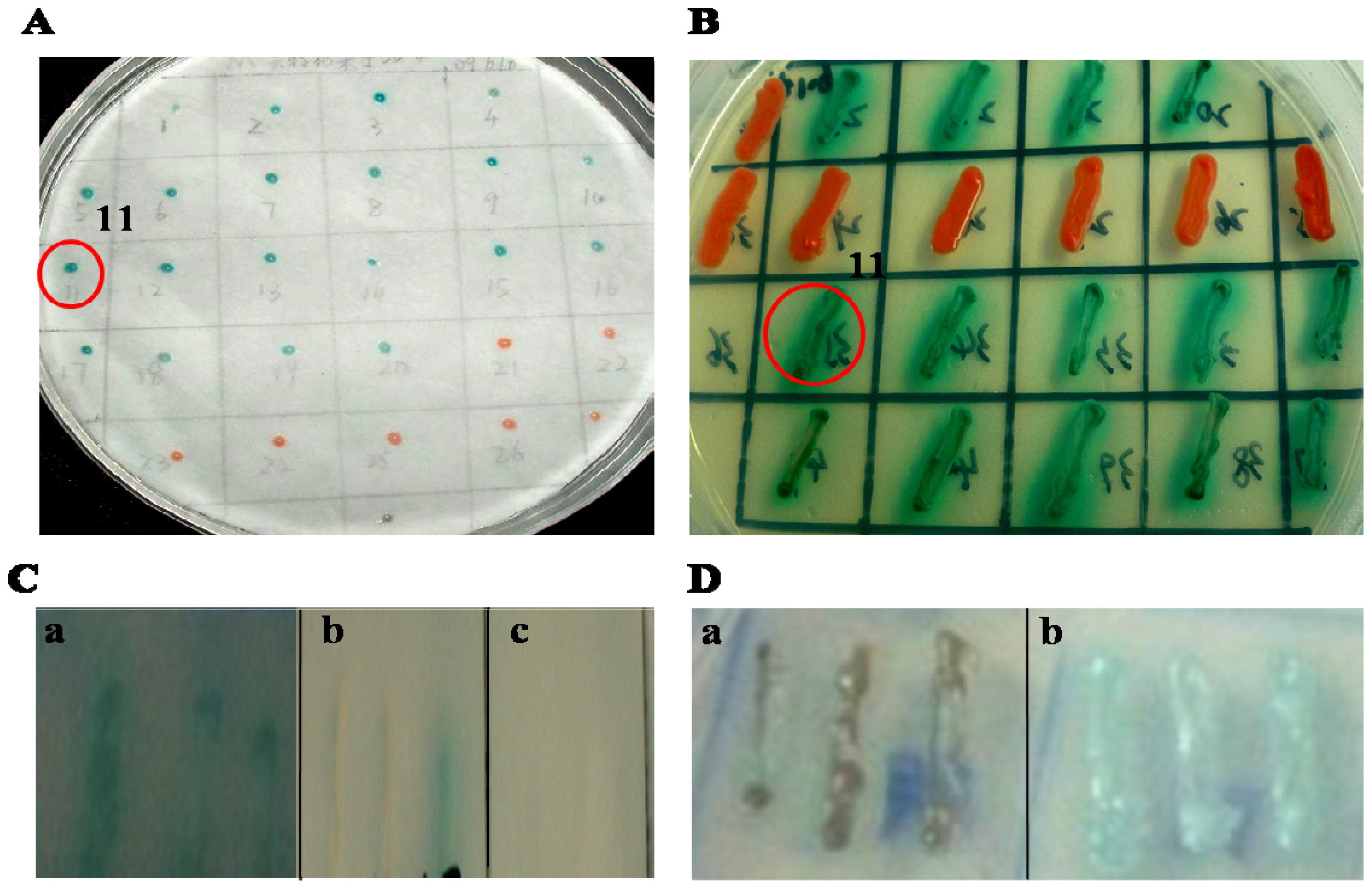
2.1. Identification of Gal-1 as an Interactive Protein of SelM′
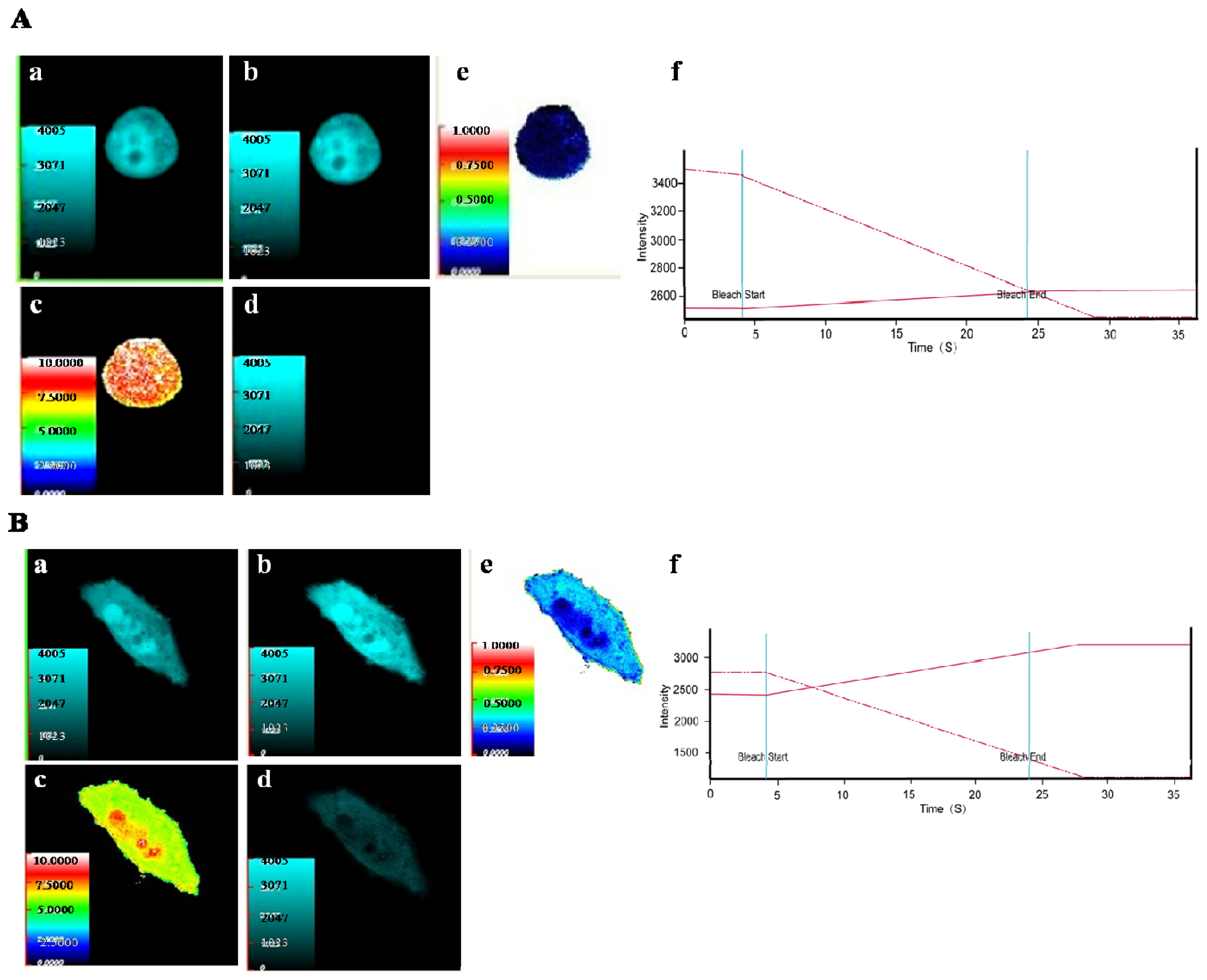
2.2. Verification of the Protein Interaction by FRET Method
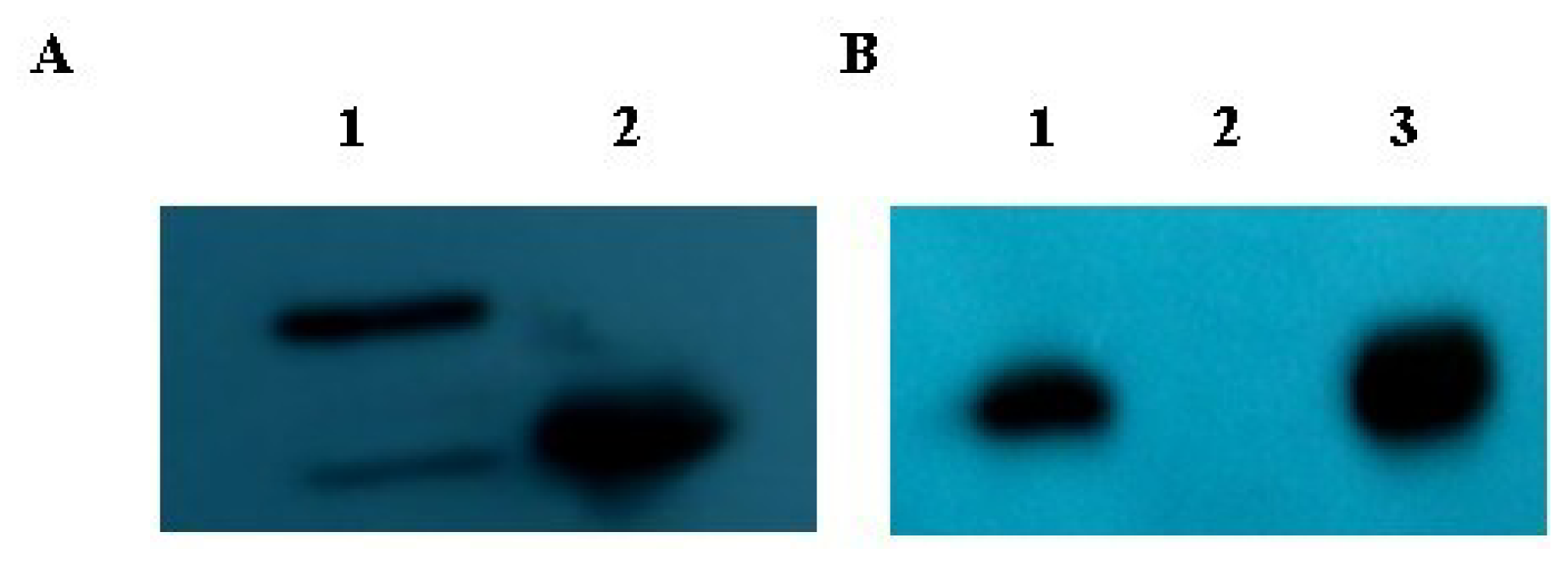
2.3. Verification of the Direct Protein Interaction by GST Pull-Down Assay
2.4. Verification of the Protein Interaction by Co-Immunoprecipitation Assay
3. Discussion
4. Experimental Section
4.1. Materials and Reagents
4.2. Gene Amplification and Plasmid Construction
4.3. Library Screening via the Yeast Two-Hybrid System
4.4. Mammalian Cell Culture
4.5. Fluorescence Resonance Energy Transfer (FRET) Detection
4.6. GST Pull-Down Assay and Western Blot Analysis
4.7. Co-Immunoprecipitation Assay
5. Conclusions





{kind=link}
{kind=link}
{kind=link}
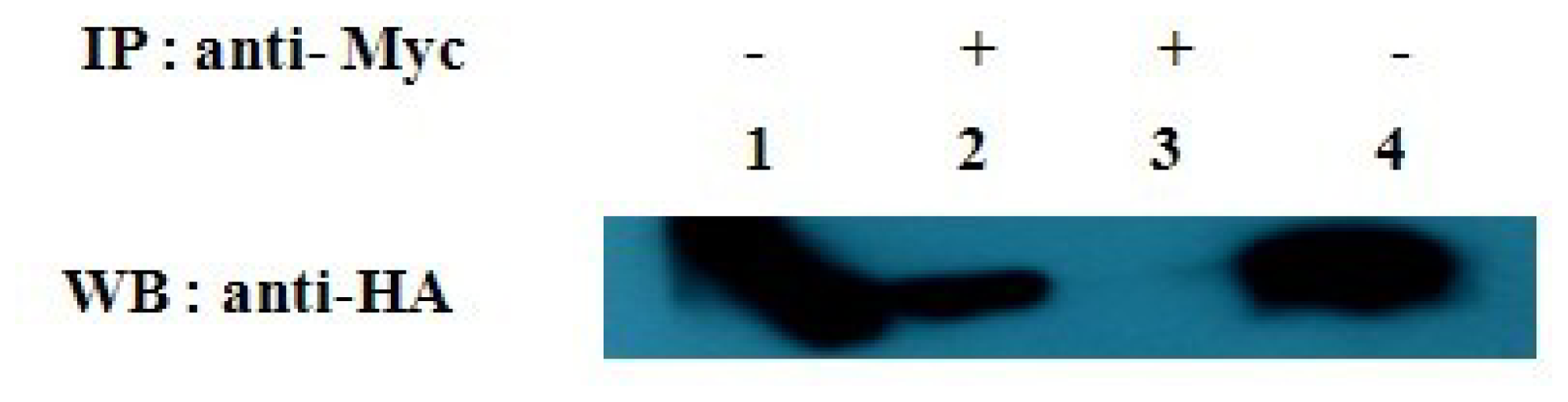{kind=link}

Acknowledgments
Conflicts of Interest
References
- Godwin, K.; Kuchel, R.; Buckley, R. The effect of selenium on infertility in ewesgrazing improved pastures. Aust. J. Exp. Agric. Anim. Husb 1970, 10, 672–678. [Google Scholar]
- Paszkowski, T.; Traub, A.; Robinson, S.; McMaster, D. Selenium dependent glutathione peroxidase activity in human follicular fluid. Clin. Chim. Acta 1995, 236, 173–180. [Google Scholar]
- Sies, H. Biochemistry of oxidative stress. Angew. Chem. Int. Ed. Engl 1986, 25, 1058–1071. [Google Scholar]
- Bellinger, F.P.; Raman, A.V.; Reeves, M.A.; Berry, M.J. Regulation and function of selenoproteins in human diseases. Biochem. J 2009, 422, 11–22. [Google Scholar]
- Koller, L.D.; Exon, J.H. The two faces of selenium deficiency and toxicity—Are similar in animals and man. Can. J. Vet. Res 1986, 50, 297–306. [Google Scholar]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [Green Version]
- Berry, M.J.; Tujebajeva, R.M.; Copeland, P.R.; Xu, X.M.; Carlson, B.A.; Martin, G.W., III; Low, S.C.; Mansell, J.B.; Grundner-Culemann, E.; Harney, J.W.; et al. Selenocysteine incorporation directed from the 3’UTR: Characterization of eukaryotic EFsec and mechanistic implications. Biofactors 2001, 14, 17–24. [Google Scholar]
- Gromer, S.; Eubel, J.K.; Jacob, J. Human selenoproteins at a glance. Cell Mol. Life Sci 2005, 62, 2414–2437. [Google Scholar]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar]
- Korotkov, K.V.; Novoselov, S.V.; Hatfield, D.L.; Gladyshev, V.N. Mammalian selenoprotein in which selenocysteine (Sec) incorporation is supported by a new form of sec insertion sequence element. Mol. Cell Biol 2002, 22, 1402–1411. [Google Scholar]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. The Sep15 protein family: Roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB Life 2007, 59, 1–5. [Google Scholar]
- Hwang, D.Y.; Sin, J.S.; Kim, M.S.; Yim, S.Y.; Kim, Y.K.; Kim, C.K.; Kim, B.G.; Shim, S.B.; Jee, S.W.; Lee, S.H.; et al. Overexpression of human selenoprotein M differentially regulates the concentrations of antioxidants and H2O2, the activity of antioxidant enzymes, and the composition of white blood cells in a transgenic rat. Int. J. Mol. Med 2008, 21, 169–179. [Google Scholar]
- Yim, S.Y.; Chae, K.R.; Shim, S.B.; Hong, J.T.; Park, J.Y.; Lee, C.Y.; Son, H.J.; Sheen, Y.Y.; Hwang, D.Y. ERK activation induced by selenium treatment significantly downregulates beta/gamma-secretase activity and Tau phosphorylation in the transgenic rat overexpressing human selenoprotein M. Int. J. Mol. Med 2009, 24, 91–96. [Google Scholar]
- Reeves, M.A.; Bellinger, F.P.; Berry, M.J. The neuroprotective functions of selenoprotein M and its role in cytosolic calcium regulation. Antioxid. Redox. Signal 2010, 12, 809–818. [Google Scholar]
- Ferguson, A.D.; Labunskyy, V.M.; Fomenko, D.E.; Araç, D.; Chelliah, Y.; Amezcua, C.A.; Rizo, J.; Gladyshev, V.N.; Deisenhofer, J. NMR structures of the selenoproteins Sep15 and SelM reveal redox activity of a new thioredoxin-like family. J. Biol. Chem 2006, 281, 3536–3543. [Google Scholar]
- Kato, T.; Ren, C.H.; Wada, M.; Kawanami, T. Galectin-1 as a potential therapeutic agent for amyotrophic lateral sclerosis. Curr. Drug Targets 2005, 6, 407–418. [Google Scholar]
- Ishibashi, S.; Kuroiwa, T.; Sakaguchi, M.; Sun, L.; Kadoya, T.; Okano, H.; Mizusawa, H. Galectin-1 regulates neurogenesis in the subventricular zone and promotes functional recovery after stroke. Exp. Neurol 2007, 207, 302–313. [Google Scholar]
- Gaudet, A.D.; Steeves, J.D.; Tetzlaff, W.; Ramer, M.S. Expression and functions of galectin-1 in sensory and motoneurons. Curr. Drug Targets 2005, 6, 419–425. [Google Scholar]
- Camby, I.; Mercier, M.L.; Lefranc, F.; Kiss, R. Galectin-1: A small protein with major functions. Glycobiology 2006, 16, 137R–157R. [Google Scholar]
- Clerch, L.B.; Whitney, P.; Hass, M.; Brew, K.; Miller, T.; Werner, R.; Massaro, D. Sequence of a full-length cDNA for rat lung beta-galactoside-binding protein: Primary and secondary structure of the lectin. Biochemistry 1988, 27, 692–629. [Google Scholar]
- Sakaguchi, M.; Shingo, T.; Shimazaki, T.; Okano, H.J.; Shiwa, M.; Ishibashi, S.; Oguro, H.; Ninomiya, M.; Kadoya, T.; Horie, H.; et al. A carbohydrate-binding protein, galectin-1, promotes proliferation of adult neural stem cells. Proc. Natl Acad. Sci. USA 2006, 103, 7112–7117. [Google Scholar]
- Imaizumi, Y.; Sakaguchi, M.; Morishita, T.; Ito, M.; Poirier, F.; Sawamoto, K.; Okano, H. Galectin-1 is expressed in early-type neural progenitor cells and down-regulates neurogenesis in the adult hippocampus. Mol. Brain 2011, 4, 7. [Google Scholar]
- Kurushima, H.; Ohno, M.; Miura, T.; Nakamura, T.Y.; Horie, H.; Kadoya, T.; Ooboshi, H.; Kitazono, T.; Ibayashi, S.; Iida, M.; et al. Selective induction of DeltaFosB in the brain after transient forebrain ischemia accompanied by an increased expression of galectin-1, and the implication of DeltaFosB and galectin-1 in neuroprotection and neurogenesis. Cell Death Differ 2005, 12, 1078–1096. [Google Scholar]
- Tahara, K.; Tsuchimoto, D.; Tominaga, Y.; Asoh, S.; Ohta, S.; Kitagawa, M.; Horie, H.; Kadoya, T.; Nakabeppu, Y. DetaFosB, but not FosB, induces delayed apoptosis independent of cell proliferation in the Rat1a embryo cell line. Cell Death Differ 2003, 10, 496–507. [Google Scholar]
- Miura, T.; Takahashi, M.; Horie, H.; Kurushima, H.; Tsuchimoto, D.; Sakumi, K.; Nakabeppu, Y. Galectin-1beta, a natural monomeric form of galectin-1 lacking its six amino-terminal residues promotes axonal regeneration but not cell death. Cell Death Differ 2004, 11, 1076–1083. [Google Scholar]
- Miura, T.; Ohnishi, Y.; Kurushima, H.; Horie, H.; Kadoya, T.; Nakabeppu, Y. Regulation of the neuronal fate by DeltaFosB and its downstream target, galectin-1. Curr. Drug Targets 2005, 6, 437–444. [Google Scholar]
- Kajitani, K.; Nomaru, H.; Ifuku, M.; Yutsudo, N.; Dan, Y.; Miura, T.; Tsuchimoto, D.; Sakumi, K.; Kadoya, T.; Horie, H.; et al. Galectin-1 promotes basal and kainate-induced proliferation of neural progenitors in the dentate gyrus of adult mouse hippocampus. Cell Death Differ 2009, 16, 417–427. [Google Scholar]
- Sakaguchi, M.; Okano, H. Neural stem cells, adult neurogenesis, and galectin-1: From bench to bedside. Dev. Neurobiol 2012, 72, 1059–1067. [Google Scholar]
- Song, H.; Stevens, C.F.; Gage, F.H. Astroglia induce neurogenesis from adult neural stem cells. Nature 2002, 417, 39–44. [Google Scholar]
- Sasaki, T.; Hirabayashi, J.; Manya, H.; Kasai, K.; Endo, T. Galectin-1 induces astrocyte differentiation, which leads to production of brain-derived neurotrophic factor. Glycobiology 2004, 14, 357–363. [Google Scholar]
- Zhao, C.; Deng, W.; Gage, F.H. Mechanisms and functional implications of adult neurogenesis. Cell 2008, 132, 645–660. [Google Scholar]
- Lee, J.; Duan, W.; Mattson, M.P. Evidence that brain-derived neurotrophic factor is required for basal neurogenesis and mediates, in part, the enhancement of neurogenesis by dietary restriction in the hippocampus of adult mice. J. Neurochem 2002, 82, 1367–1375. [Google Scholar]
- Nishioka, T.; Sakumi, K.; Miura, T.; Tahara, K.; Horie, H.; Kadoya, T.; Nakabeppu, Y. FosB gene products trigger cell proliferation and morphological alteration with an increased expression of a novel processed form of galectin-1 in the rat 3Y1 embryo cell line. J. Biochem 2002, 131, 653–661. [Google Scholar]
- Kobayashi, K.; Imagama, S.; Ohgomori, T.; Hirano, K.; Uchimura, K.; Sakamoto, K.; Hirakawa, A.; Takeuchi, H.; Suzumura, A.; Ishiguro, N.; et al. Minocycline selectively inhibits M1 polarization of microglia. Cell Death Dis 2013, 4, e525. [Google Scholar]
- Motohiro, N.; Minoru, F. Galectin-1 for neuroprotection? Immunity 2012, 37, 187–189. [Google Scholar]
- Starossom, S.C.; Mascanfroni, I.D.; Imitola, J.; Cao, L.; Raddassi, K.; Hernandez, S.F.; Bassil, R.; Croci, D.O.; Cerliani, J.P.; Delacour, D.; et al. Galectin-1 deactivates classically activated microglia and protects from inflammation-induced neurodegeneration. Immunity 2012, 37, 249–263. [Google Scholar]
- Egnaczyk, G.F.; Pomonis, J.D.; Schmidt, J.A.; Rogers, S.D.; Peters, C.; Ghilardi, J.R.; Mantyh, P.W.; Maggio, J.E. Proteomic analysis of the reactive phenotype of astrocytes following endothelin-1 exposure. Proteomics 2003, 3, 689–398. [Google Scholar]
- Chen, P.; Wang, C.; Ma, X.; Zhang, Y.; Liu, Q.; Qiu, S.; Liu, Q.; Tian, J.; Ni, J. Direct interaction of selenoprotein R with clusterin and its possible role in Alzheimer’s disease. PLoS One 2013, 8, e66384. [Google Scholar]
- Liang, X.Y.; Liu, Q.; Chen, P.; Huo, K.K.; Hu, T.Y.; Ni, J.Z. Screen and verification of interactive protein of selenoprotein k in human liver. Chem. J. Chin. Univ 2012, 23, 1–5. [Google Scholar]
- Du, X.B.; Li, H.P.; Wang, Z.; Qiu, S.; Liu, Q.; Ni, J. Selenoprotein P and selenoprotein M block Zn2+-mediated Aβ42 aggregation and toxicity. Metallomics 2013, 5, 861–870. [Google Scholar]
- Chen, P.; Wang, R.D.; Ma, X.J.; Liu, Q.; Ni, J.Z. Different forms of selenoprotein M differentially affect Aβ aggregation and ROS generation. Int. J. Mol. Sci 2013, 14, 4385–4399. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Qiao, X.; Tian, J.; Chen, P.; Wang, C.; Ni, J.; Liu, Q. Galectin-1 Is an Interactive Protein of Selenoprotein M in the Brain. Int. J. Mol. Sci. 2013, 14, 22233-22245. https://doi.org/10.3390/ijms141122233
Qiao X, Tian J, Chen P, Wang C, Ni J, Liu Q. Galectin-1 Is an Interactive Protein of Selenoprotein M in the Brain. International Journal of Molecular Sciences. 2013; 14(11):22233-22245. https://doi.org/10.3390/ijms141122233
Chicago/Turabian StyleQiao, Xifeng, Jing Tian, Ping Chen, Chao Wang, Jiazuan Ni, and Qiong Liu. 2013. "Galectin-1 Is an Interactive Protein of Selenoprotein M in the Brain" International Journal of Molecular Sciences 14, no. 11: 22233-22245. https://doi.org/10.3390/ijms141122233
APA StyleQiao, X., Tian, J., Chen, P., Wang, C., Ni, J., & Liu, Q. (2013). Galectin-1 Is an Interactive Protein of Selenoprotein M in the Brain. International Journal of Molecular Sciences, 14(11), 22233-22245. https://doi.org/10.3390/ijms141122233