Roles of Melatonin in Fetal Programming in Compromised Pregnancies



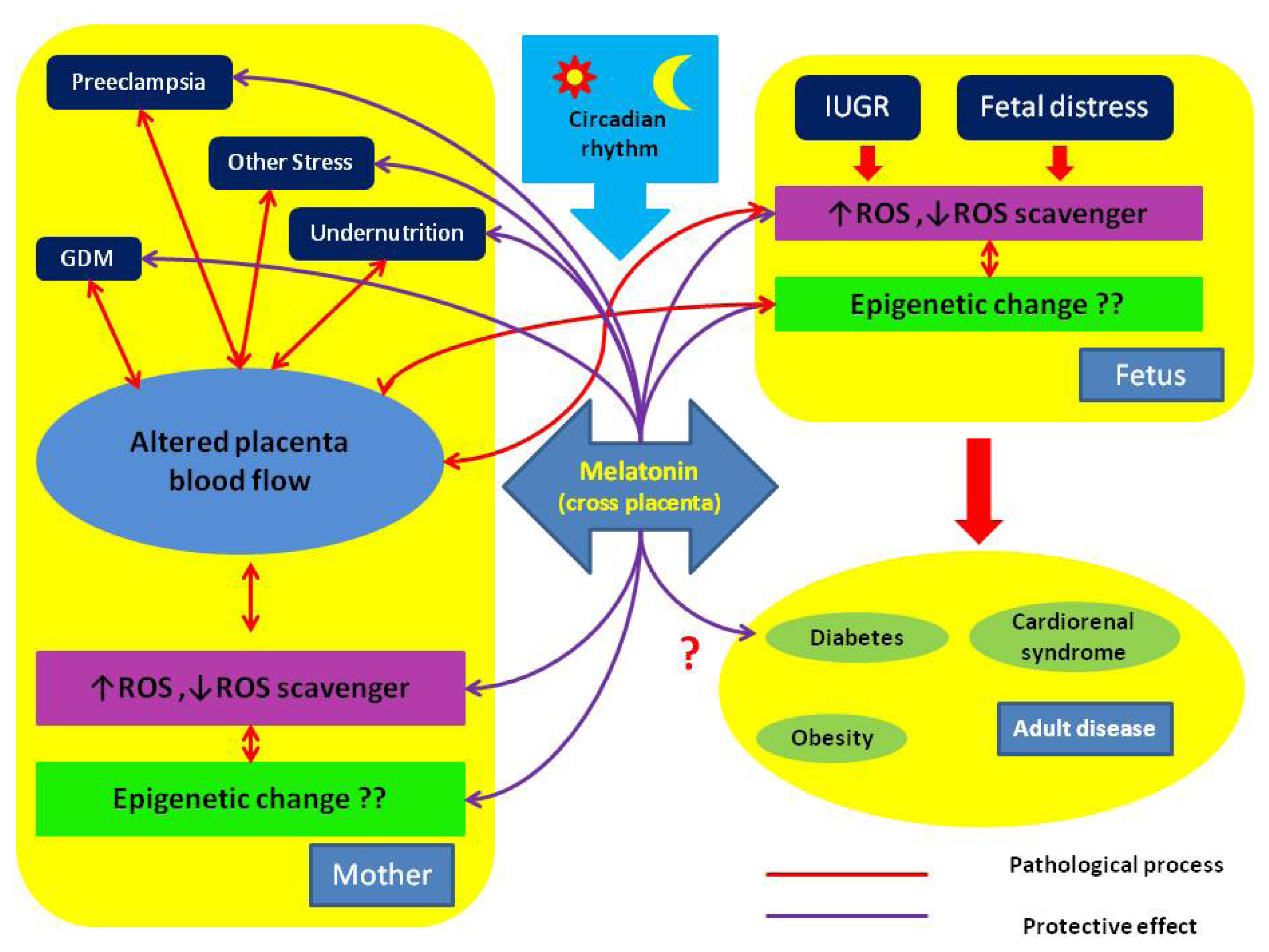{kind=link}
Abstract
:Abbreviations
| AGE | Advanced glycation end product |
| CREB | cAMP response element-binding protein |
| DNMT1 | DNA (cytosine-5)-methyltransferase 1 |
| GDM | gestational diabetis mellitus |
| GR | glucocorticoid receptor |
| IUGR | intrauterine growth retardation |
| MeCP2 | methyl CpG binding protein 2 |
| mGlu | metabotropic glutamate receptor |
| MT | melatonin receptor |
| NF-E2-related factor | nuclear factor erythroid 2-related factor 2 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| Pdx-1 | pancreatic and duodenal homeobox 1 |
| Per1 | period circadian protein homolog 1 |
| PPAR | peroxisome proliferator-activated receptor |
| ROS | reactive oxygen species |
| REM | rapid eye movement |
| SCN | suprachiasmatic nucleus |
1. Introduction
2. Melatonin Synthesis and Its Receptors
3. Melatonin Possesses Both Antioxidant and Epigenetic Modifications Abilities
4. Physiological Functions of Melatonin during Pregnancy
5. Safety Profiles and Side Effects of Melatonin in Pregnancy
6. The Concept of Fetal Programming
7. Redox and Epigenetic Mechanisms in Fetal Programming
7.1. The Role of Redox Alterations in Fetal Programming
7.2. The Role of Epigenetic Modifications in Fetal Programming
7.3. Redox Alterations in Utero Play a Role in Epigenetic Modifications
8. The Roles of Melatonin in Redox and Epigenetic Alterations in Fetal Programming
8.1. Melatonin Has a Role in Redox Modifications in Fetal Programming
8.2. Melatonin Has a Role in Epigenetic Modifications in Fetal Programming
9. The Roles of Melatonin in Compromised Pregnancies
9.1. Melatonin and GDM
9.2. Melatonin and Intrauterine Growth Restriction
9.3. Melatonin and Preeclampsia
9.4. Melatonin and Maternal Undernutrition
9.5. Melatonin and Maternal Stress
10. Conclusions
Acknowledgements
Conflict of Interest
References
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and its relation to the immune system and inflammation. Ann. N. Y. Acad. Sci 2000, 917, 376–386. [Google Scholar]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A pleiotropic molecule regulating inflammation. Biochem. Pharmacol 2010, 80, 1844–1852. [Google Scholar]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin-a pleiotropic, orchestrating regulator molecule. Prog. Neurobiol 2011, 93, 350–384. [Google Scholar]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res 2013, 54, 1–14. [Google Scholar]
- Chowdhury, I.; Sengupta, A.; Maitra, S.K. Melatonin: Fifty years of scientific journey from the discovery in bovine pineal gland to delineation of functions in human. Indian J. Biochem. Biophys 2008, 45, 289–304. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Osuna, C.; Gitto, E. Actions of melatonin in the reduction of oxidative stress. A review. J. Biomed. Sci 2000, 7, 444–458. [Google Scholar]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res 2004, 36, 1–9. [Google Scholar]
- Korkmaz, A.; Reiter, R.J. Epigenetic regulation: A new research area for melatonin? J. Pineal Res 2008, 44, 41–44. [Google Scholar]
- Korkmaz, A.; Rosales-Corral, S.; Reiter, R.J. Gene regulation by melatonin linked to epigenetic phenomena. Gene 2012, 503, 1–11. [Google Scholar]
- Mirick, D.K.; Davis, S. Melatonin as a biomarker of circadian dysregulation. Cancer Epidemiol. Biomarkers Prev 2008, 17, 3306–3313. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Burkhardt, S. Reactive oxygen and nitrogen species and cellular and organismal decline: amelioration with melatonin. Mech. Ageing Dev 2002, 123, 1007–1019. [Google Scholar]
- Cardinali, D.P.; Pagano, E.S.; Scacchi Bernasconi, P.A.; Reynoso, R.; Scacchi, P. Melatonin and mitochondrial dysfunction in the central nervous system. Horm. Behav. 2013. [Google Scholar] [CrossRef]
- Jan, J.E.; Freeman, R.D.; Fast, D.K. Melatonin treatment of sleep-wake cycle disorders in children and adolescents. Dev. Med. Child Neurol 1999, 41, 491–500. [Google Scholar]
- Dodge, N.N.; Wilson, G.A. Melatonin for treatment of sleep disorders in children with developmental disabilities. J. Child Neurol 2001, 16, 581–584. [Google Scholar]
- Okatani, Y.; Wakatsuki, A.; Reiter, R.J.; Miyahara, Y. Hepatic mitochondrial dysfunction in senescence-accelerated mice: Correction by long-term, orally administered physiological levels of melatonin. J. Pineal Res 2002, 33, 127–133. [Google Scholar]
- Gitto, E.; Aversa, S.; Reiter, R.J.; Barberi, I.; Pellegrino, S. Update on the use of melatonin in pediatrics. J. Pineal Res 2011, 50, 21–28. [Google Scholar]
- Chen, Y.C.; Tain, Y.L.; Sheen, J.M.; Huang, L.T. Melatonin utility in neonates and children. J. Formos. Med. Assoc 2012, 111, 57–66. [Google Scholar]
- Tamura, H.; Nakamura, Y.; Terron, M.P.; Flores, L.J.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and pregnancy in the human. Reprod. Toxicol 2008, 25, 291–303. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and reproduction revisited. Biol. Reprod 2009, 81, 445–456. [Google Scholar]
- Gitto, E.; Pellegrino, S.; Gitto, P.; Barberi, I.; Reiter, R.J. Oxidative stress of the newborn in the pre- and post-natal period and the clinical utility of melatonin. J. Pineal Res 2009, 46, 128–139. [Google Scholar]
- Aversa, S.; Pellegrino, S.; Barberi, I.; Reiter, R.J.; Gitto, E. Potential utility of melatonin as an antioxidant during pregnancy and in the perinatal period. J. Matern. Fetal Neonatal 2012, 25, 207–221. [Google Scholar]
- Harderland, R.; Poeggeler, B. Non-vertebrate melatonin. J. Pineal Res 2003, 34, 233–241. [Google Scholar]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol 2006, 38, 313–316. [Google Scholar]
- Facciola, G.; Hidestrand, M.; von Bahr, C.; Tybring, G. Cytochrome P450 isoforms involved in melatonin metabolism in human liver microsomes. Eur. J. Clin. Pharmacol 2001, 56, 881–888. [Google Scholar]
- Dubocovich, M.L.; Delagrange, P.; Krause, D.N.; Sugden, D.; Cardinali, D.P.; Olcese, J. International union of basic and clinical pharmacology. LXXV. Nomenclature, classification, and pharmacology of G protein-coupled melatonin receptors. Pharmacol. Rev 2010, 62, 343–380. [Google Scholar]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res 2009, 47, 109–126. [Google Scholar]
- Reiter, R.J.; Korkmaz, A.; Paredes, S.D.; Manchester, L.C.; Tan, D.X. Melatonin reduces oxidative/nitrosative stress due to drugs, toxins, metals, and herbicides. Neuro. Endocrinol. Lett 2008, 29, 609–613. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Qi, W.B.; Karbownik, M.; Calvo, J.R. Significance of melatonin in antioxidative defense system: Reactions and products. Biol. Signals Recept 2000, 9, 137–159. [Google Scholar]
- Acuña-Castroviejo, D.; Martín, M.; Macías, M.; Escames, G.; Leon, J.; Khaldy, H.; Reiter, R.J. Melatonin, mitochondria, and cellular bioenergetics. J. Pineal Res 2001, 30, 65–74. [Google Scholar]
- Korkmaz, A.; Sanchez-Barcelo, E.J.; Tan, D.X.; Reiter, R.J. Role of melatonin in the epigenetic regulation of breast cancer. Breast Cancer Res. Treat 2009, 115, 13–27. [Google Scholar]
- Sandyk, R.; Anastasiadis, P.G.; Anninos, P.A.; Tsagas, N. The pineal gland and spontaneous abortions: Implications for therapy with melatonin and magnetic field. Int. J. Neurosci 1992, 62, 243–250. [Google Scholar]
- Matsuzuka, T.; Sakamoto, N.; Ozawa, M.; Ushitani, A.; Hirabayashi, M.; Kanai, Y. Alleviation of maternal hyperthermia-induced early embryonic death by administration of melatonin to mice. J. Pineal Res 2005, 39, 217–223. [Google Scholar]
- Torres-Farfan, C.; Valenzuela, F.J.; Mondaca, M.; Valenzuela, G.J.; Krause, B.; Herrera, E.A.; Riquelme, R.; Llanos, A.J.; Seron-Ferre, M. Evidence of a role for melatonin in fetal sheep physiology: Direct actions of melatonin on fetal cerebral artery, brown adipose tissue and adrenal gland. J. Physiol 2008, 586, 4017–4027. [Google Scholar]
- Seron-Ferre, M.; Torres-Farfan, C.; Forcelledo, M.L.; Valenzuela, G.J. The development of circadian rhythms in the fetus and neonate. Semin. Perinatol 2001, 25, 363–370. [Google Scholar]
- Serón-Ferré, M.; Mendez, N.; Abarzua-Catalan, L.; Vilches, N.; Valenzuela, F.J.; Reynolds, H.E.; Llanos, A.J.; Rojas, A.; Valenzuela, G.J.; Torres-Farfan, C. Circadian rhythms in the fetus. Mol. Cell Endocrinol 2012, 349, 68–75. [Google Scholar]
- Reppert, S.M.; Weaver, D.R.; Rivkees, S.A.; Stopa, E.G. Putative melatonin receptor in a human biological clock. Science 1998, 242, 78–81. [Google Scholar]
- Wu, Y.H.; Zhou, J.N.; Balesar, R.; Unmehopa, U.; Bao, A.; Jockers, R.; Van Heerikhuize, J.; Swaab, D.F. Distribution of MT1 melatonin receptor immunoreactivity in the human hypothalamus and pituitary gland: Colocalization of MT1 with vasopressin, oxytocin, and corticotropin-releasing hormone. J. Comp. Neurol 2006, 499, 897–910. [Google Scholar]
- Simonneaux, V. Naughty melatonin: How mothers tick off their fetus. Endocrinology 2011, 152, 1734–1738. [Google Scholar]
- Karsch, F.J.; Bittman, E.L.; Foster, D.L.; Goodman, R.L.; Legan, S.J.; Robinson, J.E. Neuroendocrine basis of seasonal reproduction. Recent Prog. Horm. Res 1984, 40, 185–232. [Google Scholar]
- Malpaux, B.; Tricoire, H.; Mailliet, F.; Daveau, A.; Migaud, M.; Skinner, D.C.; Pelletier, J.; Chemineau, P. Melatonin and seasonal reproduction: Understanding the neuroendocrine mechanisms using the sheep as a model. Reprod. Suppl 2002, 59, 167–179. [Google Scholar]
- McMillen, I.C.; Houghton, D.C.; Young, I.R. Melatonin and the development of circadian and seasonal rhythmicity. J. Reprod. Fertil. Suppl 1995, 49, 137–146. [Google Scholar]
- Yellon, S.M.; Longo, L.D. Melatonin rhythms in fetal and maternal circulation during pregnancy in sheep. Am. J. Physiol 1987, 252, E799–E802. [Google Scholar]
- Deguchi, T. Ontogenesis of a biological clock for serotonin: acetyl coenzyme A N-acetyltransferase in pineal gland of rat. Proc. Natl. Acad. Sci. USA 1975, 72, 2814–2818. [Google Scholar]
- Yellon, S.M.; Longo, L.D. Effect of maternal pinealectomy and reverse photoperiod on the circadian melatonin rhythm in the sheep and fetus during the last trimester of pregnancy. Biol. Reprod 1988, 39, 1093–1099. [Google Scholar]
- Mcmillen, I.C.; Nowak, R. Maternal pinealectomy abolishes the diurnal rhythm in plasma melatonin concentrations in the fetal sheep and pregnant ewe during late gestation. J. Endocrinol 1989, 120, 459–464. [Google Scholar]
- Torres-Farfan, C.; Richter, H.G.; Germain, A.M.; Valenzuela, G.J.; Campino, C.; Rojas-García, P.; Forcelledo, M.L.; Torrealba, F.; Serón-Ferré, M. Maternal melatonin selectively inhibits cortisol production in the primate fetal adrenal gland. J. Physiol 2004, 554, 841–856. [Google Scholar]
- Bellavía, S.L.; Carpentieri, A.R.; Vaqué, A.M.; Macchione, A.F.; Vermouth, N.T. Pup circadian rhythm entrainment-effect of maternal ganglionectomy or pinealectomy. Physiol. Behav 2006, 89, 342–349. [Google Scholar]
- Mendez, N.; Abarzua-Catalan, L.; Vilches, N.; Galdames, H.A.; Spichiger, C.; Richter, H.G.; Valenzuela, G.J.; Seron-Ferre, M.; Torres-Farfan, C. Timed maternal melatonin treatment reverses circadian disruption of the fetal adrenal clock imposed by exposure to constant light. PLoS One 2012, 7, e42713. [Google Scholar]
- Chan, W.Y.; Ng, T.B. Development of pre-implantation mouse embryos under the influence of pineal indoles. J. Neural Transm. Gen. Sect 1994, 96, 19–29. [Google Scholar]
- McElhinny, A.S.; Davis, F.C.; Warner, C.M. The effect of melatonin on cleavage rate of C57BL/6 and CBA/Ca preimplantation embryos cultured in vitro. J. Pineal Res 1996, 21, 44–48. [Google Scholar]
- Jahnke, G.; Marr, M.; Myers, C.; Wilson, R.; Travlos, G.; Price, C. Maternal and developmental toxicity evaluation of melatonin administered orally to pregnant Sprague-Dawley rats. Toxicol. Sci 1999, 50, 271–279. [Google Scholar]
- Sadowsky, D.W.; Yellon, S.; Mitchell, M.D.; Nathanielsz, P.W. Lack of effect of melatonin on myometrial electromyographic activity in the pregnant sheep at 138–142 days gestation (term = 147 days gestation). Endocrinology 1991, 128, 1812–1818. [Google Scholar]
- Leach, C.M.; Thorburn, G.D. A comparison of the inhibitory effects of melatonin and indomethacin on platelet aggregation and thromboxane release. Prostaglandins 1980, 20, 51–56. [Google Scholar]
- Barker, D.J.; Winter, P.D.; Osmond, C.; Margetts, B.; Simmonds, S.J. Weight in infancy and death from ischaemic heart disease. Lancet 1989, 2, 577–580. [Google Scholar]
- Chen, M.; Zhang, L. Epigenetic mechanisms in developmental programming of adult disease. Drug Discov. Today 2011, 16, 1007–1018. [Google Scholar]
- Hanson, M.; Godfrey, K.M.; Lillycrop, K.A.; Burdge, G.C.; Gluckman, P.D. Developmental plasticity and developmental origins of non-communicable disease: Theoretical considerations and epigenetic mechanisms. Prog. Biophys. Mol. Biol 2011, 106, 272–280. [Google Scholar]
- Hochberg, Z.; Feil, R.; Constancia, M.; Fraga, M.; Junien, C.; Carel, J.C.; Boileau, P.; Le Bouc, Y.; Deal, C.L.; Lillycrop, K.; et al. Child health, developmental plasticity, and epigenetic programming. Endocr. Rev 2011, 32, 159–224. [Google Scholar]
- Thompson, L.P.; Al-Hasan, Y. Impact of oxidative stress in fetal programming. J. Pregnancy 2012, 2012, 582748. [Google Scholar]
- Xiong, F.; Zhang, L. Role of the hypothalamic-pituitary-adrenal axis in developmental programming of health and disease. Front. Neuroendocrinol 2012, 12, 62–63. [Google Scholar]
- Luo, Z.C.; Fraser, W.D.; Julien, P.; Deal, C.L.; Audibert, F.; Smith, G.N.; Xiong, X.; Walker, M. Tracing the origins of “fetal origins” of adult diseases: Programming by oxidative stress? Med. Hypotheses 2006, 66, 38–44. [Google Scholar]
- Luo, Z.C.; Xiao, L.; Nuyt, A.M. Mechanisms of developmental programming of the metabolic syndrome and related disorders. World J. Diabetes 2010, 1, 89–98. [Google Scholar]
- Tarry-Adkins, J.L.; Ozanne, S.E. Mechanisms of early life programming: Current knowledge and future directions. Am. J. Clin. Nutr 2011, 94, 1765S–1771S. [Google Scholar]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; et al. The role of melatonin as an antioxidant in the follicle. J. Ovarian Res 2012, 5, 5. [Google Scholar]
- Miller, S.L.; Wallace, E.M.; Walker, D.W. Antioxidant therapies: A potential role in perinatal medicine. Neuroendocrinology 2012, 96, 13–23. [Google Scholar]
- Nelissen, E.C.; van Montfoort, A.P.; Dumoulin, J.C.; Evers, J.L. Epigenetics and the placenta. Hum. Reprod. Update 2011, 17, 397–417. [Google Scholar]
- Huang, L.T.; Hsieh, C.S.; Chang, K.A.; Tain, Y.L. Roles of nitric oxide and asymmetric dimethylarginine in pregnancy and fetal programming. Int. J. Mol. Sci 2012, 13, 14606–14622. [Google Scholar]
- Ribatti, D.; Nico, B.; Crivellato, E. Morphological and molecular aspects of physiological vascular morphogenesis. Angiogenesis 2009, 12, 101–111. [Google Scholar]
- Dokras, A.; Coffin, J.; Field, L.; Frakes, A.; Lee, H.; Madan, A.; Nelson, T.; Ryu, G.Y.; Yoon, J.G.; Madan, A. Epigenetic regulation of maspin expression in the human placenta. Mol. Hum. Reprod 2006, 12, 611–617. [Google Scholar]
- Longtine, M.S.; Nelson, D.M. Placental dysfunction and fetal programming: The importance of placental size, shape, histopathology, and molecular composition. Semin. Reprod. Med 2011, 29, 187–196. [Google Scholar]
- Yuen, R.K.; Chen, B.; Blair, J.D.; Robinson, W.P.; Nelson, D.M. Hypoxia alters the epigenetic profile in cultured human placental trophoblasts. Epigenetics 2013, 8, 192–202. [Google Scholar]
- Rugg-Gunn, P.J. Epigenetic features of the mouse trophoblast. Reprod. Biomed. Online 2012, 25, 21–30. [Google Scholar]
- Cerda, S.; Weitzman, S.A. Influence of oxygen radical injury onDNA methylation. Mutat. Res 1997, 386, 141–152. [Google Scholar]
- Hitchler, M.J.; Domann, F.E. An epigenetic perspective on the free radical theory of development. Free Radic. Biol. Med 2007, 43, 1023–1036. [Google Scholar]
- Ziech, D.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Reactive Oxygen Species (ROS)-induced genetic and epigenetic alterations in human carcinogenesis. Mutat. Res 2011, 711, 167–173. [Google Scholar]
- Brzezinski, A.; Seibel, M.M.; Lynch, H.J.; Deng, M.H.; Wurtman, R.J. Melatonin in human preovulatory follicular fluid. J. Clin. Endocrinol. Metab. 1987, 64, 865–867. [Google Scholar]
- Okatani, Y.; Wakatsuki, A.; Shinohara, K.; Kaneda, C.; Fukaya, T. Melatonin stimulates glutathione peroxidase activity in human chorion. J. Pineal Res 2001, 30, 199–205. [Google Scholar]
- Zhu, J.L.; Hjollund, N.H.; Andersen, A.M.; Olsen, J. Shift work, job stress, and late fetal loss: The National Birth Cohort in Denmark. J. Occup. Environ. Med 2004, 46, 1144–1149. [Google Scholar]
- Croteau, A.; Marcoux, S.; Brisson, C. Work activity in pregnancy, preventive measures, and the risk of delivering a small-for-gestational-age infant. Am. J. Public Health 2006, 96, 846–855. [Google Scholar]
- Ferreira, D.S.; Amaral, F.G.; Mesquita, C.C.; Barbosa, A.P.; Lellis-Santos, C.; Turati, A.O.; Santos, L.R.; Sollon, C.S.; Gomes, P.R.; Faria, J.A.; et al. Maternal melatonin programs the daily pattern of energy metabolism in adult offspring. PLoS One 2012, 7, e38795. [Google Scholar]
- McCurdy, C.E.; Bishop, J.M.; Williams, S.M.; Grayson, B.E.; Smith, M.S.; Friedman, J.E.; Grove, K.L. Maternal high-fat diet triggers lipotoxicity in the fetal livers of nonhuman primates. J. Clin. Invest 2009, 119, 323–335. [Google Scholar]
- Suter, M.; Bocock, P.; Showalter, L.; Hu, M.; Shope, C.; McKnight, R.; Grove, K.; Lane, R.; Aagaard-Tillery, K. Epigenomics: Maternal high-fat diet exposure in utero disrupts peripheral circadian gene expression in nonhuman primates. FASEB J 2011, 25, 714–726. [Google Scholar]
- Sun, Z.; Chin, Y.E.; Zhang, D.D. Acetylation of Nrf2 by p300/CBP augments promoter-specific DNA binding of Nrf2 during the antioxidant response. Mol. Cell. Biol 2009, 29, 2658–2672. [Google Scholar]
- Kawai, Y.; Garduno, L.; Theodore, M.; Yang, J.; Arinze, I.J. Acetylation-deacetylation of the transcription factor Nrf2 (nuclear factor erythroid 2-related factor 2) regulates its transcriptional activity and nucleocytoplasmic localization. J. Biol. Chem 2011, 286, 7629–7640. [Google Scholar]
- Kennaway, D.J. Programming of the fetal suprachiasmatic nucleus and subsequent adult rhythmicity. Trends Endocrinol. Metab 2002, 13, 398–402. [Google Scholar]
- Cagampang, F.R.; Poore, K.R.; Hanson, M.A. Developmental origins of the metabolic syndrome: Body clocks and stress responses. Brain Behav. Immun 2011, 25, 214–220. [Google Scholar]
- Kennaway, D.J.; Goble, F.C.; Stamp, G.E. Factors influencing the development of melatonin rhythmicity in humans. J. Clin. Endocrinol. Metab 1996, 81, 1525–1532. [Google Scholar]
- Holditch-Davis, D.; Edwards, L.J. Temporal organization of sleep-wake states in preterm infants. Dev. Psychobiol 1998, 33, 257–269. [Google Scholar]
- Mirmiran, M.; Maas, Y.G.; Ariagno, R.L. Development of fetal and neonatal sleep and circadian rhythms. Sleep Med. Rev 2003, 7, 321–334. [Google Scholar]
- Muñoz-Hoyos, A.; Rodriguez-Cabezas, T.; Molina-Carballo, A.; Martinez-Sempere, J.J.; Ruiz-Cosano, C.; Acuña-Castroviejo, D. Melatonin concentration in the umbilical artery and vein in human preterm and term neonates and neonates with acute fetal distress. J. Pineal Res 1992, 13, 184–191. [Google Scholar]
- Simmons, D. Epidemiology of Diabetes in Pregnancy. In Practical Management of Diabetes in Pregnancy; McCance, D., Maresh, M., Eds.; Blackwell Publishing: London, UK, 2010. [Google Scholar]
- Muñoz-Hoyos, A.; Bonillo-Perales, A.; Avila-Villegas, R.; González-Ripoll, M.; Uberos, J.; Florido-Navío, J.; Molina-Carballo, A. Melatonin levels during the first week of life and their relation with the antioxidant response in the perinatal period. Neonatology 2007, 92, 209–216. [Google Scholar]
- Saito, S.; Tachibana, T.; Choi, Y.H.; Denbow, D.M.; Furuse, M. ICV melatonin reduces acute stress responses in neonatal chicks. Behav. Brain Res 2005, 165, 197–203. [Google Scholar]
- Owens, J.A.; Witmans, M. Sleep problems. Curr. Probl. Pediatr. Adolesc. Health Care 2004, 34, 154–179. [Google Scholar]
- Chung, S.S.; Ho, E.C.; Lam, K.S.; Chung, S.K. Contribution of polyol pathway to diabetes-induced oxidative stress. J. Am. Soc. Nephrol 2003, 14, S233–S236. [Google Scholar]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar]
- Gao, L.; Mann, G.E. Vascular NAD(P)H oxidase activation in diabetes: A double-edged sword in redox signalling. Cardiovasc. Res 2009, 82, 9–20. [Google Scholar]
- Brownlee, M. The pathological implications of protein glycation. Clin. Invest. Med 1995, 18, 275–281. [Google Scholar]
- Agil, A.; Navarro-Alarcón, M.; Ruiz, R.; Abuhamadah, S.; El-Mir, M.Y.; Vázquez, G.F. Beneficial effects of melatonin on obesity and lipid profile in young zucker diabetic fatty rats. J. Pineal Res 2011, 50, 207–212. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Fuentes-Broto, L.; Paredes, S.D.; Reiter, R.J. Significance and application of melatonin in the regulation of brown adipose tissue metabolism: Relation to human obesity. Obes. Rev 2011, 12, 167–188. [Google Scholar]
- Liao, S.; Liu, Y.; Tan, Y.; Gan, L.; Mei, J.; Song, W.; Chi, S.; Dong, X.; Chen, X.; Deng, S. Association of genetic variants of melatonin receptor 1B with gestational plasma glucose level and risk of glucose intolerance in pregnant Chinese women. PLoS One 2012, 7, e40113. [Google Scholar]
- Nistala, R.; Hayden, M.R.; Demarco, V.G.; Henriksen, E.J.; Lackland, D.T.; Sowers, J.R. Prenatal programming and epigenetics in the genesis of the cardiorenal syndrome. Cardiorenal. Med 2011, 1, 243–254. [Google Scholar]
- Garcia-Vargas, L.; Addison, S.S.; Nistala, R.; Kurukulasuriya, D.; Sowers, J.R. Gestational diabetes and the offspring: implications in the development of the cardiorenal metabolic syndrome in offspring. Cardiorenal. Med 2012, 2, 134–142. [Google Scholar]
- Miehle, K.; Stephan, H.; Fasshauer, M. Leptin, adiponectin, and other adipokines in gestational diabetes mellitus and preeclampsia. Clin. Endocrinol 2012, 76, 2–11. [Google Scholar]
- Mandruzzato, G.; Antsaklis, A.; Botet, F.; Chervenak, F.A.; Figueras, F.; Grunebaum, A.; Puerto, B.; Skupski, D.; Stanojevic, M. Intrauterine restriction (IUGR). J. Perinat. Med 2008, 36, 277–281. [Google Scholar]
- Forsén, T.; Eriksson, J.; Tuomilehto, J.; Reunanen, A.; Osmond, C.; Barker, D. The fetal and childhood growth of persons who develop type 2 diabetes. Ann. Intern. Med 2000, 133, 176–182. [Google Scholar]
- Pilgaard, K.; Hammershaimb Mosbech, T.; Grunnet, L.; Eiberg, H.; Van Hall, G.; Fallentin, E.; Larsen, T.; Larsen, R.; Poulsen, P.; Vaag, A. Differential nongenetic impact of birth weight versus third-trimester growth velocity on glucose metabolism and magnetic resonance imaging abdominal obesity in young healthy twins. J. Clin. Endocrinol. Metab 2011, 96, 2835–2843. [Google Scholar]
- Painter, R.C.; de Rooij, S.R.; Bossuyt, P.M.; Simmers, T.A.; Osmond, C.; Barker, D.J.; Bleker, O.P.; Roseboom, T.J. Early onset of coronary artery disease after prenatal exposure to the Dutch famine. Am. J. Clin. Nutr 2006, 84, 322–327. [Google Scholar]
- Karowicz-Bilinska, A.; Kedziora-Kornatowska, K.; Bartosz, G. Indices of oxidative stress in pregnancy with fetal growth restriction. Free Radic. Res 2007, 41, 870–873. [Google Scholar]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Invest 2007, 64, 187–192. [Google Scholar]
- Mohn, A.; Chiavaroli, V.; Cerruto, M.; Blasetti, A.; Giannini, C.; Bucciarelli, T.; Chiarelli, F. Increased oxidative stress in prepubertal children born small for gestational age. J. Clin. Endocrinol. Metab 2007, 92, 1372–1378. [Google Scholar]
- Cambonie, G.; Comte, B.; Yzydorczyk, C.; Ntimbane, T.; Germain, N.; Le, N.L.; Pladys, P.; Gauthier, C.; Lahaie, I.; Abran, D.; et al. Antenatal antioxidant prevents adult hypertension, vascular dysfunction, and microvascular rarefaction associated with in utero exposure to a low-protein diet. Am. J. Physiol. Regul. Integr. Comp. Physiol 2007, 292, R1236–R1245. [Google Scholar]
- Maggioni, C.; Cornelissen, G.; Antinozzi, R.; Ferrario, M.; Grafe, A.; Halberg, F. A half-yearly aspect of circulating melatonin in pregnancies complicated by intrauterine growth retardation. Neuro. Endocrinol. Lett 1999, 20, 55–68. [Google Scholar]
- Lemley, C.O.; Meyer, A.M.; Camacho, L.E.; Neville, T.L.; Newman, D.J.; Caton, J.S.; Vonnahme, K.A. Melatonin supplementation alters uteroplacental hemodynamics and fetal development in an ovine model of intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol 2012, 302, R454–467. [Google Scholar]
- Gheorghe, C.P.; Goyal, R.; Mittal, A.; Longo, L.D. Gene expression in the placenta: Maternal stress and epigenetic responses. Int. J. Dev. Biol 2010, 54, 507–523. [Google Scholar]
- Fu, Q.; McKnight, R.A.; Yu, X.; Callaway, C.W.; Lane, R.H. Growth retardation alters the epigenetic characteristics of hepatic dual specificity phosphatase 5. FASEB J 2006, 20, 2127–2129. [Google Scholar]
- Lillycrop, K.A.; Slater-Jefferies, J.L.; Hanson, M.A.; Godfrey, K.M.; Jackson, A.A.; Burdge, G.C. Induction of altered epigenetic regulation of the hepatic glucocorticoid receptor in the offspring of rats fed a protein-restricted diet during pregnancy suggests that reduced DNA methyltransferase-1 expression is involved in impaired DNA methylation and changes in histone modifications. Br. J. Nutr 2007, 97, 1064–1073. [Google Scholar]
- Patterson, A.J.; Xiao, D.; Xiong, F.; Dixon, B.; Zhang, L. Hypoxia-derived oxidative stress mediates epigenetic repression of PKCɛ gene in foetal rat hearts. Cardiovasc. Res 2012, 93, 302–310. [Google Scholar]
- Pinney, S.E.; Simmons, R.A. Metabolic programming, epigenetics, and gestational diabetes mellitus. Curr. Diab. Rep 2012, 12, 67–74. [Google Scholar]
- Bogdarina, I.; Haase, A.; Langley-Evans, S.; Clark, A.J.L. Glucocorticoid effects on the programming of AT1b angiotensin receptor gene methylation and expression in the rat. PLoS One 2010, 5, e9237. [Google Scholar]
- Rexhaj, E.; Bloch, J.; Jayet, P.Y.; Rimoldi, S.F.; Dessen, P.; Mathieu, C.; Tolsa, J.F.; Nicod, P.; Scherrer, U.; Sartori, C. Fetal programming of pulmonary vascular dysfunction in mice: Role of epigenetic mechanisms. Am. J. Physiol. Heart Circ. Physiol 2011, 301, H247–H252. [Google Scholar]
- Ke, X.; Lei, Q.; James, S.J.; Kelleher, S.L.; Melnyk, S.; Jernigan, S.; Yu, X.; Wang, L.; Callaway, C.W.; Gill, G.; et al. Uteroplacental insufficiency affects epigenetic determinants of chromatin structure in brains of neonatal and juvenile IUGR rats. Physiol. Genom 2006, 25, 16–28. [Google Scholar]
- Raychaudhuri, N.; Raychaudhuri, S.; Thamotharan, M.; Devaskar, S.U. Histone code modifications repress glucose transporter 4 expression in the intrauterine growth-restricted offspring. J. Biol. Chem 2008, 283, 13611–13626. [Google Scholar]
- Noris, M.; Perico, N.; Remuzzi, G. Mechanisms of disease: Pre-eclampsia. Nat. Clin. Pract. Nephrol 2005, 1, 98–114. [Google Scholar]
- Simmons, R.A. Developmental origins of adult disease. Pediatr. Clin. North. Am 2009, 56, 449–466. [Google Scholar]
- Seabra, M.L.; Bignotto, M.; Pinto, L.R., Jr; Tufik, S. Randomized, double-blind clinical trial, controlled with placebo, of the toxicology of chronic melatonin treatment. J. Pineal Res. 2000, 29, 193–200. [Google Scholar]
- Gitto, E.; Reiter, R.J.; Amodio, A.; Romeo, C.; Cuzzocrea, E.; Sabatino, G.; Buonocore, G.; Cordaro, V.; Trimarchi, G.; Barberi, I. Early indicators of chronic lung disease in preterm infants with respiratory distress syndrome and their inhibition by melatonin. J. Pineal Res 2004, 36, 250–255. [Google Scholar]
- Lanoix, D.; Guérin, P.; Vaillancourt, C. Placental melatonin production and melatonin receptor expression are altered in preeclampsia: New insights into the role of this hormone in pregnancy. J. Pineal Res 2012, 53, 417–425. [Google Scholar]
- Gordon, N. The therapeutics of melatonin: A paediatric perspective. Brain Dev 2000, 22, 213–217. [Google Scholar]
- Kulkarni, A.; Chavan-Gautam, P.; Mehendale, S.; Yadav, H.; Joshi, S. Global DNA methylation patterns in placenta and its association with maternal hypertension in pre-eclampsia. DNA Cell Biol 2011, 30, 79–84. [Google Scholar]
- Chim, S.S.; Tong, Y.K.; Chiu, R.W.; Lau, T.K.; Leung, T.N.; Chan, L.Y.; Oudejans, C.B.; Ding, C.; Lo, Y.M. Detection of the placental epigenetic signature of the maspin gene in maternal plasma. Proc. Natl. Acad. Sci. USA 2005, 102, 14753–14758. [Google Scholar]
- Fowden, A.L.; Giussani, D.A.; Forhead, A.J. Intrauterine programming of physiological systems: Causes and consequences. Physiology (Bethesda) 2006, 21, 29–37. [Google Scholar]
- Gupta, P.; Narang, M.; Banerjee, B.D.; Basu, S. Oxidative stress in term small for gestational age neonates born to undernourished mothers: A case control study. BMC Pediatr 2004, 4, 14. [Google Scholar]
- Richter, H.G.; Hansell, J.A.; Raut, S.; Giussani, D.A. Melatonin improves placental efficiency and birth weight and increases the placental expression of antioxidant enzymes in undernourished pregnancy. J. Pineal Res 2009, 46, 357–364. [Google Scholar]
- Nagai, R.; Watanabe, K.; Wakatsuki, A.; Hamada, F.; Shinohara, K.; Hayashi, Y.; Imamura, R.; Fukaya, T. Melatonin preserves fetal growth in rats by protecting against ischemia/reperfusion-induced oxidative/nitrosative mitochondrial damage in the placenta. J. Pineal Res 2008, 45, 271–276. [Google Scholar]
- Duran, P.; Cintra, L.; Galler, J.R.; Tonkiss, J. Prenatal protein malnutrition induces a phase shift advance of the spontaneous locomotor rhythm and alters the rest/activity ratio in adult rats. Nutr. Neurosci 2005, 8, 167–172. [Google Scholar]
- Stevens, A.; Begum, G.; Cook, A.; Connor, K.; Rumball, C.; Oliver, M.; Challis, J.; Bloomfield, F.; White, A. Epigenetic changes in the hypothalamic proopiomelanocortin and glucocorticoid receptor genes in the ovine fetus after periconceptional undernutrition. Endocrinology 2010, 151, 3652–3664. [Google Scholar]
- Lillycrop, K.A.; Phillips, E.S.; Jackson, A.A.; Hanson, M.A.; Burdge, G.C. Dietary protein restriction in the pregnant rat induces altered epigenetic regulation of the glucocorticoid receptor and peroxisomal proliferator-activated receptor alpha in the heart of the offspring which is prevented by folic acid. Proc. Nutr. Soc 2006, 65, 65A. [Google Scholar]
- Burdge, G.C.; Hanson, M.A.; Slater-Jefferies, J.L.; Lillycrop, K.A. Epigenetic regulation of transcription: A mechanism for inducing variations in phenotype (fetal programming) by differences in nutrition during early life? Br. J. Nutr 2007, 97, 1036–1046. [Google Scholar]
- Li, Y.; Gonzalez, P.; Zhang, L. Fetal stress and programming of hypoxic/ischemic-sensitive phenotype in the neonatal brain: Mechanisms and possible interventions. Prog. Neurobiol 2012, 98, 145–165. [Google Scholar]
- Lui, C.C.; Wang, J.Y.; Tain, Y.L.; Chen, Y.C.; Chang, K.A.; Lai, M.C.; Huang, L.T. Prenatal stress in rat causes long-term spatial memory deficit and hippocampus MRI abnormality: Differential effects of postweaning enriched environment. Neurochem. Int 2011, 58, 434–441. [Google Scholar]
- Weinstock, M. Alterations induced by gestational stress in brain morphology and behaviour of the offspring. Prog. Neurobiol 2001, 65, 427–451. [Google Scholar]
- Dugovic, C.; Maccari, S.; Weibel, L.; Turek, F.W.; Van Reeth, O. High corticosterone levels in prenatally stressed rats predict persistent paradoxical sleep alterations. J. Neurosci 1999, 19, 8656–8664. [Google Scholar]
- Mairesse, J.; Silletti, V.; Laloux, C.; Zuena, A.R.; Giovine, A.; Consolazione, M.; van Camp, G.; Malagodi, M.; Gaetani, S.; Cianci, S.; et al. Chronic agomelatine treatment corrects the abnormalities in the circadian rhythm of motor activity and sleep/wake cycle induced by prenatal restraint stress in adult rats. Int. J. Neuropsychopharmacol 2012, 6, 1–16. [Google Scholar]
- Basta, M.; Chrousos, G.P.; Vela-Bueno, A.; Vgontzas, A.N. Chronic insomnia and stress system. Sleep Med. Clin 2007, 2, 279–291. [Google Scholar]
- Allan, S.M.; Rothwell, N.J. Cytokines and acute neurodegeneration. Nat. Rev. Neurosci 2001, 2, 734–744. [Google Scholar]
- Soumyarani, V.S.; Jayakumari, N. Oxidatively modified high density lipoprotein promotes inflammatory response in human monocytes-macrophages by enhanced production of ROS, TNF-α, MMP-9, and MMP-2. Mol. Cell Biochem 2012, 366, 277–285. [Google Scholar]
- Darnaudéry, M.; Maccari, S. Epigenetic programming of the stress response in male and female rats by prenatal restraint stress. Brain Res. Rev 2008, 57, 571–585. [Google Scholar]
- Morley-Fletcher, S.; Mairesse, J.; Soumier, A.; Banasr, M.; Fagioli, F.; Gabriel, C.; Mocaer, E.; Daszuta, A.; McEwen, B.; Nicoletti, F.; et al. Chronic agomelatine treatment corrects behavioral, cellular, and biochemical abnormalities induced by prenatal stress in rats. Psychopharmacology (Berl) 2011, 217, 301–313. [Google Scholar]
- Matrisciano, F.; Tueting, P.; Maccari, S.; Nicoletti, F.; Guidotti, A. Pharmacological activation of group-II metabotropic glutamate receptors corrects a schizophrenia-like phenotype induced by prenatal stress in mice. Neuropsychopharmacology 2012, 37, 929–938. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Y.-C.; Sheen, J.-M.; Tiao, M.-M.; Tain, Y.-L.; Huang, L.-T. Roles of Melatonin in Fetal Programming in Compromised Pregnancies. Int. J. Mol. Sci. 2013, 14, 5380-5401. https://doi.org/10.3390/ijms14035380
Chen Y-C, Sheen J-M, Tiao M-M, Tain Y-L, Huang L-T. Roles of Melatonin in Fetal Programming in Compromised Pregnancies. International Journal of Molecular Sciences. 2013; 14(3):5380-5401. https://doi.org/10.3390/ijms14035380
Chicago/Turabian StyleChen, Yu-Chieh, Jiunn-Ming Sheen, Miao-Meng Tiao, You-Lin Tain, and Li-Tung Huang. 2013. "Roles of Melatonin in Fetal Programming in Compromised Pregnancies" International Journal of Molecular Sciences 14, no. 3: 5380-5401. https://doi.org/10.3390/ijms14035380
APA StyleChen, Y. -C., Sheen, J. -M., Tiao, M. -M., Tain, Y. -L., & Huang, L. -T. (2013). Roles of Melatonin in Fetal Programming in Compromised Pregnancies. International Journal of Molecular Sciences, 14(3), 5380-5401. https://doi.org/10.3390/ijms14035380