Towards Lipidomics of Low-Abundant Species for Exploring Tumor Heterogeneity Guided by High-Resolution Mass Spectrometry Imaging


Abstract
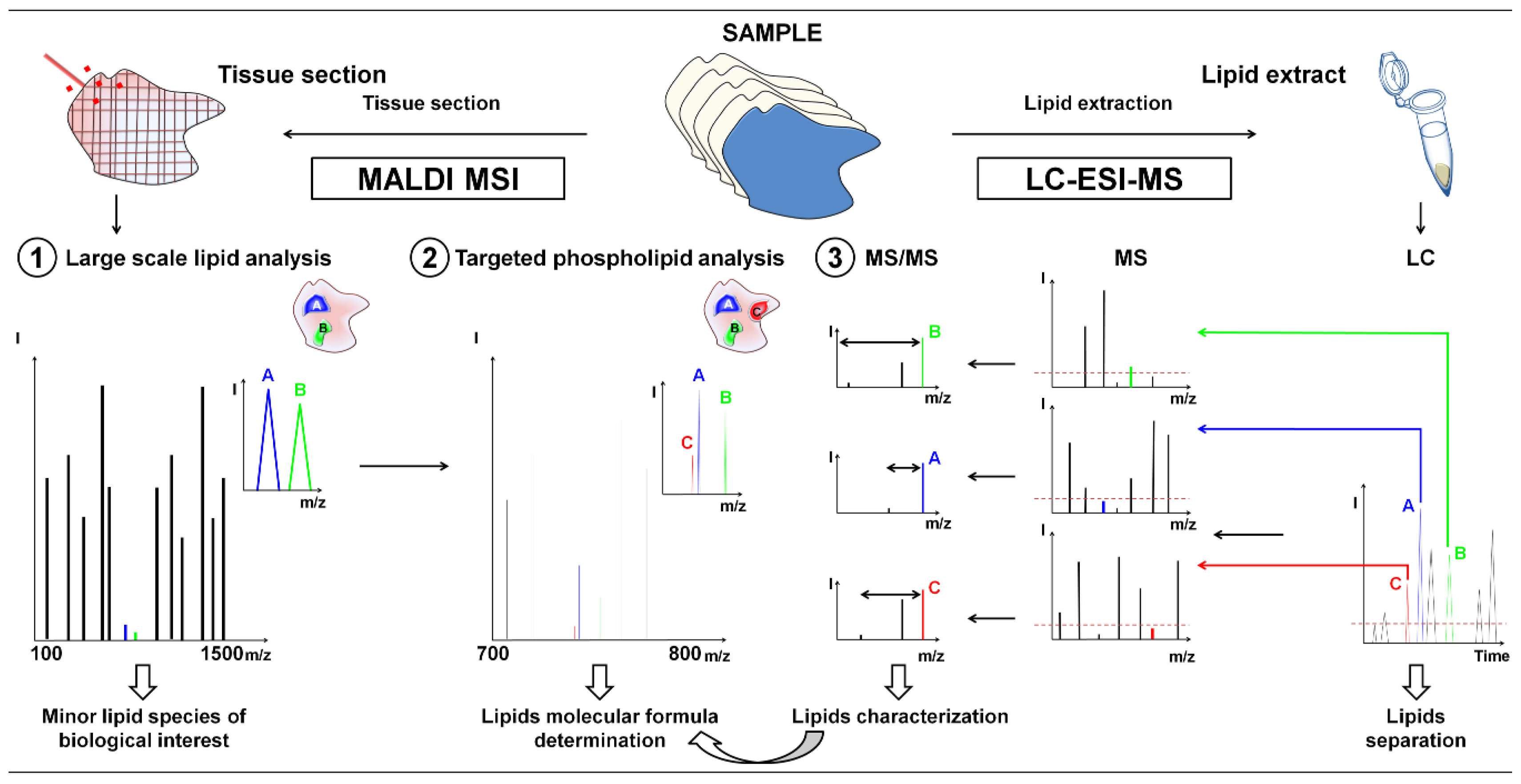
:1. Introduction
2. Results and Discussion
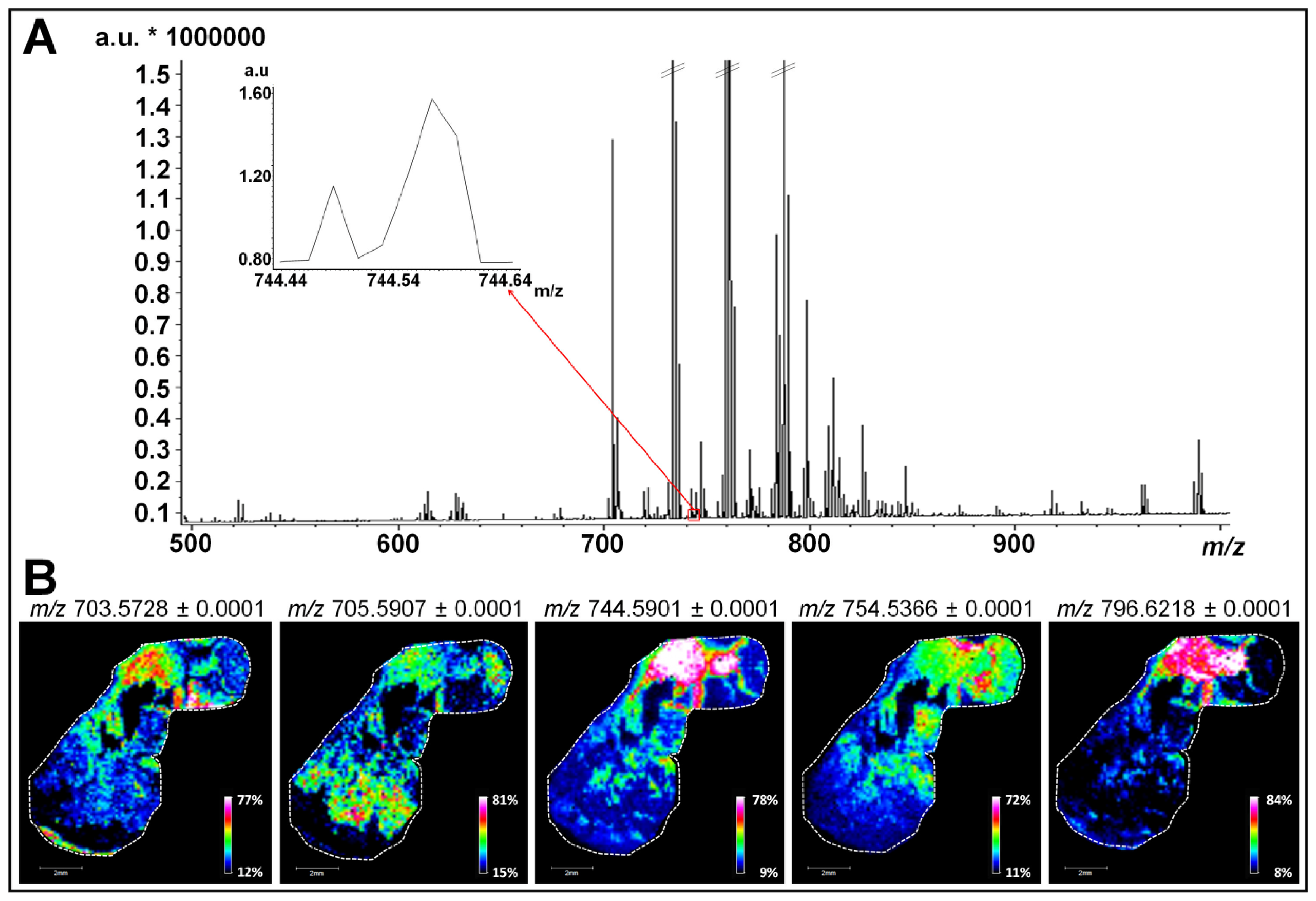
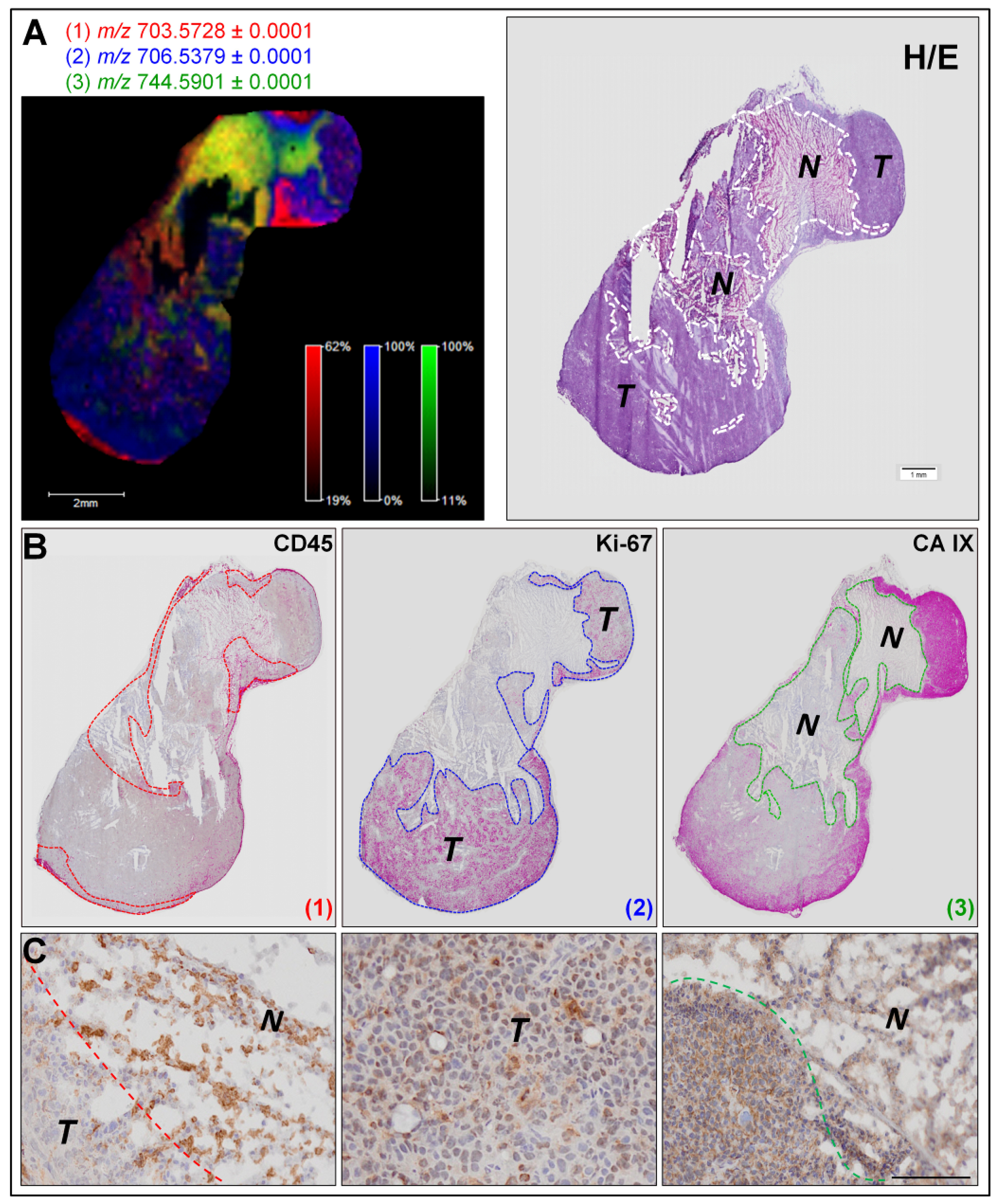
2.1. In Situ Analysis by MALDI-FTICR MSI and Immunohistochemistry Reveal Specific Low-Abundant Lipid Species and Breast Tumor Heterogeneity
2.2. Use of High-Resolution Mass Spectrometry for the Identification of Minor Lipid Species with Biological Relevance
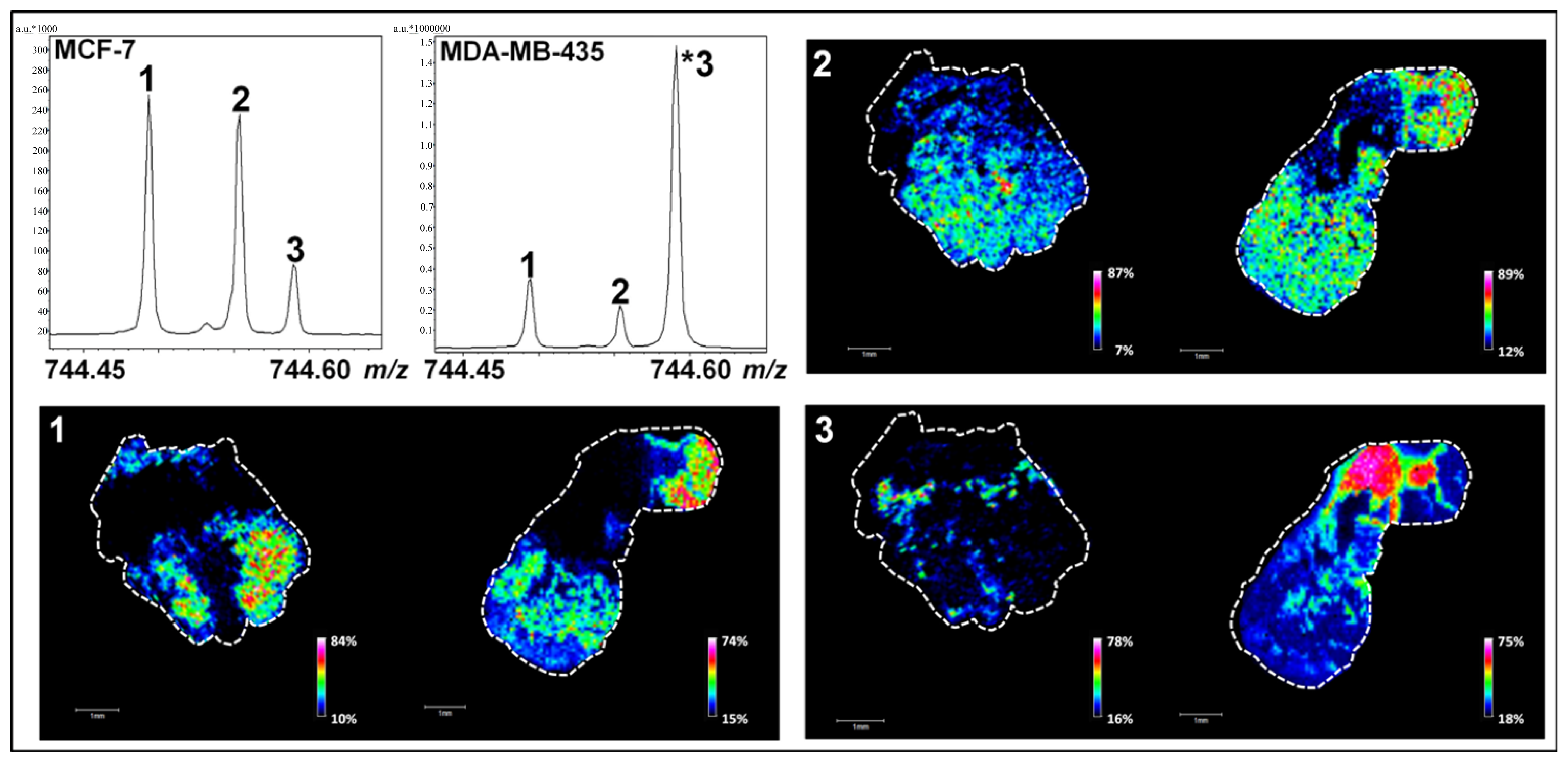
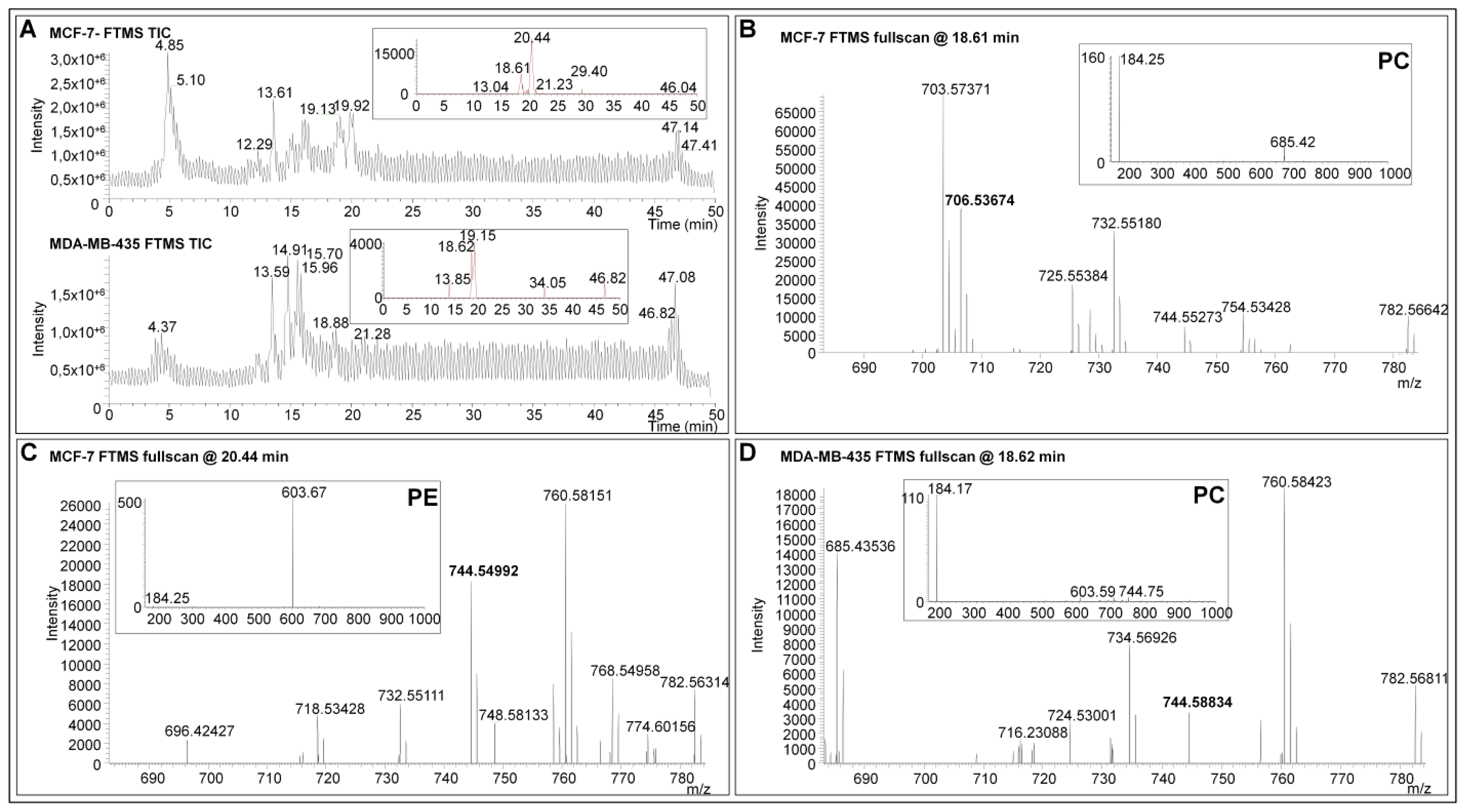
2.3. Targeted Characterization of Specific Low-Abundant Phospholipid Species in Complex Mixtures by LC-ESI-MS
3. Experimental Section
3.1. Reagents
3.2. Phospholipid Standards
3.3. Xenografts
3.4. Tissue Preparation and Immunochemistry
3.5. MALDI Fourier Transform-Ion Cyclotron Resonance MSI Analyses
3.5.1. Matrix Deposition
3.5.2. Mass Spectrometry
3.6. Probabilistic Latent Semantic Analysis
3.7. LC-ESI-MS Analyses
3.7.1. Phospholipid Standard Preparations
3.7.2. Lipid Extraction from MDA-MB-231, MDA-MB-435, and MCF-7 Tumors
3.7.3. LC-ESI-MS Analyses
4. Conclusions
Supplementary Information
ijms-14-24560-s002.pdfAcknowledgments
Conflicts of Interest
References
- Shevchenko, A.; Simons, K. Lipidomics: Coming to grips with lipid diversity. Nat. Rev 2010, 11, 593–598. [Google Scholar]
- Kasson, P.M.; Pande, V.S. Control of membrane fusion mechanism by lipid composition: Predictions from ensemble molecular dynamics. PLoS Comput. Boil 2007, 3, e220. [Google Scholar]
- Aguilar, P.S.; Heiman, M.G.; Walther, T.C.; Engel, A.; Schwudke, D.; Gushwa, N.; Kurzchalia, T.; Walter, P. Structure of sterol aliphatic chains affects yeast cell shape and cell fusion during mating. Proc. Natl. Acad. Sci. USA 2010, 107, 4170–4175. [Google Scholar]
- Fujimoto, K.; Iwasaki, C.; Kawaguchi, H.; Yasugi, E.; Oshima, M. Cell membrane dynamics and the induction of apoptosis by lipid compounds. FEBS Lett 1999, 446, 113–116. [Google Scholar]
- Johansen, J.; Ramanathan, V.; Beh, C.T. Vesicle trafficking from a lipid perspective: Lipid regulation of exocytosis in Saccharomyces cerevisiae. Cell. Logist 2012, 2, 151–160. [Google Scholar]
- Janda, M.; Planchais, S.; Djafi, N.; Martinec, J.; Burketova, L.; Valentova, O.; Zachowski, A.; Ruelland, E. Phosphoglycerolipids are master players in plant hormone signal transduction. Plant Cell Rep 2013, 32, 839–851. [Google Scholar]
- Amstalden van Hove, E.R.; Blackwell, T.R.; Klinkert, I.; Eijkel, G.B.; Heeren, R.M.; Glunde, K. Multimodal mass spectrometric imaging of small molecules reveals distinct spatio-molecular signatures in differentially metastatic breast tumor models. Cancer Res 2010, 70, 9012–9021. [Google Scholar]
- Filippov, V.; Song, M.A.; Zhang, K.; Vinters, H.V.; Tung, S.; Kirsch, W.M.; Yang, J.; Duerksen-Hughes, P.J. Increased ceramide in brains with Alzheimer’s and other neurodegenerative diseases. J. Alzheimers Dis 2012, 29, 537–547. [Google Scholar]
- Bennett, S.A.; Valenzuela, N.; Xu, H.; Franko, B.; Fai, S.; Figeys, D. Using neurolipidomics to identify phospholipid mediators of synaptic (dys)function in Alzheimer’s disease. Front. Physiol 2013. [Google Scholar] [CrossRef]
- Pellieux, C.; Montessuit, C.; Papageorgiou, I.; Pedrazzini, T.; Lerch, R. Differential effects of high-fat diet on myocardial lipid metabolism in failing and nonfailing hearts with angiotensin II-mediated cardiac remodeling in mice. Am. J. Physiol. Heart Circ. Physiol 2012, 302, 1795–1805. [Google Scholar]
- Minkler, P.E.; Hoppel, C.L. Separation and characterization of cardiolipin molecular species by reverse-phase ion pair high-performance liquid chromatography-mass spectrometry. J Lipid Res 2010, 51, 856–865. [Google Scholar]
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma lysophosphatidylcholine levels are reduced in obesity and type 2 diabetes. PLoS One 2012, 7, e41456. [Google Scholar]
- Wenk, M.R. Lipidomics: New tools and applications. Cell 2010, 143, 888–895. [Google Scholar]
- Kimura, K.; Yamaoka, M.; Kamisaka, Y. Rapid estimation of lipids in oleaginous fungi and yeasts using Nile red fluorescence. J. Microbiol. Methods 2004, 56, 331–338. [Google Scholar]
- Van Goor, H.; Gerrits, P.O.; Grond, J. The application of lipid-soluble stains in plastic-embedded sections. Histochemistry 1986, 85, 251–253. [Google Scholar]
- Govender, T.; Ramanna, L.; Rawat, I.; Bux, F. BODIPY staining, an alternative to the Nile Red fluorescence method for the evaluation of intracellular lipids in microalgae. Bioresour. Technol 2012, 114, 507–511. [Google Scholar]
- Cooper, M.S.; Hardin, W.R.; Petersen, T.W.; Cattolico, R.A. Visualizing “green oil” in live algal cells. J. Biosci. Bioeng 2010, 109, 198–201. [Google Scholar]
- Wenk, M.R. The emerging field of lipidomics. Nat. Rev. Drug. Discov 2005, 4, 594–610. [Google Scholar]
- Sparvero, L.J.; Amoscato, A.A.; Dixon, C.E.; Long, J.B.; Kochanek, P.M.; Pitt, B.R.; Bayir, H.; Kagan, V.E. Mapping of phospholipids by MALDI imaging (MALDI-MSI): Realities and expectations. Chem. Phys. Lipids 2012, 165, 545–562. [Google Scholar]
- Berry, K.A.; Hankin, J.A.; Barkley, R.M.; Spraggins, J.M.; Caprioli, R.M.; Murphy, R.C. MALDI imaging of lipid biochemistry in tissues by mass spectrometry. Chem. Rev 2011, 111, 6491–6512. [Google Scholar]
- Chen, Y.; Allegood, J.; Liu, Y.; Wang, E.; Cachon-Gonzalez, B.; Cox, T.M.; Merrill, A.H., Jr.; Sullards, M.C. Imaging MALDI mass spectrometry using an oscillating capillary nebulizer matrix coating system and its application to analysis of lipids in brain from a mouse model of Tay-Sachs/Sandhoff disease. Anal. Chem 2008, 80, 2780–2788. [Google Scholar]
- Murphy, R.C.; Hankin, J.A.; Barkley, R.M. Imaging of lipid species by MALDI mass spectrometry. J. Lipid Res 2009, 50, 317–322. [Google Scholar]
- Bure, C.; Ayciriex, S.; Testet, E.; Schmitter, J.M. A single run LC-MS/MS method for phospholipidomics. Anal. Bioanal. Chem 2013, 405, 203–213. [Google Scholar]
- Vichi, S.; Cortes-Francisco, N.; Caixach, J. Ultrahigh resolution mass spectrometry and accurate mass measurements for high-throughput food lipids profiling. J. Mass Spectrom 2012, 47, 1177–1190. [Google Scholar]
- Jove, M.; Ayala, V.; Ramirez-Nunez, O.; Serrano, J.C.; Cassanye, A.; Arola, L.; Caimari, A.; del Bas, J.M.; Crescenti, A.; Pamplona, R.; et al. Lipidomic and metabolomic analyses reveal potential plasma biomarkers of early atheromatous plaque formation in hamsters. Cardiovasc. Res 2013, 97, 642–652. [Google Scholar]
- Schuhmann, K.; Almeida, R.; Baumert, M.; Herzog, R.; Bornstein, S.R.; Shevchenko, A. Shotgun lipidomics on a LTQ Orbitrap mass spectrometer by successive switching between acquisition polarity modes. J. Mass Spectrom 2012, 47, 96–104. [Google Scholar] [Green Version]
- Schuhmann, K.; Herzog, R.; Schwudke, D.; Metelmann-Strupat, W.; Bornstein, S.R.; Shevchenko, A. Bottom-up shotgun lipidomics by higher energy collisional dissociation on LTQ Orbitrap mass spectrometers. Anal. Chem 2011, 83, 5480–5487. [Google Scholar]
- Chaurand, P.; Cornett, D.S.; Angel, P.M.; Caprioli, R.M. From whole-body sections down to cellular level, multiscale imaging of phospholipids by MALDI mass spectrometry. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef]
- Thomas, A.; Charbonneau, J.L.; Fournaise, E.; Chaurand, P. Sublimation of new matrix candidates for high spatial resolution imaging mass spectrometry of lipids: Enhanced information in both positive and negative polarities after 1,5-diaminonapthalene deposition. Anal. Chem 2012, 84, 2048–2054. [Google Scholar]
- Horn, P.J.; Korte, A.R.; Neogi, P.B.; Love, E.; Fuchs, J.; Strupat, K.; Borisjuk, L.; Shulaev, V.; Lee, Y.J.; Chapman, K.D. Spatial mapping of lipids at cellular resolution in embryos of cotton. Plant cell 2012, 24, 622–636. [Google Scholar]
- Vance, J.E. Phosphatidylserine and phosphatidylethanolamine in mammalian cells: Two metabolically related aminophospholipids. J. Lipid Res 2008, 49, 1377–1387. [Google Scholar]
- Tyurin, V.A.; Tyurina, Y.Y.; Ritov, V.B.; Lysytsya, A.; Amoscato, A.A.; Kochanek, P.M.; Hamilton, R.; Dekosky, S.T.; Greenberger, J.S.; Bayir, H.; et al. Oxidative lipidomics of apoptosis: Quantitative assessment of phospholipid hydroperoxides in cells and tissues. Methods Mol. Boil 2010, 610, 353–374. [Google Scholar]
- Mock, J.N.; Taliaferro, J.P.; Lu, X.; Patel, S.K.; Cummings, B.S.; Long, T.E. Haloenol pyranones and morpholinones as antineoplastic agents of prostate cancer. Bioorg. Med. Chem. Lett 2012, 22, 4854–4858. [Google Scholar]
- Girotti, A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J. Lipid Res 1998, 39, 1529–1542. [Google Scholar]
- Guijas, C.; Astudillo, A.M.; Gil-de-Gomez, L.; Rubio, J.M.; Balboa, M.A.; Balsinde, J. Phospholipid sources for adrenic acid mobilization in RAW 264.7 macrophages. Comparison with arachidonic acid. Biochim. Biophys. Acta 2012, 1821, 1386–1393. [Google Scholar] [Green Version]
- Feige, E.; Mendel, I.; George, J.; Yacov, N.; Harats, D. Modified phospholipids as anti-inflammatory compounds. Curr. Opin. Lipidol 2010, 21, 525–529. [Google Scholar]
- Damnjanovic, Z.; Jovanovic, M.; Nagorni, A.; Radojkovic, M.; Sokolovic, D.; Damnjanovic, G.; Djindjic, B.; Smiljkovic, I.; Kamenov, A.; Damnjanovic, I. Correlation of inflammation parameters and biochemical markers of cholestasis with the intensity of lipid peroxidation in patients with choledocholithiasis. Vojnosanit. Pregl 2013, 70, 170–176. [Google Scholar]
- Simoes, C.; Silva, A.C.; Domingues, P.; Laranjeira, P.; Paiva, A.; Domingues, M.R. Modified phosphatidylethanolamines induce different levels of cytokine expression in monocytes and dendritic cells. Chem. Phys. Lipids 2013, 175–176, 57–64. [Google Scholar]
- Leibundgut, G.; Scipione, C.; Yin, H.; Schneider, M.; Boffa, M.B.; Green, S.; Yang, X.; Dennis, E.A.; Witztum, J.L.; Koschinsky, M.L.; Tsimikas, S. Determinants of binding of oxidized phospholipids on apolipoprotein (a) and lipoprotein (a). J. Lipid Res 2013, 54, 2815–2830. [Google Scholar]
- Shan, X.; Wang, D.; Chen, J.; Xiao, X.; Jiang, Y.; Wang, Y.; Fan, Y. Necrosis degree displayed in computed tomography images correlated with hypoxia and angiogenesis in breast cancer. J. Comput. Assist. Tomogr 2013, 37, 22–28. [Google Scholar]
- Kim, K.; Park, W.Y.; Kim, J.Y.; Sol, M.Y.; Shin, D.H.; Park, D.Y.; Lee, C.H.; Lee, J.H.; Choi, K.U. Prognostic relevance of the expression of CA IX, GLUT-1, and VEGF in ovarian epithelial cancers. Korean J. Pathol 2012, 46, 532–540. [Google Scholar]
- Doria, M.L.; Cotrim, C.Z.; Simoes, C.; Macedo, B.; Domingues, P.; Domingues, M.R.; Helguero, L.A. Lipidomic analysis of phospholipids from human mammary epithelial and breast cancer cell lines. J. cell. Physiol 2012, 228, 457–468. [Google Scholar]
- Monteggia, E.; Colombo, I.; Guerra, A.; Berra, B. Phospholipid distribution in murine mammary adenocarcinomas induced by activated neu oncogene. Cancer Detect. Prev 2000, 24, 207–211. [Google Scholar]
- Dueck, D.A.; Chan, M.; Tran, K.; Wong, J.T.; Jay, F.T.; Littman, C.; Stimpson, R.; Choy, P.C. The modulation of choline phosphoglyceride metabolism in human colon cancer. Mol. Cell. Biochem 1996, 162, 97–103. [Google Scholar]
- Ruiz-Cabello, J.; Cohen, J.S. Phospholipid metabolites as indicators of cancer cell function. NMR Biomed 1992, 5, 226–233. [Google Scholar]
- Zhang, X.H.; Zhao, C.; Seleznev, K.; Song, K.; Manfredi, J.J.; Ma, Z.A. Disruption of G1-phase phospholipid turnover by inhibition of Ca2+-independent phospholipase A2 induces a p53-dependent cell-cycle arrest in G1 phase. J. Cell Sci 2006, 119, 1005–1015. [Google Scholar]
- Schifferer, R.; Liebisch, G.; Bandulik, S.; Langmann, T.; Dada, A.; Schmitz, G. ApoA-I induces a preferential efflux of monounsaturated phosphatidylcholine and medium chain sphingomyelin species from a cellular pool distinct from HDL(3) mediated phospholipid efflux. Biochim. Biophys. Acta 2007, 1771, 853–863. [Google Scholar]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab 2006, 3, 321–331. [Google Scholar]
- Maciel, E.; Domingues, P.; Domingues, M.R. Liquid chromatography/tandem mass spectrometry analysis of long-chain oxidation products of cardiolipin induced by the hydroxyl radical. Rapid Commun. Mass Spectrom 2013, 25, 316–326. [Google Scholar]
- Stubiger, G.; Aldover-Macasaet, E.; Bicker, W.; Sobal, G.; Willfort-Ehringer, A.; Pock, K.; Bochkov, V.; Widhalm, K.; Belgacem, O. Targeted profiling of atherogenic phospholipids in human plasma and lipoproteins of hyperlipidemic patients using MALDI-QIT-TOF-MS/MS. Atherosclerosis 2012, 224, 177–186. [Google Scholar]
- Uhl, O.; Glaser, C.; Demmelmair, H.; Koletzko, B. Reversed phase LC/MS/MS method for targeted quantification of glycerophospholipid molecular species in plasma. J. Chromatogr. B 2011, 879, 3556–3564. [Google Scholar]
- Murphy, R.C.; Fiedler, J.; Hevko, J. Analysis of nonvolatile lipids by mass spectrometry. Chem. Rev 2001, 101, 479–526. [Google Scholar]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar]
- Knochenmuss, R.; Dubois, F.; Dale, M.J.; Zenobi, R. The matrix suppression effect and ionization mechanisms in matrix-assisted laser desorption/ionization. Rapid Commun. Mass Spectrom 1996, 10, 871–877. [Google Scholar]
- Chan, T.-W.D.; Colburn, A.W.; Derrick, P.J. Matrix-assisted UV laser desorption. Suppression of the matrix peaks. J. Mass Spectrom 1991, 26, 342–344. [Google Scholar]
- Marusyk, A.; Almendro, V.; Polyak, K. Intra-tumour heterogeneity: A looking glass for cancer? Nat. Rev. Cancer 2012, 12, 323–334. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar]
- Sud, M.; Fahy, E.; Cotter, D.; Brown, A.; Dennis, E.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; et al. LMSD: LIPID MAPS structure database. Nucleic Acids Res 2007, 35, 527–532. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Boil. Chem 1957, 226, 497–509. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/zobs | Necrosis | Inflammation | Tumor |
|---|---|---|---|
| 703.5728 | − | + | − |
| 705.5821 | + | − | − |
| 706.5379 | − | − | + |
| 718.5748 | + | − | − |
| 720.5812 | + | − | − |
| 725.5574 | + | − | − |
| 728.5197 | − | + | − |
| 739.4666 | − | − | + |
| 740.4714 | − | − | + |
| 740.4993 | − | − | + |
| 740.5207 | + | − | − |
| 740.5573 | + | − | − |
| 741.5311 | − | − | + |
| 744.5512 | − | − | + |
| 744.5901 | + | − | − |
| 765.4836 | − | − | + |
| 768.5901 | + | − | − |
| 770.5107 | − | − | + |
| 772.5248 | − | − | + |
| 772.6217 | + | − | − |
| 774.6378 | + | − | − |
| 790.5098 | − | − | + |
| 790.5742 | + | − | − |
| 796.5252 | − | − | + |
| 796.5854 | − | + | − |
| 796.6218 | + | − | − |
| 798.5416 | − | − | + |
| m/zobs | Necrosis | Inflammation | Tumor |
|---|---|---|---|
| 704.5221 | − | + | + |
| 705.5904 | + | − | + |
| 720.5542 | − | + | + |
| 723.4943 | + | − | + |
| 730.5384 | − | + | + |
| 731.6066 | + | + | + |
| 734.5693 | − | + | + |
| 746.5698 | + | + | + |
| 746.6065 | + | − | + |
| 754.5364 | + | + | + |
| 772.5847 | + | + | + |
| 784.5725 | + | + | − |
| 784.5853 | − | + | + |
| 790.5419 | + | + | − |
| 794.6049 | + | + | − |
| m/zobs | Elemental formula 1 | m/zth | Error (ppm) |
|---|---|---|---|
| 744.49419 | C38H76NO8P ([M + K]+) | 744.49401 | 0.24 |
| 744.55385 | C41H79NO8P ([M + H]+) | 744.55378 | 0.09 |
| 744.59019 | C42H83NO7P ([M + H]+) | 744.59017 | 0.04 |
| m/zobs | Molecular formula 1 | m/zth | Error (ppm) |
|---|---|---|---|
| 796.52524 | C42H80NO8P ([M + K]+) | 796.52531 | 0.1 |
| 796.58547 | C45H83NO8P ([M + H]+) | 796.58508 | 0.5 |
| 796.62181 | C46H87NO7P ([M + H]+) | 796.62147 | 0.4 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cimino, J.; Calligaris, D.; Far, J.; Debois, D.; Blacher, S.; Sounni, N.E.; Noel, A.; De Pauw, E. Towards Lipidomics of Low-Abundant Species for Exploring Tumor Heterogeneity Guided by High-Resolution Mass Spectrometry Imaging. Int. J. Mol. Sci. 2013, 14, 24560-24580. https://doi.org/10.3390/ijms141224560
Cimino J, Calligaris D, Far J, Debois D, Blacher S, Sounni NE, Noel A, De Pauw E. Towards Lipidomics of Low-Abundant Species for Exploring Tumor Heterogeneity Guided by High-Resolution Mass Spectrometry Imaging. International Journal of Molecular Sciences. 2013; 14(12):24560-24580. https://doi.org/10.3390/ijms141224560
Chicago/Turabian StyleCimino, Jonathan, David Calligaris, Johann Far, Delphine Debois, Silvia Blacher, Nor Eddine Sounni, Agnès Noel, and Edwin De Pauw. 2013. "Towards Lipidomics of Low-Abundant Species for Exploring Tumor Heterogeneity Guided by High-Resolution Mass Spectrometry Imaging" International Journal of Molecular Sciences 14, no. 12: 24560-24580. https://doi.org/10.3390/ijms141224560
APA StyleCimino, J., Calligaris, D., Far, J., Debois, D., Blacher, S., Sounni, N. E., Noel, A., & De Pauw, E. (2013). Towards Lipidomics of Low-Abundant Species for Exploring Tumor Heterogeneity Guided by High-Resolution Mass Spectrometry Imaging. International Journal of Molecular Sciences, 14(12), 24560-24580. https://doi.org/10.3390/ijms141224560