A Novel Moderate Constitutive Promoter Derived from Poplar (Populus tomentosa Carrière)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Cloning and Sequence Analysis of the PtMCP Promoter
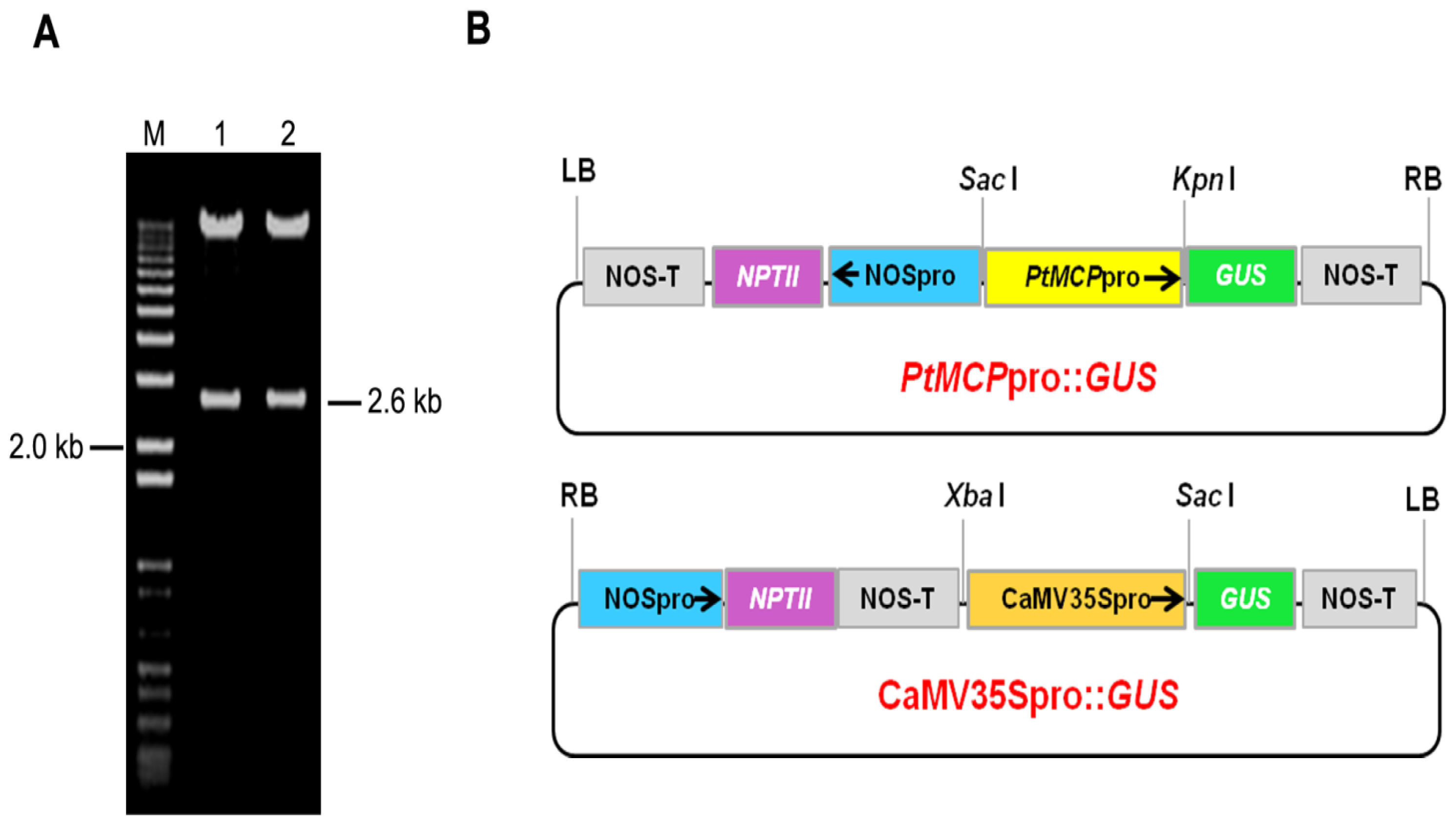
2.1.2. Construction and Transformation of the PtMCP Promoter::GUS Construct into Arabidopsis
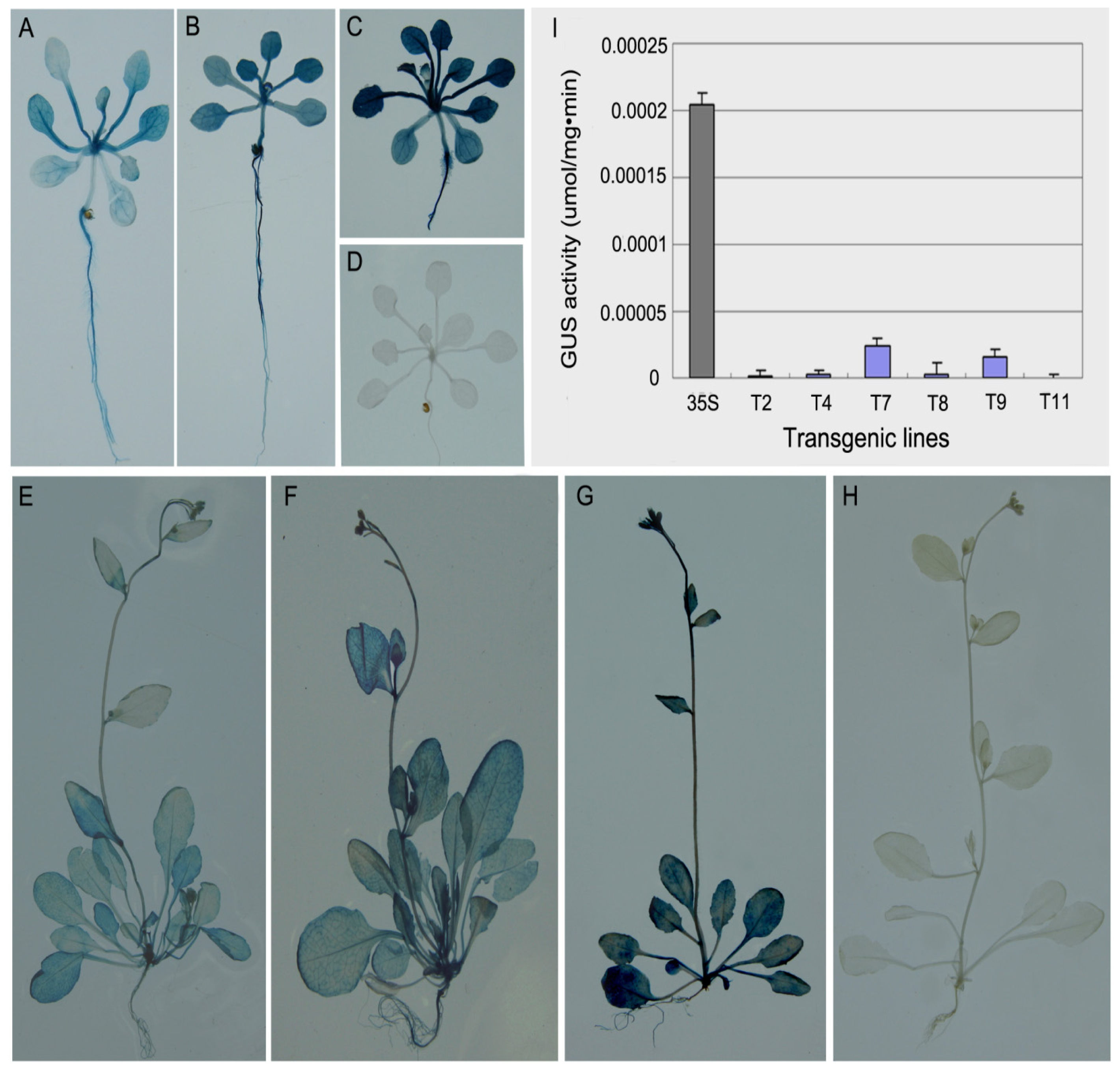
2.1.3. Characterization and Analysis of Transgenic Arabidopsis
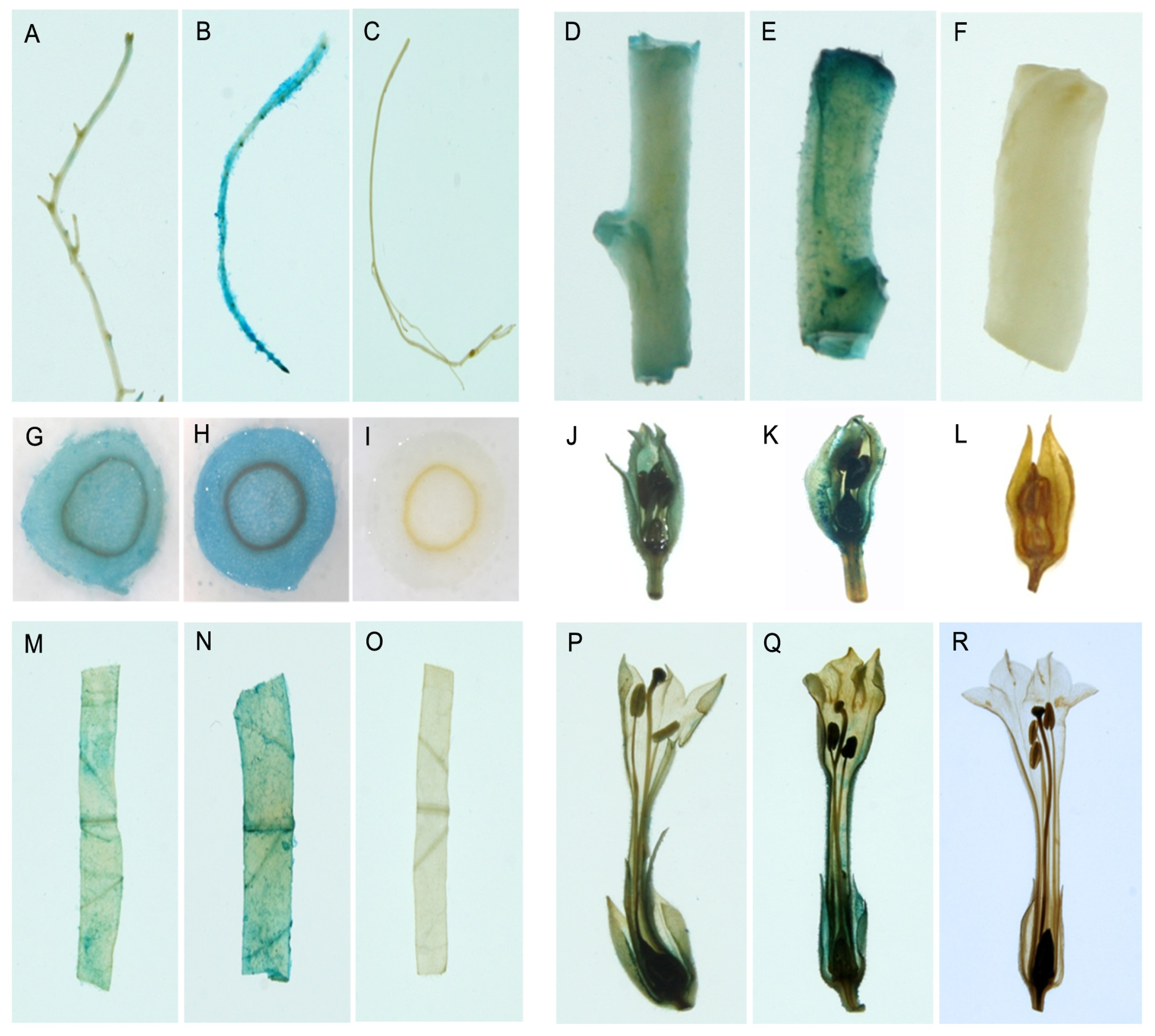
2.1.4. Histochemical and Fluorometric GUS Assays
2.1.5. Transient Expression of the PtMCP Promoter in Tobacco
2.2. Discussion
3. Experimental Section
3.1. Plant Materials and Growth Conditions
3.2. Bacterial Strains
3.3. Construction of the PtMCP Promoter::GUS Plasmid
3.4. Arabidopsis Transformation
3.5. Genomic PCR and RT-PCR Assays
3.6. Histochemical and Fluorometric GUS Assays
3.7. Agrobacterium-Mediated Transient Expression in Tobacco
4. Conclusions
Supplementary Information
ijms-14-06187-s001.docxAcknowledgments
Conflict of Interest
References
- Odell, J.T.; Nagy, F.; Chua, N.H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 1985, 313, 810–812. [Google Scholar]
- Battraw, M.J.; Hall, T.C. Histochemical analysis of CaMV 35S promoter-beta-glucuronidase gene expression in transgenic rice plants. Plant Mol. Biol 1990, 15, 527–538. [Google Scholar]
- Benfey, P.N.; Ren, L.; Chua, N.H. Tissue-specific expression from CaMV 35S enhancer subdomains in early stages of plant development. EMBO J 1990, 9, 1677–1684. [Google Scholar]
- Datla, R.S.S.; Bekkaoui, F.; Hammerlindl, J.K.; Pilate, G.; Dunstan, D.I.; Crosby, W.L. Improved high-level constitutive foreign gene expression in plants using an AMV RNA4 untranslated leader sequence. Plant Sci 1993, 94, 139–149. [Google Scholar]
- Holtorf, S.; Apel, K.; Bohlmann, H. Comparison of different constitutive and inducible promoters for the overexpression of transgenes in Arabidopsis thaliana. Plant Mol. Biol 1995, 29, 637–646. [Google Scholar]
- Wilmink, A.; van de Ven, B.C.; Dons, J.J. Activity of constitutive promoters in various species from the Liliaceae. Plant Mol. Biol 1995, 28, 949–955. [Google Scholar]
- Christensen, A.H.; Sharrock, R.A.; Quail, P.H. Maize polyubiquitin genes: Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol 1992, 18, 675–689. [Google Scholar]
- Cornejo, M.J.; Luth, D.; Blankenship, K.M.; Anderson, O.D.; Blechl, A.E. Activity of a maize ubiquitin promoter in transgenic rice. Plant Mol. Biol 1993, 23, 567–581. [Google Scholar]
- Lu, J.; Sivamani, E.; Li, X.; Qu, R. Activity of the 5′ regulatory regions of the rice polyubiquitin rubi3 gene in transgenic rice plants as analyzed by both GUS and GFP reporter genes. Plant Cell Rep 2008, 27, 1587–1600. [Google Scholar]
- Chiera, J.M.; Bouchard, R.A.; Dorsey, S.L.; Park, E.; Buenrostro-Nava, M.T.; Ling, P.P.; Finer, J.J. Isolation of two highly active soybean (Glycine max (L.) Merr.) promoters and their characterization using a new automated image collection and analysis system. Plant Cell Rep 2007, 26, 1501–1509. [Google Scholar]
- Hernandez-Garcia, C.M.; Martinelli, A.P.; Bouchard, R.A.; Finer, J.J. A soybean (Glycine max) polyubiquitin promoter gives strong constitutive expression in transgenic soybean. Plant Cell Rep 2009, 28, 837–849. [Google Scholar]
- Wei, H.; Albert, H.; Moore, P. Differential expression of sugarcane polyubiquitin genes and isolation of promoters from two highly-expressed members of the gene family. J. Plant Physiol 1999, 155, 513–519. [Google Scholar]
- Wei, H.; Wang, M.L.; Moore, P.H.; Albert, H.H. Comparative expression analysis of two sugarcane polyubiquitin promoters and flanking sequences in transgenic plants. J. Plant Physiol 2003, 160, 1241–1251. [Google Scholar]
- Garbarino, J.E.; Oosumi, T.; Belknap, W.R. Isolation of a polyubiquitin promoter and its expression in transgenic potato plants. Plant Physiol 1995, 109, 1371–1378. [Google Scholar]
- Rockhold, D.R.; Chang, S.; Taylor, N.; Allen, P.V.; McCue, K.F.; Belknap, W.R. Structure of two Solanum bulbocastanum polyubiquitin genes and expression of their promoters in transgenic potatoes. Am. J. Potato Res 2008, 85, 219–226. [Google Scholar]
- Foster, E.; Hattori, J.; Labbe, H.; Ouellet, T.; Fobert, P.R.; James, L.E.; Iyer, V.N.; Miki, B.L. A tobacco cryptic constitutive promoter, tCUP, revealed by T-DNA tagging. Plant Mol. Biol 1999, 41, 45–55. [Google Scholar]
- Malik, K.; Wu, K.; Li, X.Q.; Martin-Heller, T.; Hu, M.; Foster, E.; Tian, L.; Wang, C.; Ward, K.; Jordan, M.; et al. A constitutive gene expression system derived from the tCUP cryptic promoter elements. Theor. Appl. Genet 2002, 105, 505–514. [Google Scholar]
- Wu, K.; Malik, K.; Tian, L.; Hu, M.; Martin, T.; Foster, E.; Brown, D.; Miki, B. Enhancers and core promoter elements are essential for the activity of a cryptic gene activation sequence from tobacco, tCUP. Mol. Genet. Genomics 2001, 265, 763–770. [Google Scholar]
- McElroy, D.; Blowers, A.D.; Jenes, B.; Wu, R. Construction of expression vectors based on the rice actin 1 (Act1) 5′ region for use in monocot transformation. Mol. Gen. Genet 1991, 231, 150–160. [Google Scholar]
- McElroy, D.; Zhang, W.; Cao, J.; Wu, R. Isolation of an efficient actin promoter for use in rice transformation. Plant Cell 1990, 2, 163–171. [Google Scholar]
- Pater, B.S.; Mark, F.; Rueb, S.; Katagiri, F.; Chua, N.H.; Schilperoort, R.A.; Hensgens, L.A.M. The promoter of the rice gene GOS2 is active in various different monocot tissues and binds rice nuclear factor ASF-1. Plant J 1992, 2, 837–844. [Google Scholar]
- Jeon, J.S.; Lee, S.; Jung, K.H.; Jun, S.H.; Kim, C.; An, G. Tissue-preferential expression of a rice alpha-tubulin gene, OsTubA1, mediated by the first intron. Plant Physiol 2000, 123, 1005–1014. [Google Scholar]
- Jang, I.C.; Choi, W.B.; Lee, K.H.; Song, S.I.; Nahm, B.H.; Kim, J.K. High-level and ubiquitous expression of the rice cytochrome c gene OsCc1 and its promoter activity in transgenic plants provides a useful promoter for transgenesis of monocots. Plant Physiol 2002, 129, 1473–1481. [Google Scholar]
- Wang, J.; Oard, J.H. Rice ubiquitin promoters: Deletion analysis and potential usefulness in plant transformation systems. Plant Cell Rep 2003, 22, 129–134. [Google Scholar]
- He, C.; Lin, Z.; McElroy, D.; Wu, R. Identification of a rice actin2 gene regulatory region for high-level expression of transgenes in monocots. Plant Biotechnol. J 2009, 7, 227–239. [Google Scholar]
- Qin, J.Y.; Zhang, L.; Clift, K.L.; Hulur, I.; Xiang, A.P.; Ren, B.Z.; Lahn, B.T. Systematic comparison of constitutive promoters and the doxycycline-inducible promoter. PLoS One 2010, 5, e10611. [Google Scholar]
- Cheng, X.; Sardana, R.; Kaplan, H.; Altosaar, I. Agrobacterium-transformed rice plants expressing synthetic cryIA(b) and cryIA(c) genes are highly toxic to striped stem borer and yellow stem borer. Proc. Natl. Acad. Sci. USA 1998, 95, 2767–2772. [Google Scholar]
- Van der Salm, T.; Bosch, D.; Honee, G.; Feng, L.; Munsterman, E.; Bakker, P.; Stiekema, W.J.; Visser, B. Insect resistance of transgenic plants that express modified Bacillus thuringiensis cryIA(b) and cryIC genes: A resistance management strategy. Plant Mol. Biol 1994, 26, 51–59. [Google Scholar]
- Chen, L.H.; Zhang, B.; Xu, Z.Q. Salt tolerance conferred by overexpression of Arabidopsis vacuolar Na(+)/H(+) antiporter gene AtNHX1 in common buckwheat (Fagopyrum esculentum). Transgenic Res 2008, 17, 121–132. [Google Scholar]
- Xu, D.; Duan, X.; Wang, B.; Hong, B.; Ho, T.; Wu, R. Expression of a late embryogenesis abundant protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol 1996, 110, 249–257. [Google Scholar]
- Botterman, J.; Leemans, J. Discovery, transfer to crops, expression and biological significance of a bialaphos resistance gene. Monogr.-Br. Crop Prot. Counc 1989, 42, 63–68. [Google Scholar]
- Korban, S.S. Targeting and expression of antigenic proteins in transgenic plants for production of edible oral vaccines. In Vitro Cell Dev. Biol. -Plant 2002, 38, 231–236. [Google Scholar]
- Gittins, J.R.; Pellny, T.K.; Hiles, E.R.; Rosa, C.; Biricolti, S.; James, D.J. Transgene expression driven by heterologous ribulose-1,5-bisphosphate carboxylase/oxygenase small-subunit gene promoters in the vegetative tissues of apple (Malus pumila Mill.). Planta 2000, 210, 232–240. [Google Scholar]
- Robinson, D.J. Environmental risk assessment of releases of transgenic plants containing virus-derived inserts. Transgenic Res 1996, 5, 359–362. [Google Scholar]
- Kohli, A.; Griffiths, S.; Palacios, N.; Twyman, R.M.; Vain, P.; Laurie, D.A.; Christou, P. Molecular characterization of transforming plasmid rearrangements in transgenic rice reveals a recombination hotspot in the CaMV 35S promoter and confirms the predominance of microhomology mediated recombination. Plant J 1999, 17, 591–601. [Google Scholar]
- Myhre, M.R.; Fenton, K.A.; Eggert, J.; Nielsen, K.M.; Traavik, T. The 35S CaMV plant virus promoter is active in human enterocyte-like cells. Eur. Food Res. Technol 2006, 222, 185–193. [Google Scholar]
- Gibbs, M.J.; Weiller, G.F. Evidence that a plant virus switched hosts to infect a vertebrate and then recombined with a vertebrate-infecting virus. Proc. Natl. Acad. Sci. USA 1999, 96, 8022. [Google Scholar]
- Matzke, M.A.; Matzke, A. How and why do plants inactivate homologous (trans)genes? Plant Physiol 1995, 107, 679–685. [Google Scholar]
- Stam, M.; Mol, J.N.M.; Kooter, J.M. The Silence of genes in transgenic plants. Ann. Bot.-Lond 1997, 79, 3–12. [Google Scholar]
- Al-Kaff, N.S.; Kreike, M.M.; Covey, S.N.; Pitcher, R.; Page, A.M.; Dale, P.J. Plants rendered herbicide-susceptible by cauliflower mosaic virus-elicited suppression of a 35S promoter-regulated transgene. Nat. Biotechnol 2000, 18, 995–999. [Google Scholar]
- Dong, Y.; von Arnim, A.G. Novel plant activation-tagging vectors designed to minimize 35S enhancer-mediated gene silencing. Plant Mol. Biol. Rep 2003, 21, 349–358. [Google Scholar]
- Jakowitsch, J.; Papp, I.; Moscone, E.A.; van der Winden, J.; Matzke, M.; Matzke, A.J. Molecular and cytogenetic characterization of a transgene locus that induces silencing and methylation of homologous promoters in trans. Plant J 1999, 17, 131–140. [Google Scholar]
- McCabe, M.S.; Schepers, F.; van der Arend, A.; Mohapatra, U.; de Laat, A.M.; Power, J.B.; Davey, M.R. Increased stable inheritance of herbicide resistance in transgenic lettuce carrying a petE promoter-bar gene compared with a CaMV 35S-bar gene. TAG Theor. Appl. Genet 1999, 99, 587–592. [Google Scholar]
- Mette, M.F.; Aufsatz, W.; van der Winden, J.; Matzke, M.A.; Matzke, A.J. Transcriptional silencing and promoter methylation triggered by double-stranded RNA. EMBO J 2000, 19, 5194–5201. [Google Scholar]
- Park, Y.D.; Papp, I.; Moscone, E.A.; Iglesias, V.A.; Vaucheret, H.; Matzke, A.J.; Matzke, M.A. Gene silencing mediated by promoter homology occurs at the level of transcription and results in meiotically heritable alterations in methylation and gene activity. Plant J 1996, 9, 183–194. [Google Scholar]
- Al-Kaff, N.S.; Covey, S.N.; Kreike, M.M.; Page, A.M.; Pinder, R.; Dale, P.J. Transcriptional and posttranscriptional plant gene silencing in response to a pathogen. Science 1998, 279, 2113–2115. [Google Scholar]
- Vaucheret, H.; Beclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Mourrain, P.; Palauqui, J.C.; Vernhettes, S. Transgene-induced gene silencing in plants. Plant J 1998, 16, 651–659. [Google Scholar]
- Wang, L.; Li, L.; Xu, L.; Zhou, J.; Zhuang, H.; Gong, X.; Wang, M.; Sun, S.S.M.; Zhuge, Q. Isolation and functional analysis of the Poplar RbcS gene promoter. Plant Mol. Biol. Rep 2013, 31, 120–127. [Google Scholar]
- Yang, Y.; Yang, G.; Liu, S.; Guo, X.; Zheng, C. Isolation and functional analysis of a strong specific promoter in photosynthetic tissues. Sci. China Ser. C: Life Sci 2003, 46, 651–660. [Google Scholar]
- Noh, S.A.; Lee, H.S.; Huh, G.H.; Oh, M.J.; Paek, K.H.; Shin, J.S.; Bae, J.M. A sweetpotato SRD1 promoter confers strong root-, taproot-, and tuber-specific expression in Arabidopsis, carrot, and potato. Transgenic Res 2012, 21, 265–278. [Google Scholar]
- Chawla, R.; Shakya, R.; Rommens, C.M. Tuber-specific silencing of asparagine synthetase-1 reduces the acrylamide-forming potential of potatoes grown in the field without affecting tuber shape and yield. Plant Biotechnol. J 2012, 10, 913–924. [Google Scholar]
- Arango, J.; Salazar, B.; Welsch, R.; Sarmiento, F.; Beyer, P.; Al-Babili, S. Putative storage root specific promoters from cassava and yam: Cloning and evaluation in transgenic carrots as a model system. Plant Cell Rep 2010, 29, 651–659. [Google Scholar]
- Xu, L.; Ye, R.; Zheng, Y.; Wang, Z.; Zhou, P.; Lin, Y.; Li, D. Isolation of the endosperm-specific LPAAT gene promoter from coconut (Cocos nucifera L.) and its functional analysis in transgenic rice plants. Plant Cell Rep 2010, 29, 1061–1068. [Google Scholar]
- Zavallo, D.; Lopez Bilbao, M.; Hopp, H.E.; Heinz, R. Isolation and functional characterization of two novel seed-specific promoters from sunflower (Helianthus annuus L.). Plant Cell Rep 2010, 29, 239–248. [Google Scholar]
- Kato, H.; Xie, G.; Sato, Y.; Imai, R. Isolation of anther-specific gene promoters suitable for transgene expression in rice. Plant Mol. Biol. Rep 2010, 28, 381–387. [Google Scholar]
- Huang, Z.; Gan, Z.; He, Y.; Li, Y.; Liu, X.; Mu, H. Functional analysis of a rice late pollen-abundant UDP-glucose pyrophosphorylase (OsUgp2) promoter. Mol. Biol. Rep 2011, 38, 4291–4302. [Google Scholar]
- Yi, N.; Kim, Y.S.; Jeong, M.H.; Oh, S.J.; Jeong, J.S.; Park, S.H.; Jung, H.; Choi, Y.D.; Kim, J.K. Functional analysis of six drought-inducible promoters in transgenic rice plants throughout all stages of plant growth. Planta 2010, 232, 743–754. [Google Scholar]
- Freeman, J.; Sparks, C.A.; West, J.; Shewry, P.R.; Jones, H.D. Temporal and spatial control of transgene expression using a heat-inducible promoter in transgenic wheat. Plant Biotechnol. J 2011, 9, 788–796. [Google Scholar] [Green Version]
- Tittarelli, A.; Santiago, M.; Morales, A.; Meisel, L.A.; Silva, H. Isolation and functional characterization of cold-regulated promoters, by digitally identifying peach fruit cold-induced genes from a large EST dataset. BMC Plant Biol 2009, 9, 121. [Google Scholar]
- Su, C.F.; Wang, Y.C.; Hsieh, T.H.; Lu, C.A.; Tseng, T.H.; Yu, S.M. A novel MYBS3-dependent pathway confers cold tolerance in rice. Plant Physiol 2010, 153, 145–158. [Google Scholar]
- Kovalchuk, N.; Li, M.; Wittek, F.; Reid, N.; Singh, R.; Shirley, N.; Ismagul, A.; Eliby, S.; Johnson, A.; Milligan, A.S. Defensin promoters as potential tools for engineering disease resistance in cereal grains. Plant Biotechnol. J 2010, 8, 47–64. [Google Scholar]
- Feuillet, C.; Lauvergeat, V.; Deswarte, C.; Pilate, G.; Boudet, A.; Grima-Pettenati, J. Tissue- and cell-specific expression of a cinnamyl alcohol dehydrogenase promoter in transgenic poplar plants. Plant Mol. Biol 1995, 27, 651–667. [Google Scholar]
- Hollick, J.B.; Gordon, M.P. Transgenic analysis of a hybrid poplar wound-inducible promoter reveals developmental patterns of expression similar to that of storage protein genes. Plant Physiol 1995, 109, 73–85. [Google Scholar]
- Wei, H.; Meilan, R.; Brunner, A.M.; Skinner, J.S.; Ma, C.; Gandhi, H.T.; Strauss, S.H. Field trial detects incomplete barstar attenuation of vegetative cytotoxicity in Populus trees containing a poplar LEAFY promoter::barnase sterility transgene. Mol. Breed 2007, 19, 69–85. [Google Scholar]
- Yevtushenko, D.P.; Sidorov, V.A.; Romero, R.; Kay, W.W.; Misra, S. Wound-inducible promoter from poplar is responsive to fungal infection in transgenic potato. Plant Sci 2004, 167, 715–724. [Google Scholar]
- Zheng, H.; Lei, Y.; Zhang, Z.; Lin, S.; Zhang, Q.; Liu, W.; Du, J.; An, X.; Zhao, X. Analysis of promoter activity of PtDrl02 gene in white poplars. J. Plant Biochem. Biot 2012, 21, 88–97. [Google Scholar]
- Zheng, H.; Lin, S.; Zhang, Q.; Lei, Y.; Hou, L.; Zhang, Z. Functional identification and regulation of the PtDrl02 gene promoter from triploid white poplar. Plant Cell Rep 2010, 29, 449–460. [Google Scholar]
- Zhong, R.; Lee, C.; Ye, Z.H. Functional characterization of poplar wood-associated NAC domain transcription factors. Plant Physiol 2010, 152, 1044–1055. [Google Scholar]
- Park, S.H.; Yi, N.; Kim, Y.S.; Jeong, M.H.; Bang, S.W.; Choi, Y.D.; Kim, J.K. Analysis of five novel putative constitutive gene promoters in transgenic rice plants. J. Exp. Bot 2010, 61, 2459–2467. [Google Scholar]
- Shahmuradov, I.A.; Solovyev, V.V.; Gammerman, A.J. Plant promoter prediction with confidence estimation. Nucleic Acids Res 2005, 33, 1069–1076. [Google Scholar]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res 1999, 27, 297–300. [Google Scholar]
- Prestridge, D.S. SIGNAL SCAN: A computer program that scans DNA sequences for eukaryotic transcriptional elements. Comput. Appl. Biosci 1991, 7, 203–206. [Google Scholar]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 2002, 30, 325–327. [Google Scholar]
- Donald, R.G.; Cashmore, A.R. Mutation of either G box or I box sequences profoundly affects expression from the Arabidopsis rbcS-1A promoter. EMBO J 1990, 9, 1717–1726. [Google Scholar]
- Orozco, B.M.; Ogren, W.L. Localization of light-inducible and tissue-specific regions of the spinach ribulose bisphosphate carboxylase/oxygenase (rubisco) activase promoter in transgenic tobacco plants. Plant Mol. Biol 1993, 23, 1129–1138. [Google Scholar]
- Ozkaynak, E.; Finley, D.; Solomon, M.J.; Varshavsky, A. The yeast ubiquitin genes: A family of natural gene fusions. EMBO J 1987, 6, 1429–1439. [Google Scholar]
- Gausing, K.; Jensen, C.B. Two ubiquitin-long-tail fusion genes arranged as closely spaced direct repeats in barley. Gene 1990, 94, 165–171. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 1998, 16, 735–743. [Google Scholar]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc 2006, 1, 641–646. [Google Scholar]
- Jia, Z.; Gou, J.; Sun, Y.; Yuan, L.; Tang, Q.; Yang, X.; Pei, Y.; Luo, K. Enhanced resistance to fungal pathogens in transgenic Populus tomentosa Carr. by overexpression of an nsLTP-like antimicrobial protein gene from motherwort (Leonurus japonicus). Tree Physiol 2010, 30, 1599–1605. [Google Scholar]
- Hofgen, R.; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res 1988, 16, 9877. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep 1993, 11, 113–116. [Google Scholar]
- Jefferson, R.A.; Bevan, M.; Kavanagh, T. The use of the Escherichia coli beta-glucuronidase as a gene fusion marker for studies of gene expression in higher plants. Biochem. Soc. Trans 1987, 15, 17–18. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]






© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Z.; Wang, J.; Ye, M.-X.; Li, H.; Ji, L.-X.; Li, Y.; Cui, D.-Q.; Liu, J.-M.; An, X.-M. A Novel Moderate Constitutive Promoter Derived from Poplar (Populus tomentosa Carrière). Int. J. Mol. Sci. 2013, 14, 6187-6204. https://doi.org/10.3390/ijms14036187
Chen Z, Wang J, Ye M-X, Li H, Ji L-X, Li Y, Cui D-Q, Liu J-M, An X-M. A Novel Moderate Constitutive Promoter Derived from Poplar (Populus tomentosa Carrière). International Journal of Molecular Sciences. 2013; 14(3):6187-6204. https://doi.org/10.3390/ijms14036187
Chicago/Turabian StyleChen, Zhong, Jia Wang, Mei-Xia Ye, Hao Li, Le-Xiang Ji, Ying Li, Dong-Qing Cui, Jun-Mei Liu, and Xin-Min An. 2013. "A Novel Moderate Constitutive Promoter Derived from Poplar (Populus tomentosa Carrière)" International Journal of Molecular Sciences 14, no. 3: 6187-6204. https://doi.org/10.3390/ijms14036187
APA StyleChen, Z., Wang, J., Ye, M. -X., Li, H., Ji, L. -X., Li, Y., Cui, D. -Q., Liu, J. -M., & An, X. -M. (2013). A Novel Moderate Constitutive Promoter Derived from Poplar (Populus tomentosa Carrière). International Journal of Molecular Sciences, 14(3), 6187-6204. https://doi.org/10.3390/ijms14036187