Drought Stress Acclimation Imparts Tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
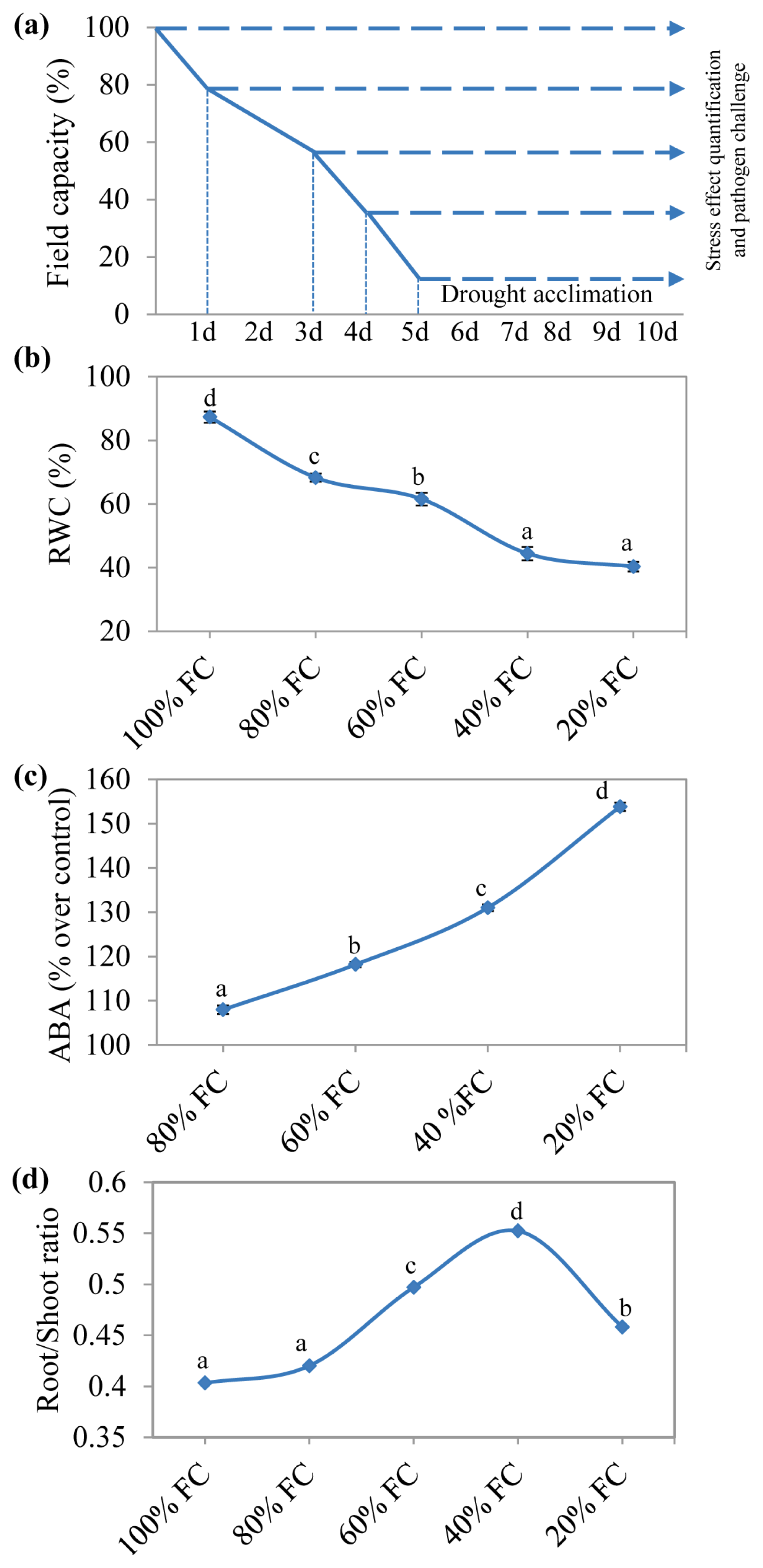
2.1. Drought Acclimated N. benthamiana Plants Showed Increased ABA and Root-Shoot Ratio
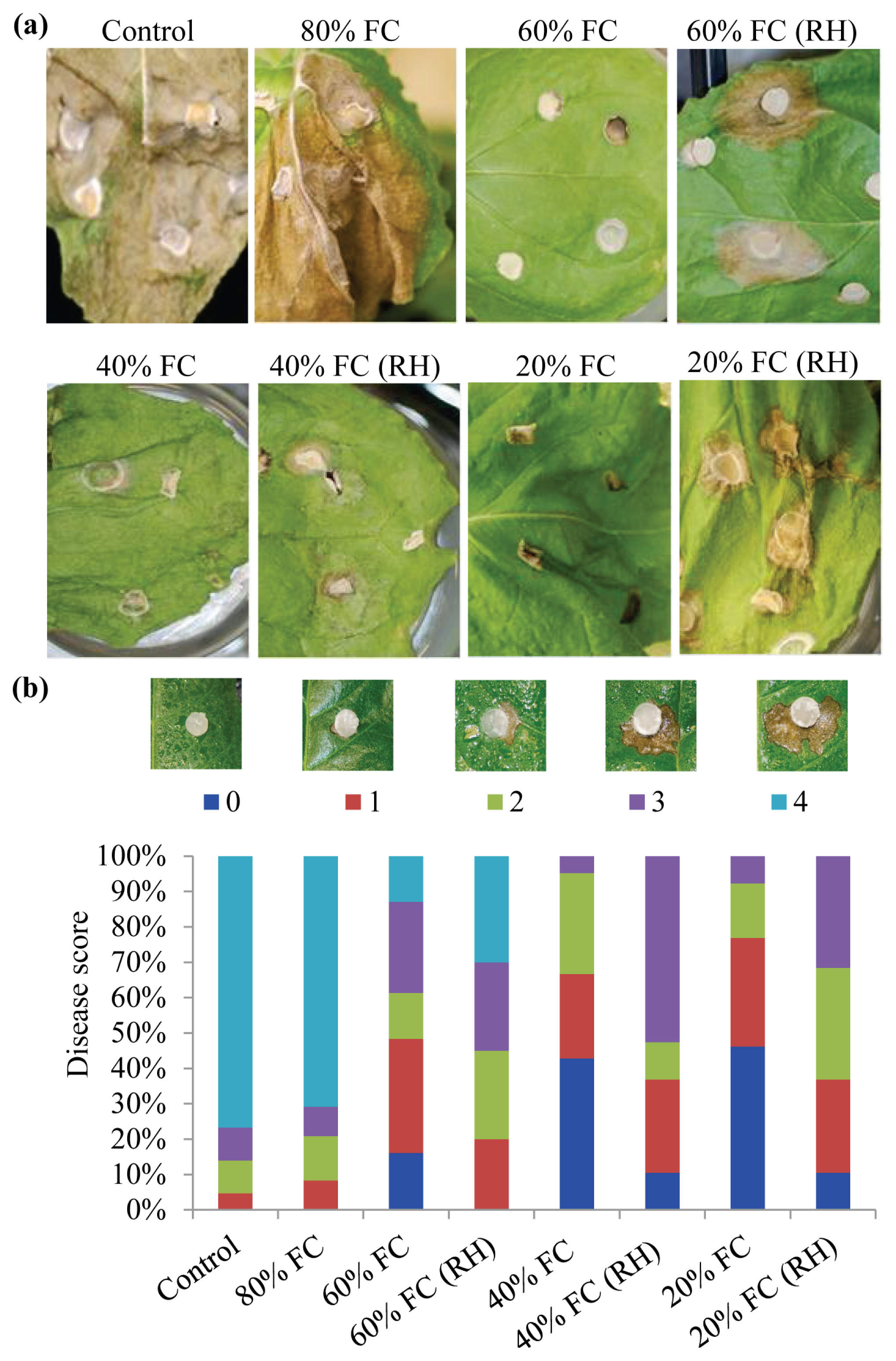
2.2. Drought Acclimation Reduced Sclerotinia sclerotiorum-Induced Cell Death in N. benthamiana
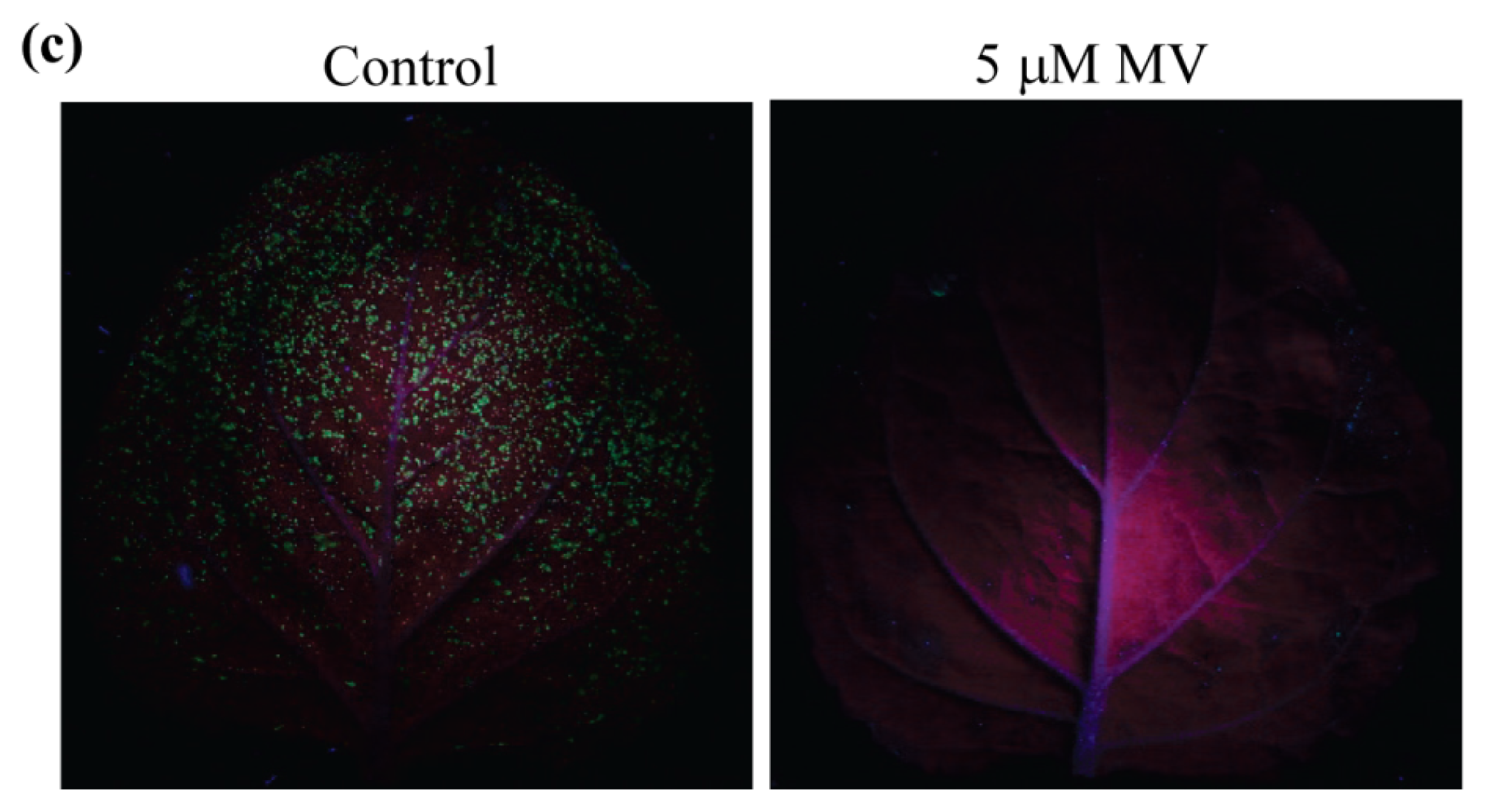
2.3. Acclimation to Moderate Drought Stress Enhanced Tolerance to Pseudomonas syringae pv. tabaci in N. benthamiana
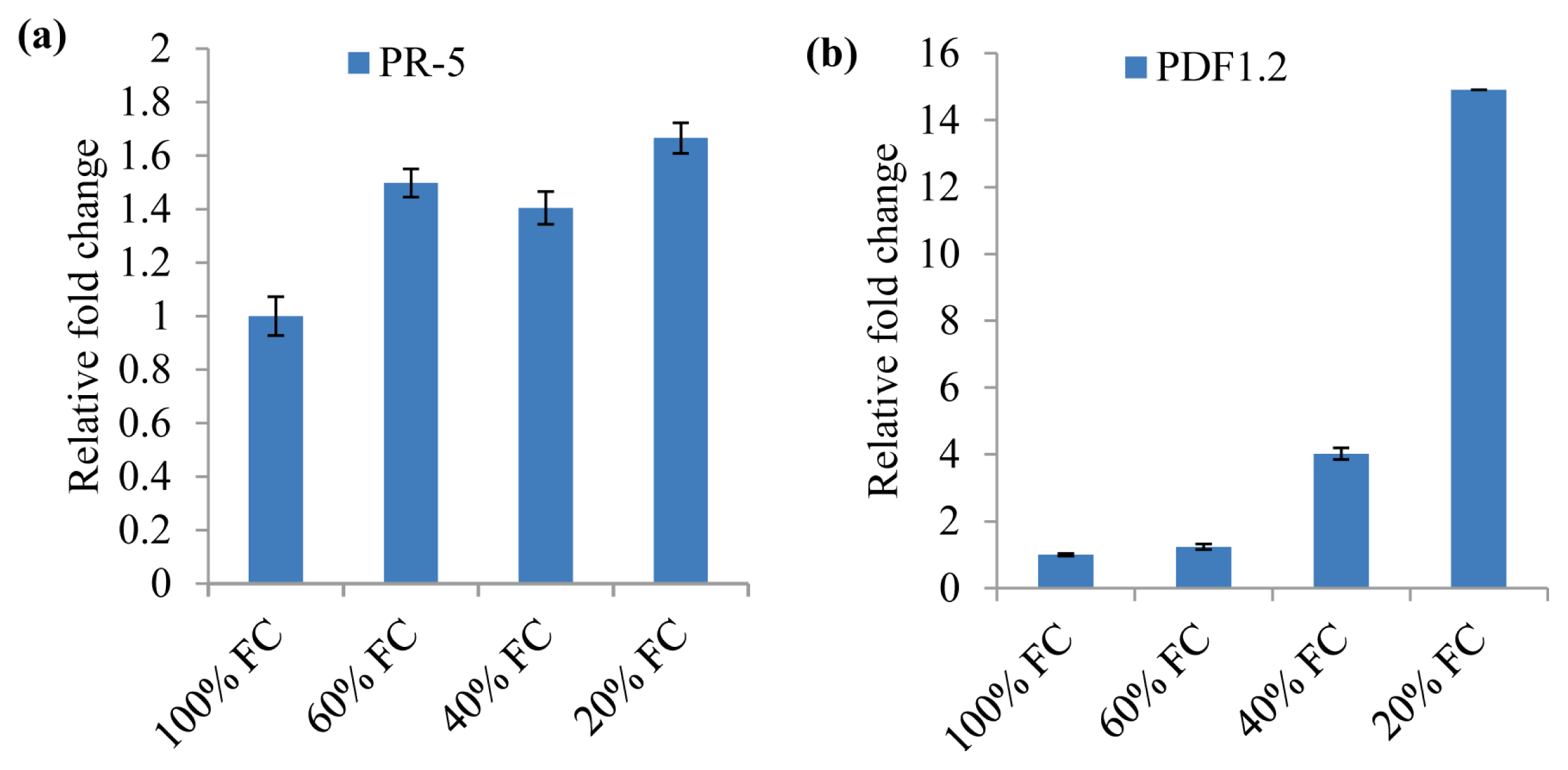
2.5. Drought Acclimation in N. benthamiana Induces Plant Defense Genes
3. Experimental Section
3.1. Plant Growth Condition
3.2. Drought Stress Imposition
3.3. RWC and Root-Shoot Ratio Measurement
3.4. ABA Quantification
3.5. Pseudomonas syringae pv. tabaci Inoculation and Scoring the Extent of Disease-Induced Cell Death
3.6. Sclerotinia sclerotiorum Inoculum Preparation and Plant Inoculation
3.7. Quantification of ROS in Drought Acclimated Plants
3.8. Inoculation and Monitoring Bacterial Growth in Methyl Viologen Pre-Treated Plants
3.9. Real-Time Quantitative RT-PCR (qRT-PCR) Analysis
4. Conclusion
Supplementary Information
ijms-14-09497-s001.pdfAcknowledgments
Conflict of Interest
References
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar]
- Hammond-Kosack, K.E.; Jones, J.D.G. Response to Plant Pethogens. In Biochemistry and Molecular Biology of Plants; Buchannan, B., Gruissem, W., Jones, R.L., Eds.; Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1102–1157. [Google Scholar]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol 2004, 134, 1683–1696. [Google Scholar]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot 2012, 63, 3523–3543. [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant. Biol 2006, 57, 781–803. [Google Scholar]
- Bostock, R.M. Signal crosstalk and induced resistance: Straddling the line between cost and benefit. Annu. Rev. Phytopathol 2005, 43, 545–580. [Google Scholar]
- Lorenzo, O.; Solano, R. Molecular players regulating the jasmonate signalling network. Curr. Opin. Plant Biol 2005, 8, 532–540. [Google Scholar]
- Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. Responses to Abiotic Stresses; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1158–1203. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol 2000, 51, 463–499. [Google Scholar]
- Lang, V.; Palva, E.T. The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of Arabidopsis thaliana (L.) Heynh. Plant Mol. Biol 1992, 20, 951–962. [Google Scholar]
- Knight, H.; Brandt, S.; Knight, M.R. A history of stress alters drought calcium signalling pathways in Arabidopsis. Plant J 1998, 16, 681–687. [Google Scholar]
- Bridger, G.M.; Yang, W.; Falk, D.E.; McKersie, B.D. Cold acclimation increases tolerance of activated oxygen in winter cereals. J. Plant Physiol 1994, 144, 235–240. [Google Scholar]
- Gilmour, S.J.; Hajela, R.K.; Thomashow, M.F. Cold Acclimation in Arabidopsis thaliana. Plant Physiol 1988, 87, 745–750. [Google Scholar]
- Jennings, P.; Saltveit, M.E. Temperature effects on imbibition and germination of cucumber (Cucumis sativus) seeds. J. Am. Soc. Hortic. Sci 1994, 119, 464–467. [Google Scholar]
- Prasad, T.K.; Anderson, M.D.; Martin, B.A.; Stewart, C.R. Evidence for chilling-induced oxidative stress in maize seedlings and a regulatory role for hydrogen peroxide. Plant Cell 1994, 6, 65–74. [Google Scholar]
- Mantyla, E.; Lang, V.; Palva, E.T. Role of abscisic acid in drought-induced freezing tolerance, cold acclimation, and accumulation of LT178 and RAB18 proteins in Arabidopsis thaliana. Plant Physiol 1995, 107, 141–148. [Google Scholar]
- Wang, X.; Cai, J.; Jiang, D.; Liu, F.; Dai, T.; Cao, W. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat. J. Plant Physiol 2011, 168, 585–593. [Google Scholar]
- Senthil-Kumar, M.; Srikanthbabu, V.; Mohan Raju, B.; Ganeshkumar; Shivaprakash, N.; Udayakumar, M. Screening of inbred lines to develop a thermotolerant sunflower hybrid using the temperature induction response (TIR) technique: A novel approach by exploiting residual variability. J. Exp. Bot 2003, 54, 2569–2578. [Google Scholar]
- Jakab, G.; Ton, J.; Flors, V.; Zimmerli, L.; Metraux, J.P.; Mauch-Mani, B. Enhancing Arabidopsis salt and drought stress tolerance by chemical priming for its abscisic acid responses. Plant Physiol 2005, 139, 267–274. [Google Scholar]
- Conrath, U.; Pieterse, C.M.; Mauch-Mani, B. Priming in plant-pathogen interactions. Trends Plant Sci 2002, 7, 210–216. [Google Scholar]
- Kauss, H.; Krause, K.; Jeblick, W. Methyl jasmonate conditions parsley suspension cells for increased elicitation of phenylpropanoid defense responses. Biochem. Biophys. Res. Commun 1992, 189, 304–308. [Google Scholar]
- Kauss, H.; Jeblick, W. Pretreatment of parsley suspension cultures with salicylic acid enhances spontaneous and elicited production of H2O2. Plant Physiol 1995, 108, 1171–1178. [Google Scholar]
- Katz, V.A.; Thulke, O.U.; Conrath, U. A benzothiadiazole primes parsley cells for augmented elicitation of defense responses. Plant Physiol 1998, 117, 1333–1339. [Google Scholar]
- Zimmerli, L.; Jakab, G.; Metraux, J.P.; Mauch-Mani, B. Potentiation of pathogen-specific defense mechanisms in Arabidopsis by beta-aminobutyric acid. Proc. Natl. Acad. Sci. USA 2000, 97, 12920–12925. [Google Scholar]
- Ahn, I.P.; Kim, S.; Lee, Y.H.; Suh, S.C. Vitamin B1-induced priming is dependent on hydrogen peroxide and the NPR1 gene in Arabidopsis. Plant Physiol 2007, 143, 838–848. [Google Scholar]
- Borges, A.A.; Dobon, A.; Exposito-Rodriguez, M.; Jimenez-Arias, D.; Borges-Perez, A.; Casanas-Sanchez, V.; Perez, J.A.; Luis, J.C.; Tornero, P. Molecular analysis of menadione-induced resistance against biotic stress in Arabidopsis. Plant Biotechnol. J 2009, 7, 744–762. [Google Scholar]
- Zhang, S.; Yang, X.; Sun, M.; Sun, F.; Deng, S.; Dong, H. Riboflavin-induced priming for pathogen defense in Arabidopsis thaliana. J. Integr. Plant Biol 2009, 51, 167–174. [Google Scholar]
- Eshraghi, L.; Anderson, J.; Aryamanesh, N.; Shearer, B.; McComb, J.; Hardy, G.E.S.; O’Brien, P.A. Phosphite primed defence responses and enhanced expression of defence genes in Arabidopsis thaliana infected with Phytophthora cinnamomi. Plant Pathol 2011, 60, 1086–1095. [Google Scholar]
- Massoud, K.; Barchietto, T.; Le Rudulier, T.; Pallandre, L.; Didierlaurent, L.; Garmier, M.; Ambard-Bretteville, F.; Seng, J.M.; Saindrenan, P. Dissecting phosphite-induced priming in Arabidopsis infected with Hyaloperonospora arabidopsidis. Plant Physiol 2012, 159, 286–298. [Google Scholar]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol 2006, 9, 436–442. [Google Scholar]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol 2008, 180, 911–921. [Google Scholar]
- Sharma, Y.K.; Leon, J.; Raskin, I.; Davis, K.R. Ozone-induced responses in Arabidopsis thaliana: The role of salicylic acid in the accumulation of defense-related transcripts and induced resistance. Proc. Natl. Acad. Sci. USA 1996, 93, 5099–5104. [Google Scholar]
- Yalpani, N.; Enyedi, A.J.; Leon, J.; Raskin, I. UV light and ozone stimulate accumulation of salicylic acid, pathogenesis-related protein and virus resistance in tobacco. Planta 1994, 193, 372–376. [Google Scholar]
- Lamb, C.; Dixon, R.A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol 1997, 48, 251–275. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Torres, M.A.; Dangl, J.L. Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development. Curr. Opin. Plant Biol 2005, 8, 397–403. [Google Scholar]
- Laloi, C.; Apel, K.; Danon, A. Reactive oxygen signalling: The latest news. Curr. Opin. Plant Biol 2004, 7, 323–328. [Google Scholar]
- Guan, L.M.; Zhao, J.; Scandalios, J.G. Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J 2000, 22, 87–95. [Google Scholar]
- Doke, N. Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Plant Pathol 1983, 23, 345–357. [Google Scholar]
- Auh, C.K.; Murphy, T.M. Plasma membrane redox enzyme is involved in the synthesis of O2− and H2O2 by phytophthora elicitor-stimulated rose cells. Plant Physiol 1995, 107, 1241–1247. [Google Scholar]
- Grant, M.; Brown, I.; Adams, S.; Knight, M.; Ainslie, A.; Mansfield, J. The RPM1 plant disease resistance gene facilitates a rapid and sustained increase in cytosolic calcium that is necessary for the oxidative burst and hypersensitive cell death. Plant J 2000, 23, 441–450. [Google Scholar]
- Zhang, A.; Jiang, M.; Zhang, J.; Tan, M.; Hu, X. Mitogen-activated protein kinase is involved in abscisic acid-induced antioxidant defense and acts downstream of reactive oxygen species production in leaves of maize plants. Plant Physiol 2006, 141, 475–487. [Google Scholar]
- Davies, W.J.; Zhang, J.H. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Physiol. Plant Mol. Biol 1991, 42, 55–76. [Google Scholar]
- Westgate, M.E.; Boyer, J.S. Osmotic adjustment and the inhibition of leaf, root, stem and silk growth at low water potentials in maize. Planta 1985, 164, 540–549. [Google Scholar]
- Sharp, R.E.; Davies, W.J. Regulation of growth and development of plants growing with a restricted supply of water. In Plants Under Stress; Jones, H.G., Flowers, T.J., Jones, M.B., Eds.; Cambridge University Press: Cambridge, UK, 1989; pp. 71–93. [Google Scholar]
- Williams, B.; Kabbage, M.; Kim, H.J.; Britt, R.; Dickman, M.B. Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS Pathog 2011, 7, e1002107. [Google Scholar]
- Achuo, E.A.; Prinsen, E.; Höfte, M. Influence of drought, salt stress and abscisic acid on the resistance of tomato to Botrytis cinerea and Oidium neolycopersici. Plant Pathol 2006, 55, 178–186. [Google Scholar]
- Salle, A.; Ye, H.; Yart, A.; Lieutier, F. Seasonal water stress and the resistance of Pinus yunnanensis to a bark-beetle-associated fungus. Tree Physiol 2008, 28, 679–687. [Google Scholar]
- Gachomo, E.W.; Kotchoni, S.O. Extract from drought-stress leaves enhances disease resistance through induction of pathogenesis related proteins and accumulation of reactive molecules. Biotechnology 2008, 7, 273–279. [Google Scholar]
- Rojas, C.M.; Senthil-Kumar, M.; Wang, K.; Ryu, C.M.; Kaundal, A.; Mysore, K.S. Glycolate oxidase modulates reactive oxygen species-mediated signal transduction during nonhost resistance in Nicotiana benthamiana and Arabidopsis. Plant Cell 2012, 24, 336–352. [Google Scholar]
- Taguchi, F.; Suzuki, T.; Inagaki, Y.; Toyoda, K.; Shiraishi, T.; Ichinose, Y. The siderophore pyoverdine of Pseudomonas Syringae pv. tabaci 6605 is an intrinsic virulence factor in host tobacco infection. J. Bacteriol 2010, 192, 117–126. [Google Scholar]
- Wang, K.; Uppalapati, S.R.; Zhu, X.; Dinesh-Kumar, S.P.; Mysore, K.S. SGT1 positively regulates the process of plant cell death during both compatible and incompatible plant-pathogen interactions. Mol. Plant Pathol 2010, 11, 597–611. [Google Scholar]
- Uppalapati, S.R.; Ishiga, Y.; Ryu, C.M.; Ishiga, T.; Wang, K.; Noël, L.D.; Parker, J.E.; Mysore, K.S. SGT1 contributes to coronatine signaling and Pseudomonas syringae pv. tomato disease symptom development in tomato and Arabidopsis. New Phytol 2011, 189, 83–93. [Google Scholar]
- Asselbergh, B.; De Vleesschauwer, D.; Hofte, M. Global switches and fine-tuning-ABA modulates plant pathogen defense. Mol. Plant Microbe Interact 2008, 21, 709–719. [Google Scholar]
- Yasuda, M.; Ishikawa, A.; Jikumaru, Y.; Seki, M.; Umezawa, T.; Asami, T.; Maruyama-Nakashita, A.; Kudo, T.; Shinozaki, K.; Yoshida, S.; et al. Antagonistic interaction between systemic acquired resistance and the abscisic acid-mediated abiotic stress response in Arabidopsis. Plant Cell 2008, 20, 1678–1692. [Google Scholar]
- Kusajima, M.; Yasuda, M.; Kawashima, A.; Nojiri, H.; Yamane, H.; Nakajima, M.; Akutsu, K.; Nakashita, H. Suppressive effect of abscisic acid on systemic acquired resistance in tobacco plants. J. Gen. Plant Pathol 2010, 76, 161–167. [Google Scholar]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar]
- Asselbergh, B.; Curvers, K.; Franca, S.C.; Audenaert, K.; Vuylsteke, M.; Van Breusegem, F.; Hofte, M. Resistance to Botrytis cinerea in sitiens, an abscisic acid-deficient tomato mutant, involves timely production of hydrogen peroxide and cell wall modifications in the epidermis. Plant Physiol 2007, 144, 1863–1877. [Google Scholar]
- Ayres, P.G. The interaction between environmental stress injury and biotic disease physiology. Annu. Rev. Phytopathol 1984, 22, 53–75. [Google Scholar]
- Ton, J.; Flors, V.; Mauch-Mani, B. The multifaceted role of ABA in disease resistance. Trends Plant Sci 2009, 14, 310–317. [Google Scholar]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ 2010, 33, 453–467. [Google Scholar]
- Wang, K.; Kang, L.; Anand, A.; Lazarovits, G.; Mysore, K.S. Monitoring in planta bacterial infection at both cellular and whole-plant levels using the green fluorescent protein variant GFPuv. New Phytol 2007, 174, 212–223. [Google Scholar]
- Mehler, A.H. Studies on reactions of illuminated chloroplasts. I. Mechanism of the reduction of oxygen and other Hill reagents. Arch. Biochem. Biophys 1951, 33, 65–77. [Google Scholar]
- Martin, G.B. Functional analysis of plant disease resistance genes and their downstream effectors. Curr. Opin. Plant Biol 1999, 2, 273–279. [Google Scholar]
- Glazebrook, J. Genes controlling expression of defense responses in Arabidopsis—2001 status. Curr. Opin. Plant Biol 2001, 4, 301–308. [Google Scholar]
- Hammond-Kosack, K.E.; Jones, J.D. Resistance gene-dependent plant defense responses. Plant Cell 1996, 8, 1773–1791. [Google Scholar]
- Penninckx, I.A.; Eggermont, K.; Terras, F.R.; Thomma, B.P.; De Samblanx, G.W.; Buchala, A.; Métraux, J.P.; Manners, J.M.; Broekaert, W.F. Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar]
- Zhu, B.; Chen, T.H.; Li, P.H. Expression of three osmotin-like protein genes in response to osmotic stress and fungal infection in potato. Plant Mol. Biol 1995, 28, 17–26. [Google Scholar]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol 1996, 47, 377–403. [Google Scholar]
- Ramegowda, V.; Senthil-Kumar, M.; Nataraja, K.N.; Reddy, M.K.; Mysore, K.S.; Udayakumar, M. Expression of a finger millet transcription factor, EcNAC1, in tobacco confers abiotic stress-tolerance. PLoS One 2012, 7, e40397. [Google Scholar]
- Flower, D.J.; Ludlow, M.M. Contribution of osmotic adjustment to the dehydration tolerance of water-stressed pigeonpea (Cajanus cajan (L.) millsp.) leaves. Plant Cell Environ 1986, 9, 33–40. [Google Scholar]
- Sutherland, M.W.; Learmonth, B.A. The tetrazolium dyes MTS and XTT provide new quantitative assays for superoxide and superoxide dismutase. Free Radic. Res 1997, 27, 283–289. [Google Scholar]
- Schopfer, P.; Plachy, C.; Frahry, G. Release of reactive oxygen intermediates (superoxide radicals, hydrogen peroxide, and hydroxyl radicals) and peroxidase in germinating radish seeds controlled by light, gibberellin, and abscisic acid. Plant Physiol 2001, 125, 1591–1602. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]






© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ramegowda, V.; Senthil-Kumar, M.; Ishiga, Y.; Kaundal, A.; Udayakumar, M.; Mysore, K.S. Drought Stress Acclimation Imparts Tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana. Int. J. Mol. Sci. 2013, 14, 9497-9513. https://doi.org/10.3390/ijms14059497
Ramegowda V, Senthil-Kumar M, Ishiga Y, Kaundal A, Udayakumar M, Mysore KS. Drought Stress Acclimation Imparts Tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana. International Journal of Molecular Sciences. 2013; 14(5):9497-9513. https://doi.org/10.3390/ijms14059497
Chicago/Turabian StyleRamegowda, Venkategowda, Muthappa Senthil-Kumar, Yasuhiro Ishiga, Amita Kaundal, Makarla Udayakumar, and Kirankumar S. Mysore. 2013. "Drought Stress Acclimation Imparts Tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana" International Journal of Molecular Sciences 14, no. 5: 9497-9513. https://doi.org/10.3390/ijms14059497
APA StyleRamegowda, V., Senthil-Kumar, M., Ishiga, Y., Kaundal, A., Udayakumar, M., & Mysore, K. S. (2013). Drought Stress Acclimation Imparts Tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana. International Journal of Molecular Sciences, 14(5), 9497-9513. https://doi.org/10.3390/ijms14059497