Ischemic Preconditioning Protects against Spinal Cord Ischemia-Reperfusion Injury in Rabbits by Attenuating Blood Spinal Cord Barrier Disruption

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
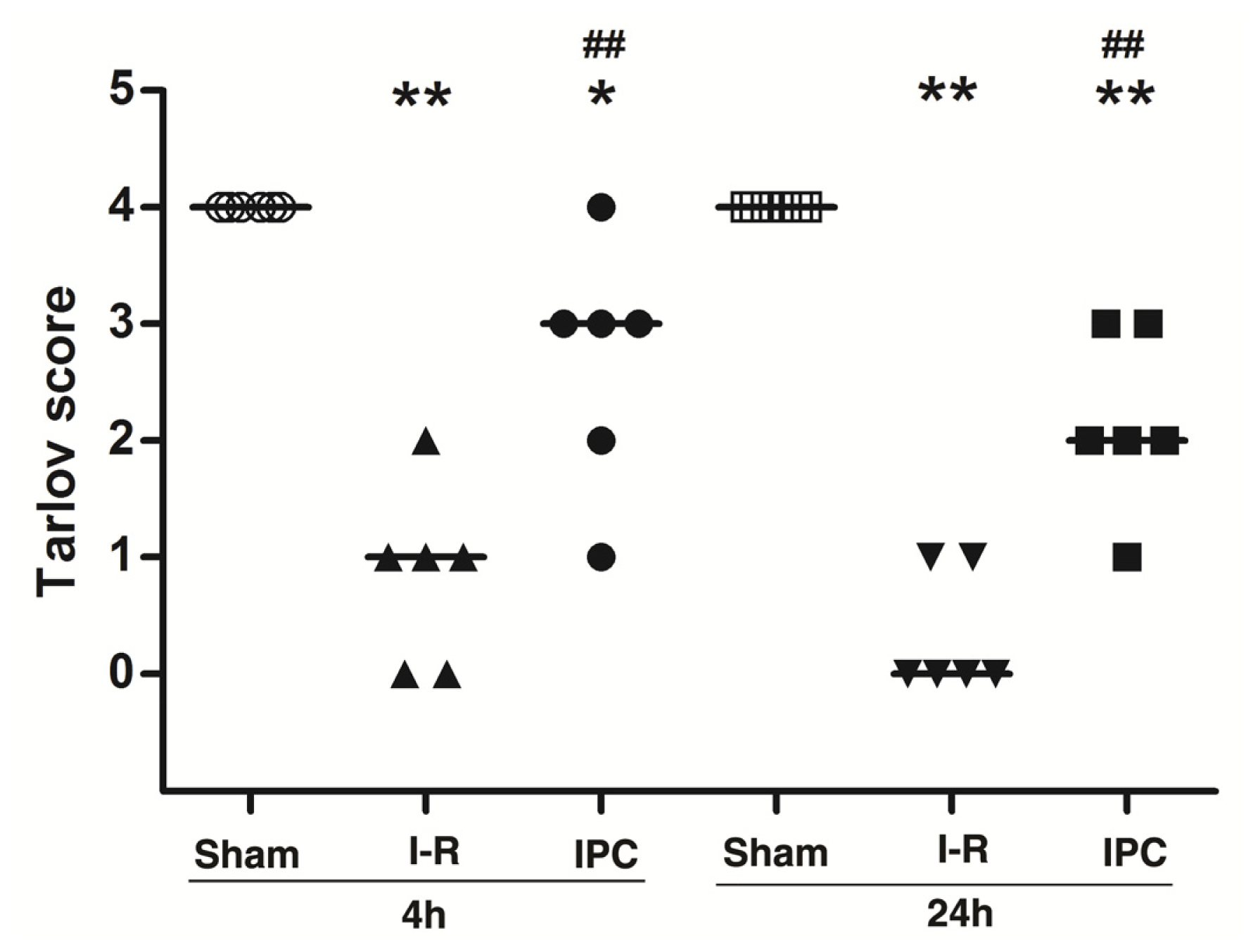
2.1. IPC Improves Neurologic Scores after Spinal Cord I-R Injury
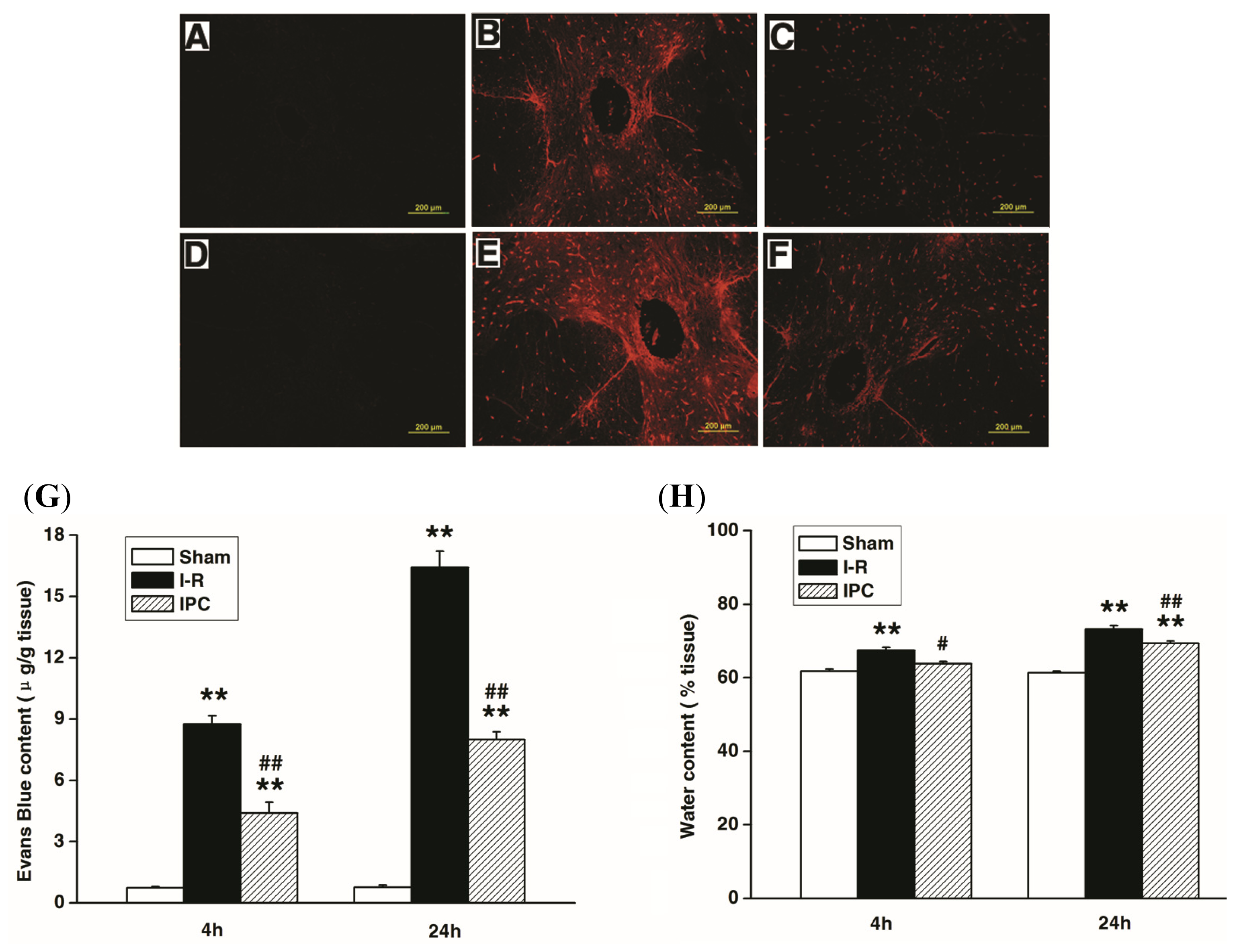
2.2. IPC Inhibits Blood Spinal Cord Barrier Breakdown after Spinal Cord I-R Injury
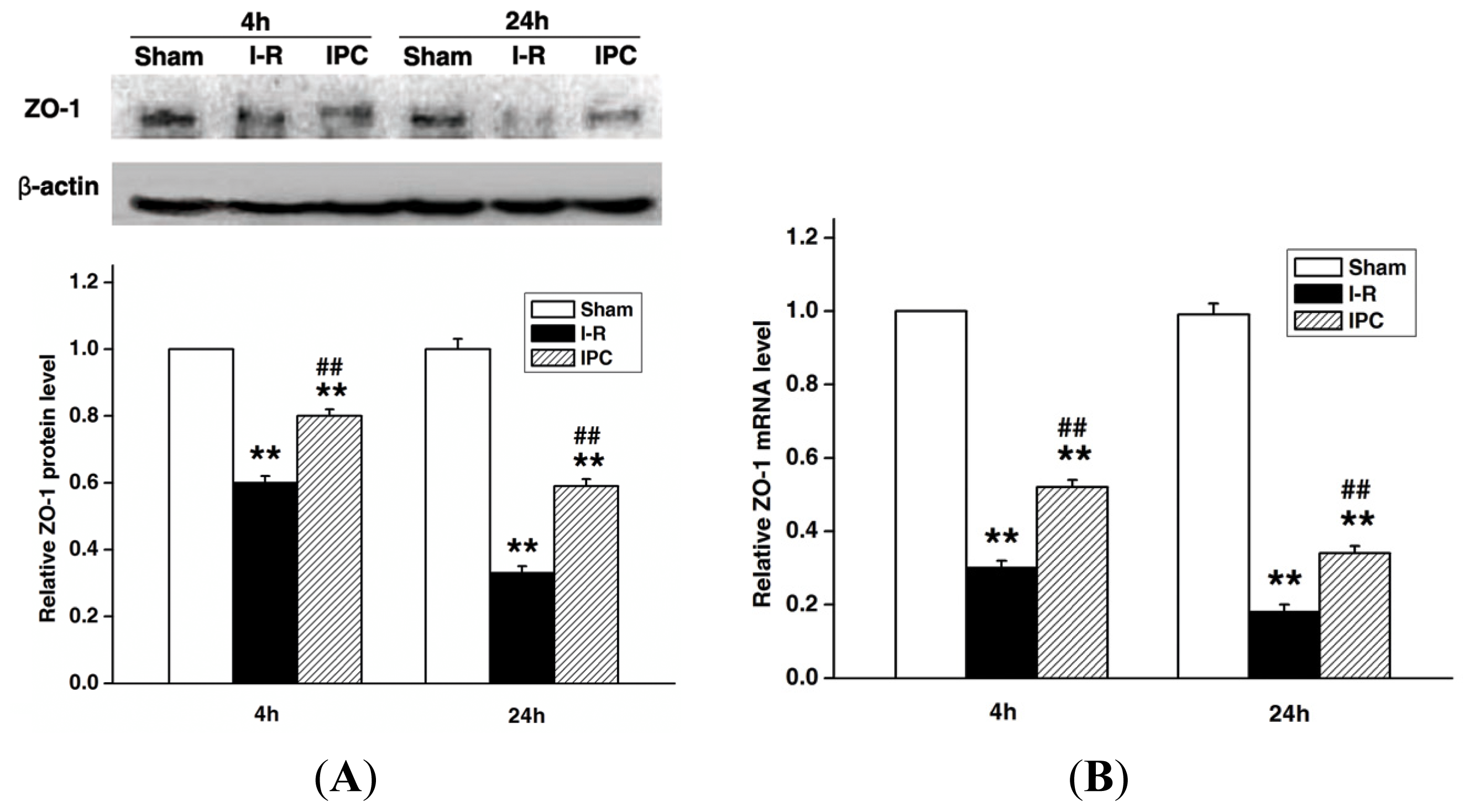
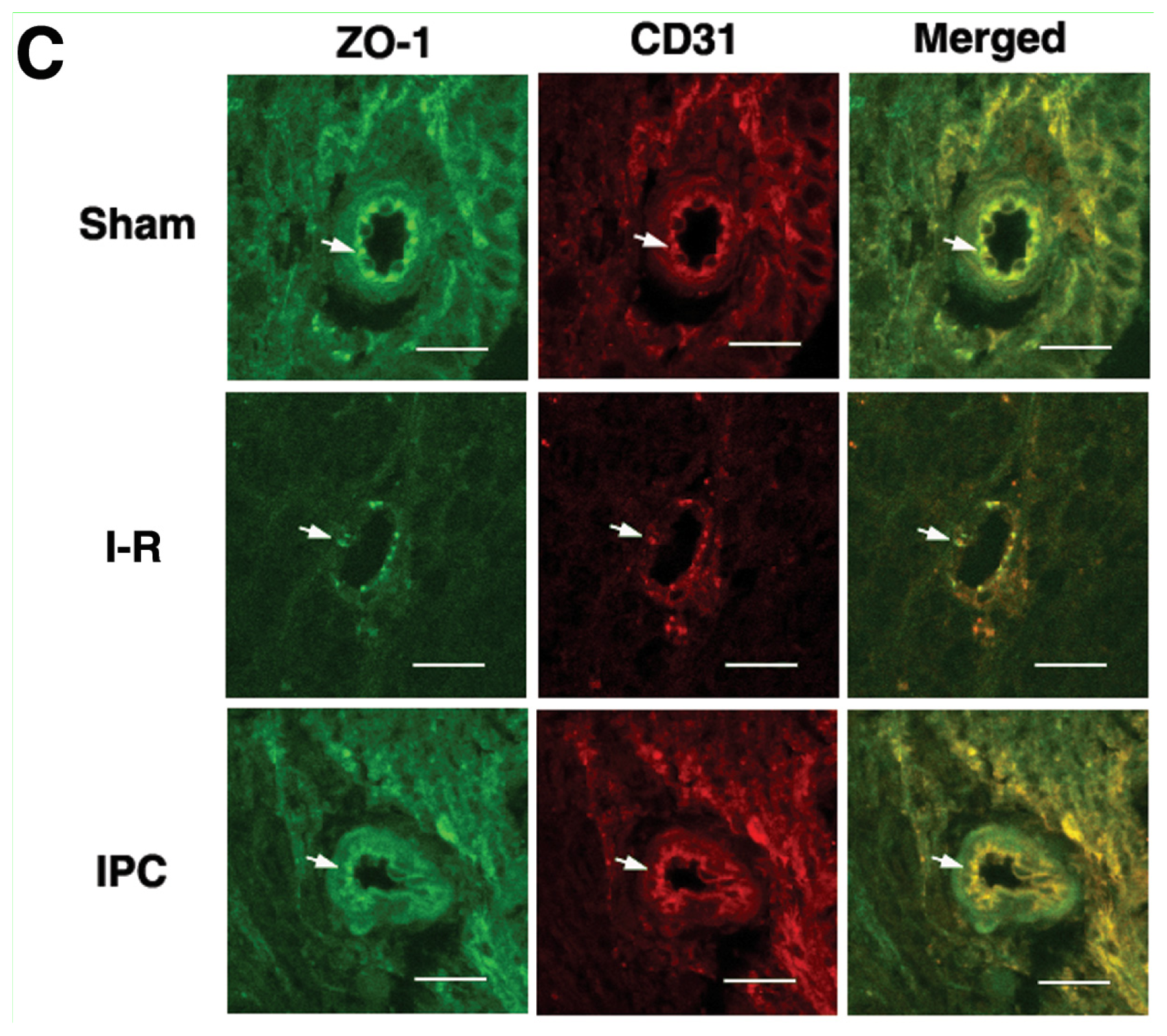
2.3. IPC Preserves Tight Junction Protein ZO-1 after Spinal Cord I-R Injury
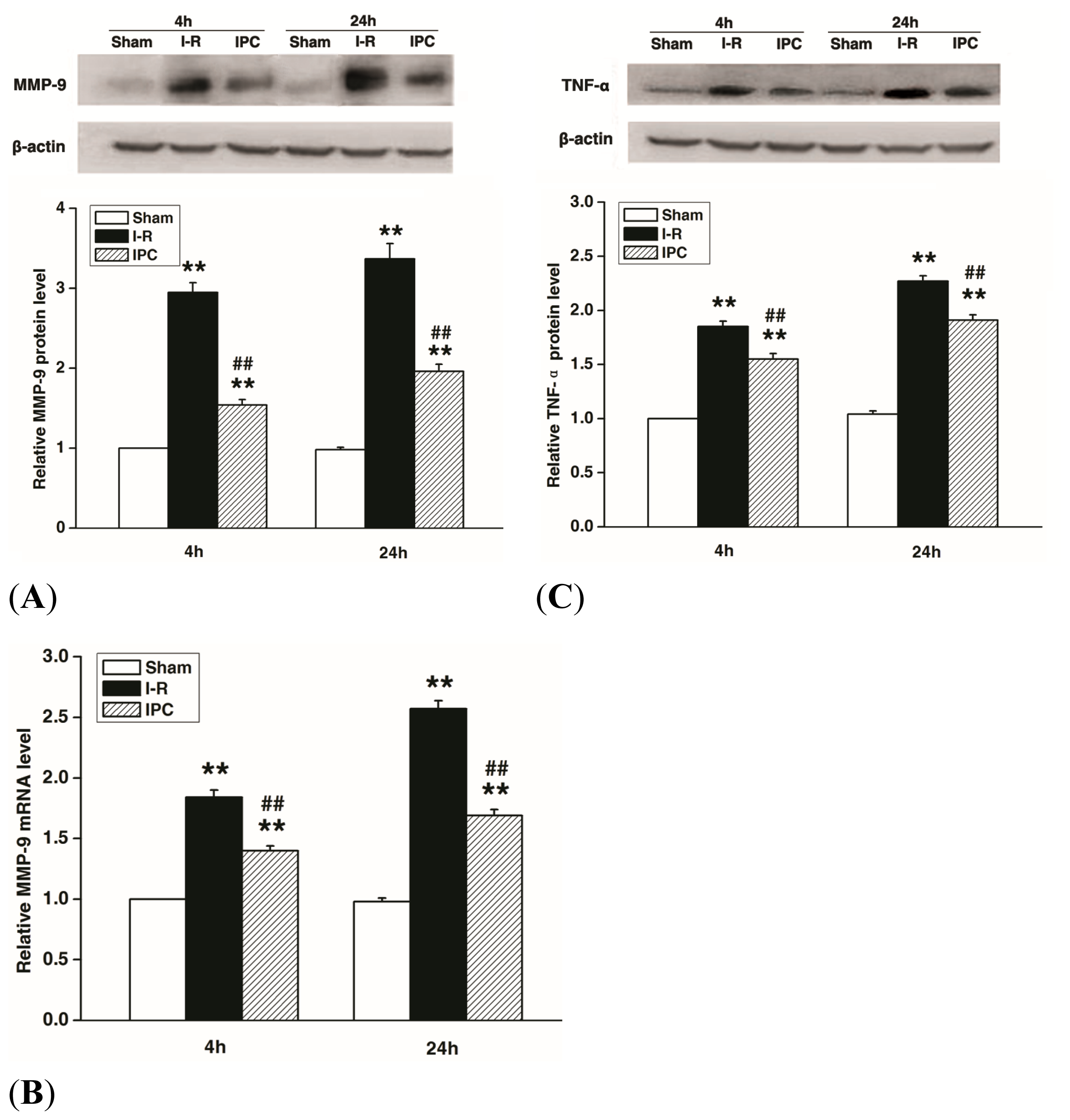
2.4. IPC Inhibits MMP-9 and TNF-α Expression after Spinal Cord I-R Injury
3. Experimental Section
3.1. Animal Care and Experimental Protocol
3.2. Surgical Procedure
3.3. Neurologic Assessment
3.4. Measurement of Blood Spinal Cord Barrier Permeability
3.5. Measurement of Spinal Cord Edema
3.6. Immunofluorescence of Tight Junction Protein ZO-1
3.7. Western Blots
3.8. Quantitative Real-Time Polymerase Chain Reaction Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Kuniyoshi, Y.; Koja, K.; Miyagi, K.; Shimoji, M.; Uezu, T.; Arakaki, K.; Yamashiro, S.; Mabuni, K.; Senaha, S.; Nakasone, Y. Prevention of postoperative paraplegia during thoracoabdominal aortic surgery. Ann. Thorac. Surg 2003, 76, 1477–1484. [Google Scholar]
- Coselli, J.S.; LeMaire, S.A.; Miller, C.C.; Schmittling, Z.C.; Köksoy, C.; Pagan, J.; Curling, P.E. Mortality and paraplegia after thoracoabdominal aortic aneurysm repair: A risk factor analysis. Ann. Thorac. Surg 2000, 69, 409–414. [Google Scholar]
- Bischoff, M.S.; Di, L.G.; Griepp, E.B.; Griepp, R.B. Spinal cord preservation in thoracoabdominal aneurysm repair. Perspect. Vasc. Surg. Endovasc. Ther 2011, 23, 214–222. [Google Scholar]
- Lee, J.Y.; Kim, H.S.; Choi, H.Y.; Oh, T.H.; Yune, T.Y. Fluoxetine inhibits matrix metalloprotease activation and prevents disruption of blood-spinal cord barrier after spinal cord injury. Brain 2012, 135, 2375–2389. [Google Scholar]
- Sharma, H.S. Pathophysiology of blood-spinal cord barrier in traumatic injury and repair. Curr. Pharm. Des 2005, 11, 1353–1389. [Google Scholar]
- Liang, C.L.; Lu, K.; Liliang, P.C.; Chen, T.B.; Chan, S.H.; Chen, H.J. Ischemic preconditioning ameliorates spinal cord ischemia-reperfusion injury by triggering autoregulation. J. Vasc. Surg 2012, 55, 1116–1123. [Google Scholar]
- Kyrou, I.E.; Papakostas, J.C.; Ioachim, E.; Koulouras, V.; Arnaoutoglou, E.; Angelidis, C.; Matsagkas, M.I. Early ischaemic preconditioning of spinal cord enhanced the binding profile of heat shock protein 70 with neurofilaments and promoted its nuclear translocation after thoraco-abdominal aortic occlusion in pigs. Eur. J. Vasc. Endovasc. Surg 2012, 43, 408–414. [Google Scholar]
- Yang, C.; Ren, Y.; Liu, F.; Cai, W.; Zhang, N.; Nagel, D.J.; Yin, G. Ischemic preconditioning suppresses apoptosis of rabbit spinal neurocytes by inhibiting ASK1–14–3-3 dissociation. Neurosci. Lett 2008, 441, 267–271. [Google Scholar]
- Masada, T.; Hua, Y.; Xi, G.; Ennis, S.R.; Keep, R.F. Attenuation of ischemic brain edema and cerebrovascular injury after ischemic preconditioning in the rat. J. Cereb. Blood. Flow. Metab 2001, 21, 22–33. [Google Scholar]
- Jiao, H.; Wang, Z.; Liu, Y.; Wang, P.; Xue, Y. Specific role of tight junction proteins claudin-5, occludin, and ZO-1 of the blood-brain barrier in a focal cerebral ischemic insult. J. Mol. Neurosci 2011, 44, 130–139. [Google Scholar]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood. Flow. Metab 2007, 27, 697–709. [Google Scholar]
- Noble, L.J.; Donovan, F.; Igarashi, T.; Goussev, S.; Werb, Z. Matrix metalloproteinases limit functional recovery after spinal cord injury by modulation of early vascular events. J. Neurosci 2002, 22, 7526–7535. [Google Scholar]
- Mayhan, W.G. Cellular mechanisms by which tumor necrosis factor-alpha produces disruption of the blood-brain barrier. Brain. Res 2002, 927, 144–152. [Google Scholar]
- Saito, T.; Tsuchida, M.; Umehara, S.; Kohno, T.; Yamamoto, H.; Hayashi, J. Reduction of spinal cord ischemia/reperfusion injury with simvastatin in rats. Anesth. Analg 2011, 113, 565–571. [Google Scholar]
- Matsuda, H.; Ogino, H.; Fukuda, T.; Iritani, O.; Sato, S.; Iba, Y.; Tanaka, H.; Sasaki, H.; Minatoya, K.; Kobayashi, J.; et al. Multidisciplinary approach to prevent spinal cord ischemia after thoracic endovascular aneurysm repair for distal descending aorta. Ann. Thorac. Surg 2010, 90, 561–565. [Google Scholar]
- Shi, E.; Jiang, X.; Wang, L.; Akuzawa, S.; Nakajima, Y.; Kazui, T. Intrathecal injection of hepatocyte growth factor gene-modified marrow stromal cells attenuates neurologic injury induced by transient spinal cord ischemia in rabbits. Anesthesiology 2010, 113, 1109–1117. [Google Scholar]
- Zhang, Q.; Huang, C.; Meng, B.; Tang, T.; Shi, Q.; Yang, H. Acute effect of ghrelin on ischemia/reperfusion injury in the rat spinal cord. Int. J. Mol. Sci 2012, 13, 9864–9876. [Google Scholar]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar]
- Nico, B.; Ribatti, D. Morphofunctional aspects of the blood-brain barrier. Curr. Drug. Metab 2012, 13, 50–60. [Google Scholar]
- Ramos-Fernandez, M.; Bellolio, M.F.; Stead, L.G. Matrix metalloproteinase-9 as a marker for acute ischemic stroke: a systematic review. J. Stroke. Cerebrovasc. Dis 2011, 20, 47–54. [Google Scholar]
- Liu, K.; Sun, T.; Wang, P.; Liu, Y.H.; Zhang, L.W.; Xue, Y.X. Effects of erythropoietin on blood-brain barrier tight junctions in ischemia-reperfusion rats. J. Mol. Neurosci 2012, 49, 369–379. [Google Scholar]
- Takata, F.; Dohgu, S.; Matsumoto, J.; Takahashi, H.; Machida, T.; Wakigawa, T.; Harada, E.; Miyaji, H.; Koga, M.; Nishioku, T.; et al. Brain pericytes among cells constituting the blood-brain barrier are highly sensitive to tumor necrosis factor-α, releasing matrix metalloproteinase-9 and migrating in vitro. J. Neuroinflammation. 2011, 8. [Google Scholar] [CrossRef]
- Herlambang, B.; Orihashi, K.; Mizukami, T.; Takahashi, S.; Uchida, N.; Hiyama, E.; Sueda, T. New method for absolute spinal cord ischemia protection in rabbits. J. Vasc. Surg 2011, 54, 1109–1116. [Google Scholar]
- Shi, E.; Kazui, T.; Jiang, X.; Washiyama, N.; Yamashita, K.; Terada, H.; Bashar, A.H. Therapeutic benefit of intrathecal injection of marrow stromal cells on ischemia-injured spinal cord. Ann. Thorac. Surg 2007, 83, 1484–1490. [Google Scholar]
- Kakimoto, M.; Kawaguchi, M.; Sakamoto, T.; Inoue, S.; Furuya, H.; Nakamura, M.; Konishi, N. Evaluation of rapid ischemic preconditioning in a rabbit model of spinal cord ischemia. Anesthesiology 2003, 99, 1112–1117. [Google Scholar]
- Jacobs, T.P.; Kempski, O.; McKinley, D.; Dutka, A.J.; Hallenbeck, J.M.; Feuerstein, G. Blood flow and vascular permeability during motor dysfunction in a rabbit model of spinal cord ischemia. Stroke 1992, 23, 367–373. [Google Scholar]
- Yu, Q.J.; Wang, Y.L.; Zhou, Q.S.; Huang, H.B.; Tian, S.F.; Duan, D.M. Effect of repetitive ischemic preconditioning on spinal cord ischemia in a rabbit model. Life Sci 2006, 79, 1479–1483. [Google Scholar]
- Drummond, J.C.; Moore, S.S. The influence of dextrose administration on neurologic outcome after temporary spinal cord ischemia in the rabbit. Anesthesiology 1989, 70, 64–70. [Google Scholar]
- Fukatsu, K.; Ueno, C.; Maeshima, Y.; Mochizuki, H.; Saitoh, D. Detrimental effects of early nutrition administration after severe gut ischemia-reperfusion. J. Surg. Res 2008, 149, 31–38. [Google Scholar]
- Yan, M.; Dai, H.; Ding, T.; Dai, A.; Zhang, F.; Yu, L.; Chen, G.; Chen, Z. Effects of dexmedetomidine on the release of glial cell line-derived neurotrophic factor from rat astrocyte cells. Neurochem. Int 2011, 58, 549–557. [Google Scholar]
- Fan, Y.Y.; Hu, W.W.; Dai, H.B.; Zhang, J.X.; Zhang, L.Y.; He, P.; Shen, Y.; Ohtsu, H.; Wei, E.Q.; Chen, Z. Activation of the central histaminergic system is involved in hypoxia-induced stroke tolerance in adult mice. J. Cereb. Blood. Flow. Metab 2011, 31, 305–314. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar]





© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fang, B.; Li, X.-M.; Sun, X.-J.; Bao, N.-R.; Ren, X.-Y.; Lv, H.-W.; Ma, H. Ischemic Preconditioning Protects against Spinal Cord Ischemia-Reperfusion Injury in Rabbits by Attenuating Blood Spinal Cord Barrier Disruption. Int. J. Mol. Sci. 2013, 14, 10343-10354. https://doi.org/10.3390/ijms140510343
Fang B, Li X-M, Sun X-J, Bao N-R, Ren X-Y, Lv H-W, Ma H. Ischemic Preconditioning Protects against Spinal Cord Ischemia-Reperfusion Injury in Rabbits by Attenuating Blood Spinal Cord Barrier Disruption. International Journal of Molecular Sciences. 2013; 14(5):10343-10354. https://doi.org/10.3390/ijms140510343
Chicago/Turabian StyleFang, Bo, Xiao-Man Li, Xi-Jia Sun, Na-Ren Bao, Xiao-Yan Ren, Huang-Wei Lv, and Hong Ma. 2013. "Ischemic Preconditioning Protects against Spinal Cord Ischemia-Reperfusion Injury in Rabbits by Attenuating Blood Spinal Cord Barrier Disruption" International Journal of Molecular Sciences 14, no. 5: 10343-10354. https://doi.org/10.3390/ijms140510343
APA StyleFang, B., Li, X. -M., Sun, X. -J., Bao, N. -R., Ren, X. -Y., Lv, H. -W., & Ma, H. (2013). Ischemic Preconditioning Protects against Spinal Cord Ischemia-Reperfusion Injury in Rabbits by Attenuating Blood Spinal Cord Barrier Disruption. International Journal of Molecular Sciences, 14(5), 10343-10354. https://doi.org/10.3390/ijms140510343