The Role of microRNA in Gastric Malignancy

Abstract
:1. Introduction
2. Helicobacter pylori and miRNA
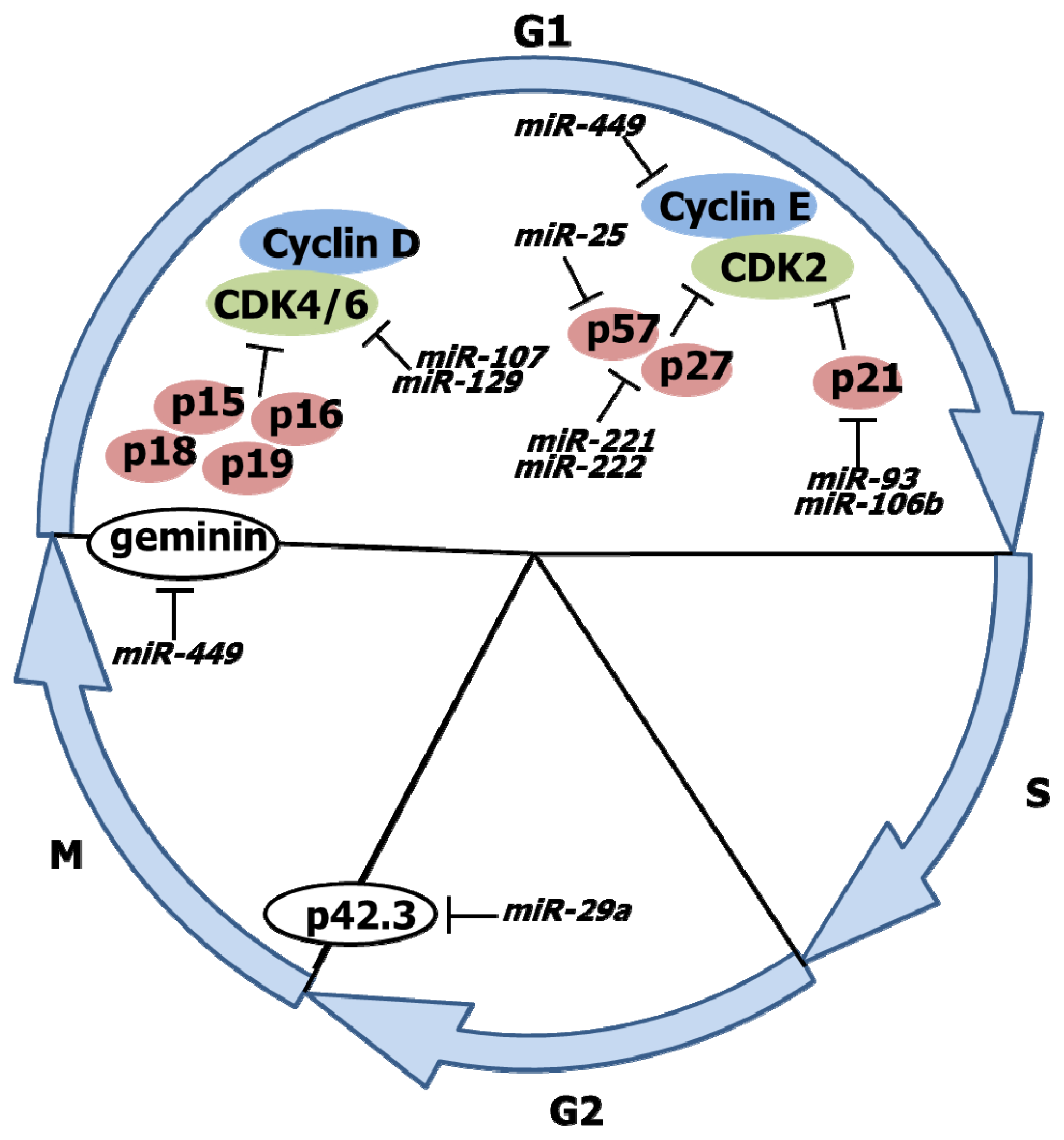
3. Cell Cycle Progression and miRNA
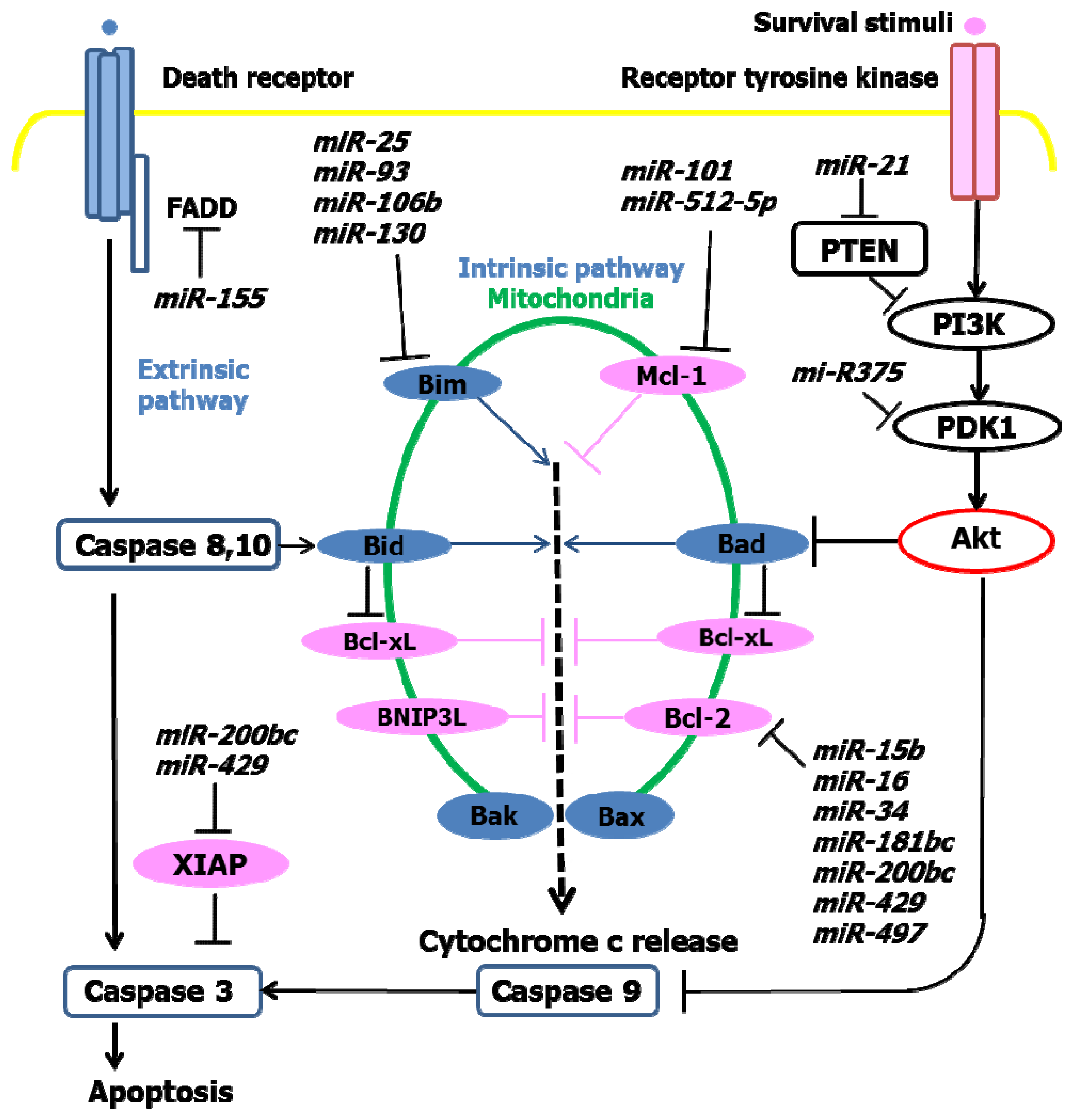
4. Inhibition of Apoptosis and miRNA
5. Metastasis and miRNA
6. miRNA and Anticancer Therapy
7. MALT Lymphoma and miRNA
8. Conclusion
Conflict of Interest
References
- Suzuki, H.; Iwasaki, E.; Hibi, T. Helicobacter pylori and gastric cancer. Gastric Cancer 2009, 12, 79–87. [Google Scholar]
- Suzuki, H.; Nishizawa, T.; Hibi, T. Helicobacter pylori eradication therapy. Future Microbiol 2010, 5, 639–648. [Google Scholar]
- Nishizawa, T.; Suzuki, H.; Nakagawa, I.; Minegishi, Y.; Masaoka, T.; Iwasaki, E.; Hibi, T. Early Helicobacter pylori eradication restores sonic hedgehog expression in the gastric mucosa of Mongolian gerbils. Digestion 2009, 79, 99–108. [Google Scholar]
- Fukase, K.; Kato, M.; Kikuchi, S.; Inoue, K.; Uemura, N.; Okamoto, S.; Terao, S.; Amagai, K.; Hayashi, S.; Asaka, M. Effect of eradication of Helicobacter pylori on incidence of metachronous gastric carcinoma after endoscopic resection of early gastric cancer: An open-label, randomised controlled trial. Lancet 2008, 372, 392–397. [Google Scholar]
- Saito, Y.; Suzuki, H.; Hibi, T. The role of microRNAs in gastrointestinal cancers. J. Gastroenterol 2009, 44, 18–22. [Google Scholar]
- Noto, J.M.; Peek, R.M. The role of microRNAs in Helicobacter pylori pathogenesis and gastric carcinogenesis. Front Cell Infect Microbiol 2011, 1, 21. [Google Scholar]
- Saito, Y.; Suzuki, H.; Tsugawa, H.; Suzuki, S.; Matsuzaki, J.; Hirata, K.; Hibi, T. Dysfunctional gastric emptying with down-regulation of muscle-specific microRNAs in Helicobacter pylori-infected mice. Gastroenterology 2011, 140, 189–198. [Google Scholar]
- Suzuki, H.; Nishizawa, T.; Tsugawa, H.; Mogami, S.; Hibi, T. Roles of oxidative stress in stomach disorders. J. Clin. Biochem. Nutr 2012, 50, 35–39. [Google Scholar]
- Tsugawa, H.; Suzuki, H.; Saya, H.; Hatakeyama, M.; Hirayama, T.; Hirata, K.; Nagano, O.; Matsuzaki, J.; Hibi, T. Reactive oxygen species-induced autophagic degradation of Helicobacter pylori caga is specifically suppressed in cancer stem-like cells. Cell Host Microbe 2012, 12, 764–777. [Google Scholar]
- Zhu, Y.; Jiang, Q.; Lou, X.; Ji, X.; Wen, Z.; Wu, J.; Tao, H.; Jiang, T.; He, W.; Wang, C.; et al. MicroRNAs up-regulated by CagA of Helicobacter pylori induce intestinal metaplasia of gastric epithelial cells. PLoS One 2012, 7, e35147. [Google Scholar]
- Shiotani, A.; Uedo, N.; Iishi, H.; Murao, T.; Kanzaki, T.; Kimura, Y.; Kamada, T.; Kusunoki, H.; Inoue, K.; Haruma, K. H. pylori eradication did not improve dysregulation of specific oncogenic miRNAs in intestinal metaplastic glands. J. Gastroenterol 2012, 47, 988–998. [Google Scholar]
- Xu, X.; Li, W.; Fan, X.; Liang, Y.; Zhao, M.; Zhang, J.; Tong, W.; Wang, J.; Yang, W.; Lu, Y. Identification and characterization of a novel p42.3 gene as tumor-specific and mitosis phase-dependent expression in gastric cancer. Oncogene 2007, 26, 7371–7379. [Google Scholar]
- Cui, Y.; Su, W.Y.; Xing, J.; Wang, Y.C.; Wang, P.; Chen, X.Y.; Shen, Z.Y.; Cao, H.; Lu, Y.Y.; Fang, J.Y. MiR-29a inhibits cell proliferation and induces cell cycle arrest through the downregulation of p42.3 in human gastric cancer. PLoS One 2011, 6, e25872. [Google Scholar]
- Kan, T.; Sato, F.; Ito, T.; Matsumura, N.; David, S.; Cheng, Y.; Agarwal, R.; Paun, B.C.; Jin, Z.; Olaru, A.V.; et al. The miR-106b-25 polycistron, activated by genomic amplification, functions as an oncogene by suppressing p21 and Bim. Gastroenterology 2009, 136, 1689–1700. [Google Scholar]
- Kim, Y.K.; Yu, J.; Han, T.S.; Park, S.Y.; Namkoong, B.; Kim, D.H.; Hur, K.; Yoo, M.W.; Lee, H.J.; Yang, H.K.; et al. Functional links between clustered microRNAs: Suppression of cell-cycle inhibitors by microRNA clusters in gastric cancer. Nucleic Acids Res 2009, 37, 1672–1681. [Google Scholar]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar]
- Xiao, B.; Liu, Z.; Li, B.S.; Tang, B.; Li, W.; Guo, G.; Shi, Y.; Wang, F.; Wu, Y.; Tong, W.D.; et al. Induction of microRNA-155 during Helicobacter pylori infection and its negative regulatory role in the inflammatory response. J. Infect Dis 2009, 200, 916–925. [Google Scholar]
- Zhu, W.; Xu, H.; Zhu, D.; Zhi, H.; Wang, T.; Wang, J.; Jiang, B.; Shu, Y.; Liu, P. miR-200bc/429 cluster modulates multidrug resistance of human cancer cell lines by targeting BCL2 and XIAP. Cancer Chemother. Pharmacol 2012, 69, 723–731. [Google Scholar]
- Tsukamoto, Y.; Nakada, C.; Noguchi, T.; Tanigawa, M.; Nguyen, L.T.; Uchida, T.; Hijiya, N.; Matsuura, K.; Fujioka, T.; Seto, M.; et al. MicroRNA-375 is downregulated in gastric carcinomas and regulates cell survival by targeting PDK1 and 14-3-3zeta. Cancer Res 2010, 70, 2339–2349. [Google Scholar]
- Guo, J.X.; Tao, Q.S.; Lou, P.R.; Chen, X.C.; Chen, J.; Yuan, G.B. miR-181b as a potential molecular target for anticancer therapy of gastric neoplasms. Asian Pac. J. Cancer Prev 2012, 13, 2263–2267. [Google Scholar]
- Gao, C.; Zhang, Z.; Liu, W.; Xiao, S.; Gu, W.; Lu, H. Reduced microRNA-218 expression is associated with high nuclear factor kappa B activation in gastric cancer. Cancer 2010, 116, 41–49. [Google Scholar]
- Tie, J.; Pan, Y.; Zhao, L.; Wu, K.; Liu, J.; Sun, S.; Guo, X.; Wang, B.; Gang, Y.; Zhang, Y.; et al. MiR-218 inhibits invasion and metastasis of gastric cancer by targeting the Robo1 receptor. PLoS Genet 2010, 6, e1000879. [Google Scholar]
- Li, B.S.; Zhao, Y.L.; Guo, G.; Zhu, E.D.; Luo, X.; Mao, X.H.; Zou, Q.M.; Yu, P.W.; Zuo, Q.F.; Li, N.; et al. Plasma microRNAs, miR-223, miR-21 and miR-218, as novel potential biomarkers for gastric cancer detection. PLoS One 2012, 7, e41629. [Google Scholar]
- Tseng, C.; Lin, C.C.; Chen, C.N.; Huang, H.C.; Juan, H.F. Integrative network analysis reveals active microRNA and their functions in gastric cancer. BioMed. Central Syst. Biol 2011, 5, 1–11. [Google Scholar]
- Kim, K.; Lee, H.C.; Park, J.L.; Kim, M.; Kim, S.Y.; Noh, S.M.; Song, K.S.; Kim, J.C.; Kim, Y.S. Epigenetic regulation of microRNA-10b and targeting of oncogenic MAPRE1 in gastric cancer. Epigenetics 2011, 6, 740–751. [Google Scholar]
- Suzuki, M.; Suzuki, H.; Minegishi, Y.; Ito, K.; Nishizawa, T.; Hibi, T. H. pylori-eradication therapy increases runx3 expression in the glandular epithelial cells in enlarged-fold gastritis. J. Clin. Biochem. Nutr 2010, 46, 259–264. [Google Scholar]
- Lai, K.W.; Koh, K.X.; Loh, M.; Tada, K.; Subramaniam, M.M.; Lim, X.Y.; Vaithilingam, A.; Salto-Tellez, M.; Iacopetta, B.; Ito, Y.; et al. MicroRNA-130b regulates the tumour suppressor RUNX3 in gastric cancer. Eur. J. Cancer 2010, 46, 1456–1463. [Google Scholar]
- Wang, M.; Li, C.; Yu, B.; Su, L.; Li, J.; Ju, J.; Yu, Y.; Gu, Q.; Zhu, Z.; Liu, B. Overexpressed miR-301a promotes cell proliferation and invasion by targeting RUNX3 in gastric cancer. J. Gastroenterol. 2013, in press. [Google Scholar]
- Saito, Y.; Suzuki, H.; Imaeda, H.; Matsuzaki, J.; Hirata, K.; Tsugawa, H.; Hibino, S.; Kanai, Y.; Saito, H.; Hibi, T. The tumor suppressor microRNA-29c is downregulated and restored by celecoxib in human gastric cancer cells. Int. J. Cancer 2013, 132, 1751–1760. [Google Scholar]
- Saito, Y.; Suzuki, H.; Tsugawa, H.; Imaeda, H.; Matsuzaki, J.; Hirata, K.; Hosoe, N.; Nakamura, M.; Mukai, M.; Saito, H.; et al. Overexpression of miR-142-5p and miR-155 in Gastric Mucosa-Associated Lymphoid Tissue (MALT) lymphoma resistant to Helicobacter pylori eradication. PLoS One 2012, 7, e47396. [Google Scholar]
- Craig, V.J.; Cogliatti, S.B.; Rehrauer, H.; Wundisch, T.; Muller, A. Epigenetic silencing of microRNA-203 dysregulates ABL1 expression and drives Helicobacter-associated gastric lymphomagenesis. Cancer Res 2011, 71, 3616–3624. [Google Scholar]
- Craig, V.J.; Cogliatti, S.B.; Imig, J.; Renner, C.; Neuenschwander, S.; Rehrauer, H.; Schlapbach, R.; Dirnhofer, S.; Tzankov, A.; Muller, A. Myc-mediated repression of microRNA-34a promotes high-grade transformation of B-cell lymphoma by dysregulation of FoxP1. Blood 2011, 117, 6227–6236. [Google Scholar] [Green Version]



{kind=link}
{kind=link}
| miRNAs | Change | Target mRNAs | Biological process targeted |
|---|---|---|---|
| let-7a | ↓ | RAB40C | Cell cycle progression |
| HMGA2 | Invasion | ||
| let-7b/d/e/f | ↓ | HMGA2 | Invasion |
| miR101 | ↓ | MCL1 | Apoptosis |
| miR-106b | ↓ | p21 | Cell cycle progression |
| BIM | Apoptosis | ||
| miR-125a | ↓ | ERBB2 | Proliferation |
| miR-141 | ↓ | FGFR2 | Proliferation |
| miR-200a | ↓ | ZEB1, ZEB2 | Epithelial to mesenchymal transition |
| miR-200b/c | ↓ | BCL2, XIAP | Apoptosis |
| miR-203 | ↓ | ABL1 | Proliferation, Invasion |
| miR-204 | ↓ | EZR | Proliferation |
| miR-218 | ↓ | ROBO1 | Invasion, Metastasis |
| miR-375 | ↓ | PDK1, 12-3-3 | Apoptosis |
| JAK2 | Proliferation | ||
| miR-429 | ↓ | BCL2, XIAP | Apoptosis |
| MYC | Proliferation | ||
| miR-17 | ↑ | p21 | Cell cycle progression |
| miR-20a | ↑ | p21 | Cell cycle progression |
| miR-21 | ↑ | PTEN | Proliferation |
| RECK | Metastasis | ||
| miR-146a | ↑ | IRAK1, TRAF6 | Proliferation, Immune response |
| SMAD4 | Apoptosis | ||
| miR-155 | ↑ | IKK-ɛ, SMAD4 | Immune response |
| FADD, PLIα | Apoptosis | ||
| miR-223 | ↑ | EPB41L3 | Invasion, Metastasis |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nishizawa, T.; Suzuki, H. The Role of microRNA in Gastric Malignancy. Int. J. Mol. Sci. 2013, 14, 9487-9496. https://doi.org/10.3390/ijms14059487
Nishizawa T, Suzuki H. The Role of microRNA in Gastric Malignancy. International Journal of Molecular Sciences. 2013; 14(5):9487-9496. https://doi.org/10.3390/ijms14059487
Chicago/Turabian StyleNishizawa, Toshihiro, and Hidekazu Suzuki. 2013. "The Role of microRNA in Gastric Malignancy" International Journal of Molecular Sciences 14, no. 5: 9487-9496. https://doi.org/10.3390/ijms14059487
APA StyleNishizawa, T., & Suzuki, H. (2013). The Role of microRNA in Gastric Malignancy. International Journal of Molecular Sciences, 14(5), 9487-9496. https://doi.org/10.3390/ijms14059487