Novel Single Nucleotide Polymorphisms of the Insulin-Like Growth Factor-I Gene and Their Associations with Growth Traits in Common Carp (Cyprinus carpio L.)


Abstract
:1. Introduction
2. Results
2.1. Identification and Genotyping of the Polymorphisms

2.2. Genotypic and Allelic Frequencies


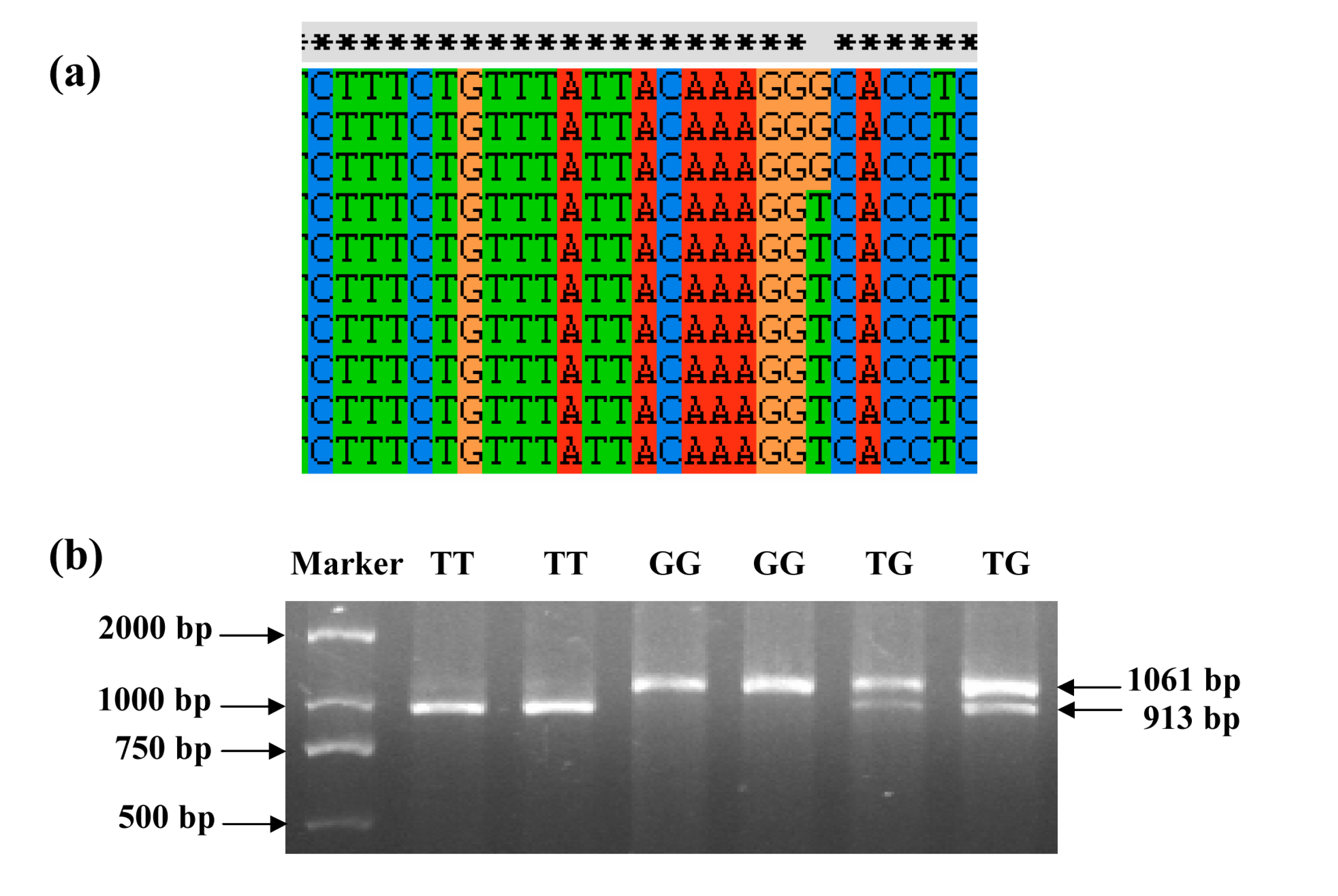{kind=link}
{kind=link}
{kind=link}
| Loci | Allelic Frequency | Genotypic Frequency | PIC | HO | HE | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| g.3759T>G | T | 0.957 | TT | 0.927 | 0.079 | 0.059 | 0.083 | 0.071 |
| G | 0.043 | TG | 0.059 | |||||
| GG | 0.014 | |||||||
| g.7627T>A | T | 0.763 | TT | 0.637 | 0.296 | 0.253 | 0.301 | 0.105 |
| A | 0.237 | TA | 0.252 | |||||
| AA | 0.111 | |||||||
| g.7722T>C | T | 0.486 | TT | 0.332 | 0.375 | 0.308 | 0.501 | 0.000 * |
| C | 0.514 | TC | 0.308 | |||||
| CC | 0.360 | |||||||
| g.7892C>T | C | 0.799 | CC | 0.685 | 0.269 | 0.228 | 0.362 | 0.002 * |
| T | 0.201 | CT | 0.228 | |||||
| TT | 0.087 | |||||||
2.3. Association Analysis of the SNPs with Growth Traits
| Genotypes | N | BL (cm) | BW (g) | K |
|---|---|---|---|---|
| TT | 184 | 18.4 ± 0.9 A | 138 ± 14 A | 2.21 ± 0.16 |
| TA | 73 | 18.5 ± 1.0 A,B | 140 ± 15 A,B | 2.19 ± 0.17 |
| AA | 32 | 19.0 ± 0.8 B | 146 ± 17 B | 2.15 ± 0.13 |
| Haplotype Combinations | N | BL (cm) | BW (g) | K |
|---|---|---|---|---|
| H1H1 (TT/TT/CC) | 47 | 18.4 ± 0.8 | 137 ± 13 | 2.19 ± 0.14 |
| H1H2 (TT/TT/CT) | 24 | 18.4 ± 0.9 | 137 ± 15 | 2.21 ± 0.15 |
| H1H3 (TT/TC/CC) | 31 | 18.3 ± 1.1 | 137 ± 17 | 2.26 ± 0.22 a |
| H1H7 (TA/TC/CC) | 15 | 18.2 ± 1.0 | 136 ± 13 | 2.28 ± 0.24 a |
| H2H2 (TT/TT/TT) | 23 | 18.5 ± 0.8 | 139 ± 14 | 2.18 ± 0.13 |
| H2H3 (TT/TC/TC) | 22 | 18.5 ± 0.7 | 139 ± 14 | 2.20 ± 0.13 |
| H2H7 (TA/TC/TC) | 20 | 18.8 ± 1.0 | 140 ± 16 | 2.10 ± 0.12 b |
| H3H3 (TT/CC/CC) | 40 | 18.2 ± 0.9 | 136 ± 15 | 2.24 ± 0.16 a |
| H3H7 (TA/CC/CC) | 33 | 18.7 ± 0.8 | 144 ± 13 | 2.19 ± 0.14 |
| H7H7 (AA/CC/CC) | 30 | 18.9 ± 0.8 | 146 ± 18 | 2.17 ± 0.13 |
| Combined SNPs | Combined Genotypes | N | BL (cm) | BW (g) | K |
|---|---|---|---|---|---|
| g.7627T>A and g.7722T>C | AA/CC | 29 | 18.9 ± 0.5 A | 145 ± 8 A | 2.15 ± 0.11 A |
| AT/CC | 37 | 19.0 ± 0.6 A | 147 ± 10 A | 2.13 ± 0.14 A | |
| AT/CT | 36 | 18.9 ± 0.6 A | 146 ± 9 A | 2.15 ± 0.12 A | |
| TT/CC | 38 | 18.9 ± 0.5 A | 147 ± 10 A | 2.17 ± 0.12 A | |
| TT/CT | 53 | 18.9 ± 0.5 A | 145 ± 11 A | 2.15 ± 0.09 A | |
| TT/TT | 93 | 17.5 ± 0.8 B | 125 ± 13 B | 2.31 ± 0.18 B | |
| g.7627T>A and g.7892C>T | AA/CC | 30 | 18.9 ± 0.9 a | 146 ± 18 a | 2.16 ± 0.13 |
| AT/CC | 52 | 18.5 ± 1.0 | 140 ± 15 | 2.23 ± 0.17 | |
| AT/CT | 21 | 18.8 ± 1.0 | 140 ± 16 | 2.11 ± 0.13 a | |
| TT/CC | 116 | 18.3 ± 0.9 b | 137 ± 15 b | 2.22 ± 0.17 b | |
| TT/CT | 45 | 18.5 ± 0.8 | 139 ± 14 | 2.21 ± 0.14 | |
| TT/TT | 23 | 18.5 ± 0.8 | 139 ± 14 | 2.18 ± 0.13 |
3. Discussion
4. Experimental Section
4.1. Experimental Fish and Phenotypic Data
4.2. Genomic DNA Extraction
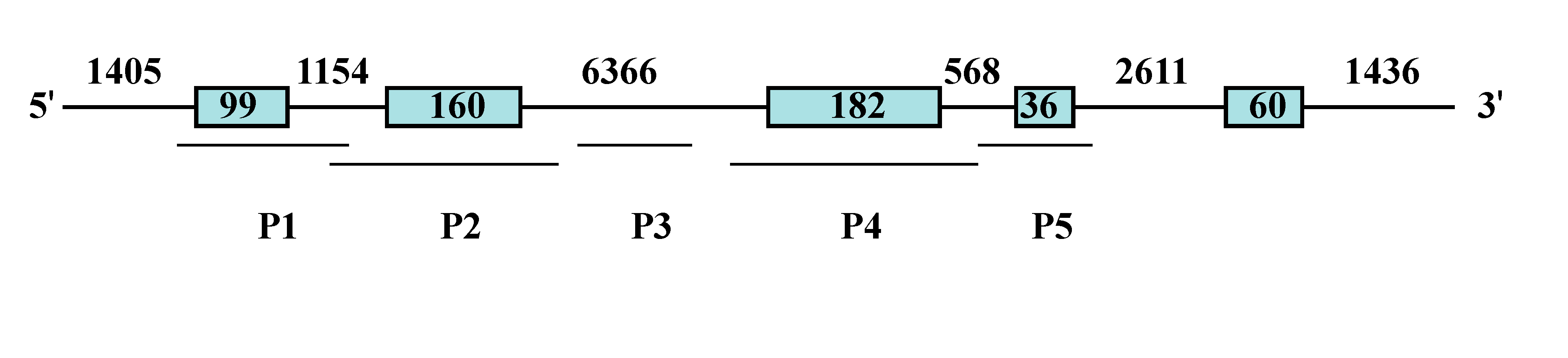
4.3. PCR Amplification and DNA Sequencing
| Primer Pairs | Primer Sequences (5'–3') | Product Length (bp) | Position |
|---|---|---|---|
| P1 | F: CAAATCCGTCTCCTGTTC | 822 | Exon 1 |
| R: ATACTGCTGCTTGAACCC | |||
| P2 | F: TTGAAGCATACTTGTGCGTTGT | 1346 | Exon 2 |
| R: AGTGTGATTGAAGGGAAGGTTT | |||
| P3 | F: GCACAATGGCTCAAGGAAGT | 1061 | Intron 2 |
| R: GTTTGTATCTGGGGAATGGG | |||
| P4 | F: AACTCAATGTGATGACAGGGT | 727 | Exon 3 |
| R: GCAACCATCCACCATCTATT | |||
| P5 | F: AATAGATGGTGGATGGTTGC | 599 | Exon 4 |
| R: CGCTGAGTTTAGAGTTTGGC |

4.4. SNP Detection and Genotyping
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ruane, J.; Colleau, J.J. Marker-assisted selection for a sex-limited character in a nucleus breeding population. J. Dairy Sci. 1996, 79, 1666–1678. [Google Scholar]
- Dekkers, J.C. Commercial application of marker- and gene-assisted selection in livestock: Strategies and lessons. J. Anim. Sci. 2004, 82, E313–328. [Google Scholar]
- Tao, W.J.; Boulding, E.G. Associations between single nucleotide polymorphisms in candidate genes and growth rate in Arctic charr (Salvelinus alpinus L.). Heredity 2003, 91, 60–69. [Google Scholar]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits, 1st ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 1998; p. 980. [Google Scholar]
- Buyse, J.; Decuypere, E. The role of the somatotrophic axis in the metabolism of the chicken. Domest. Anim. Endocrinol. 1999, 17, 245–255. [Google Scholar]
- Almuly, R.; Poleg-Danin, Y.; Gorshkov, S.; Gorshkova, G.; Rapoport, B.; Soller, M.; Kashi, Y.; Funkenstein, B. Characterization of the 5' flanking region of the growth hormone gene of the marine teleost, gilthead sea bream Sparus aurata: Analysis of a polymorphic microsatellite in the proximal promoter. Fish. Sci. 2005, 71, 479–490. [Google Scholar]
- Gross, R.; Nilsson, J. Restriction fragment length polymorphism at the growth hormone 1 gene in Atlantic salmon (Salmo salar L.) and its association with weight among the offspring of a hatchery stock. Aquaculture 1999, 173, 73–80. [Google Scholar]
- Tian, C.; Yang, M.; Lv, L.; Yuan, Y.; Liang, X.; Guo, W.; Song, Y.; Zhao, C. Single nucleotide polymorphisms in growth hormone gene and their association with growth traits in Siniperca chuatsi (Basilewsky). Int. J. Mol. Sci. 2014, 15, 7029–7036. [Google Scholar]
- Liu, L.S.; Yu, X.M; Tong, J.G. Molecular characterization of myostatin (MSTN) gene and association analysis with growth traits in the bighead carp (Aristichthys nobilis). Mol. Biol. Rep. 2012, 39, 9211–9221. [Google Scholar]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar]
- Vaessen, N.; Heutink, P.; Janssen, J.A.; Witteman, J.C.M.; Testers, L.; Hofman, A.; Lamberts, S.W.J.; Oostra, B.A.; Pols, H.A.P.; van Duijn, C.M. A polymorphism in the gene for IGF-I functional properties and risk for type 2 diabetes and myocardial infarction. Diabetes 2001, 50, 637–642. [Google Scholar]
- Rosen, C.J.; Kurland, E.S.; Vereault, D.; Adler, R.A.; Rackoff, P.J.; Craig, W.Y.; Witte, S.; Rogers, J.; Bilezikian, J.P. Association between serum insulin growth factor-I (IGF-I) and a simple sequence repeat in IGF-I gene: Implications for genetic studies of bone mineral density. J. Clin. Endocr. Metab. 1998, 83, 2286–2290. [Google Scholar]
- Bleumink, G.S.; Rietveld, I.; Janssen, J.A.M.J.L.; van Rossum, E.F.C.; Deckers, J.W.; Hofman, A.; Witteman, J.C.M.; van Duijn, C.M.; Stricker, B.H.C. Insulin-like growth factor-I gene polymorphism and risk of heart failure (the Rotterdam study). Am. J. Cardiol. 2004, 94, 384–386. [Google Scholar]
- Vaessen, N.; Janssen, J.A.; Heutink, P.; Hofman, A.; Lamberts, S.W.J.; Oostra, B.A.; Pols, H.A.P.; van Duijn, C.M. Association between genetic variation in the gene for insulin-like growth factor-I and low birthweight. Lancet 2002, 359, 1036–1037. [Google Scholar]
- Yu, H.; Li, B.D.L.; Smith, M.; Shi, R.; Berkel, H.J.; Kato, I. Polymorphic CA repeats in the IGF-I gene and breast cancer. Breast Cancer Res. Treat. 2001, 70, 117–122. [Google Scholar]
- Estany, J.; Tor, M.; Villalba, D.; Bosch, L.; Gallardo, D.; Jimenez, N.; Altet, L.; Noguera, J.L.; Reixach, J.; Amills, M.; et al. Association of CA repeat polymorphism at intron 1 of insulin-like growth factor (IGF-I) gene with circulating IGF-I concentration, growth, and fatness in swine. Physiol. Genomics 2007, 31, 236–243. [Google Scholar]
- Wang, W.J.; Ouyang, K.; Ouyang, J.; Li, H.H.; Lin, S.M.; Sun, H. Polymorphism of insulin-like growth factor I gene in six chicken breeds and its relationship with growth traits. Asian. Austral. J. Anim. 2004, 17, 301–304. [Google Scholar]
- Chung, E.R.; Kim, W.T. Association of SNP marker in IGF-1 and MYF5 candidate genes with growth traits in Korean cattle. Asian Australas. J. Anim. 2005, 18, 1061–1065. [Google Scholar]
- Li, X.H.; Bai, J.J.; Ye, X.; Hu, Y.C.; Li, S.J.; Yu, L.Y. Polymorphisms in the 5' flanking region of the insulin-like growth factor I gene are associated with growth traits in largemouth bass Micropterus salmoides. Fish. Sci. 2009, 75, 351–358. [Google Scholar]
- Tsai, H.Y.; Hamilton, A.; Guy, D.R.; Houston, R.D. Single nucleotide polymorphisms in the insulin-like growth factor 1 (IGF1) gene are associated with growth-related traits in farmed Atlantic salmon. Anim. Genet. 2014, 45, 709–715. [Google Scholar]
- Yearbook of Fishery Statistics Summary Tables. Available online: ftp://ftp.fao.org/FI/STAT/summary/default.htm#aqua (accessed on 10 August 2014).
- Zhang, Y.; Xu, P.; Lu, C.Y.; Kuang, Y.Y.; Zhang, X.F.; Cao, D.C.; Li, C.; Chang, Y.M.; Hou, N.; Li, H.D.; et al. Genetic Linkage mapping and analysis of muscle fiber-related QTLs in common carp (Cyprinus carpio L.). Mar. Biotechnol. 2011, 13, 376–392. [Google Scholar]
- Laghari, M.Y.; Zhang, Y.; Lashari, P.; Zhang, X.; Xu, P.; Xin, B.; Sun, X. Quantitative trait loci (QTL) associated with growth rate trait in common carp (Cyprinus carpio). Aquac. Int. 2013, 21, 1373–1379. [Google Scholar]
- Vong, Q.P.; Chan, K.M.; Leung, K.; Cheng, C.H.K. Common carp insulin-like growth factor-I gene: Complete nucleotide sequence and functional characterization of the 5'-flanking region. Gene 2003, 322, 145–156. [Google Scholar]
- Hu, X.S.; Li, C.T.; Shi, L.Y. A novel 79-bp insertion/deletion polymorphism in 3'-flanking region of IGF-I gene is associated with growth-related traits in common carp (Cyprinus carpio L.). Aquac. Res. 2013, 44, 1632–1638. [Google Scholar]
- Wigginton, J.E.; Cutler, D.J.; Abecasis, G.R. A note on exact tests of Hardy-Weinberg equilibrium. Am. J. Hum. Genet. 2005, 76, 887–893. [Google Scholar]
- Rousset, F.; Raymond, M. Testing heterozygote excess and deficiency. Genetics 1995, 140, 1413–1419. [Google Scholar]
- Sun, J.J.; Zhang, C.L.; Chen, H.; Fang, X.T.; Jin, Q.J.; Chen, D.X.; Shi, X.Y.; Du, Y. Two novel SNPs in coding region of the caprine Fat-inducing transcript gene and their association with growth traits. Mol. Biol. Rep. 2010, 37, 485–490. [Google Scholar]
- Ferraz, J.B.; Pinto, L.F.; Meirelles, F.V.; Eler, J.P.; de Rezende, F.M.; Oliveira, E.C.; Almeida, H.B.; Woodward, B.; Nkrumah, D. Association of single nucleotide polymorphisms with carcass traits in Nellore cattle. Genet. Mol. Res. 2009, 8, 1360–1366. [Google Scholar]
- Vaiman, D.; Mercier, D.; Moazamigoudarzi, K.; Eggen, A.; Ciampolini, R.; Lepingle, A.; Velmala, R.; Kaukinen, J.; Varvio, S.L.; Martin, P.; et al. A set of 99 cattle microsatellites: Characterization, synteny mapping, and polymorphism. Mamm. Genome 1994, 5, 288–297. [Google Scholar]
- Chasman, D.; Adams, R.M. Predicting the functional consequences of non-synonymous single nucleotide polymorphisms: Structure-based assessment of amino acid variation. J. Mol. Biol. 2001, 307, 683–706. [Google Scholar]
- Nguyen, M.T.; Barnes, A.C.; Mather, P.B.; Li, Y.T.; Lyons, R.E. Single nucleotide polymorphisms in the actin and crustacean hyperglycemic hormone genes and their correlation with individual growth performance in giant freshwater prawn Macrobrachium rosenbergii. Aquaculture 2010, 301, 7–15. [Google Scholar]
- Machado, M.B.B.; Alencar, M.M.; Pereira, A.P.; Oliveira, H.N.; Casas, E.; Coutinho, L.L.; Regitano, L.C.A. QTL affecting body weight in a candidate region of cattle chromosome 5. Genet. Mol. Biol. 2003, 26, 259–265. [Google Scholar]
- Daly, M.J.; Rioux, J.D.; Schaffner, S.F.; Hudson, T.J.; Lander, E.S. High-resolution haplotype structure in the human genome. Nat. Genet. 2001, 29, 229–232. [Google Scholar]
- Tabor, H.K.; Risch, N.J.; Myers, R.M. Candidate-gene approaches for studying complex genetic traits: Practical considerations. Nat. Rev. Genet. 2002, 3, 391–397. [Google Scholar]
- De Bakker, P.I.; Yelensky, R.; Pe’er, I.; Gabriel, S.B.; Daly, M.J.; Altshuler, D. Efficiency and power in genetic association studies. Nat. Genet. 2005, 37, 1217–1223. [Google Scholar]
- García-Magariños, M.; López-de-Ullibarri, I.; Cao, R.; Salas, A. Evaluating the ability of tree-based methods and logistic regression for the detection of SNP-SNP interaction. Ann. Hum. Genet. 2009, 73, 360–369. [Google Scholar]
- Wang, J.Q.; Lui, H.; Po, H.; Fan, L. Influence of salinity on food consumption, growth and energy conversion efficiency of common carp (Cyprinus carpio) fingerlings. Aquaculture 1997, 148, 115–124. [Google Scholar]
- Nash, R.D.M.; Valencia, A.H.; Geffen, A.J. The origin of Fulton’s condition factor-setting the record straight. Fisheries 2006, 31, 236–238. [Google Scholar]
- Taggart, J.B.; Hynes, R.A.; Prodöuhl, P.A.; Ferguson, A. A simplified protocol for routine total DNA isolation from salmonid fishes. J. Fish. Biol. 1992, 40, 963–965. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar]
- Shi, Y.Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Yu, X.; Tong, J. Novel Single Nucleotide Polymorphisms of the Insulin-Like Growth Factor-I Gene and Their Associations with Growth Traits in Common Carp (Cyprinus carpio L.). Int. J. Mol. Sci. 2014, 15, 22471-22482. https://doi.org/10.3390/ijms151222471
Feng X, Yu X, Tong J. Novel Single Nucleotide Polymorphisms of the Insulin-Like Growth Factor-I Gene and Their Associations with Growth Traits in Common Carp (Cyprinus carpio L.). International Journal of Molecular Sciences. 2014; 15(12):22471-22482. https://doi.org/10.3390/ijms151222471
Chicago/Turabian StyleFeng, Xiu, Xiaomu Yu, and Jingou Tong. 2014. "Novel Single Nucleotide Polymorphisms of the Insulin-Like Growth Factor-I Gene and Their Associations with Growth Traits in Common Carp (Cyprinus carpio L.)" International Journal of Molecular Sciences 15, no. 12: 22471-22482. https://doi.org/10.3390/ijms151222471
APA StyleFeng, X., Yu, X., & Tong, J. (2014). Novel Single Nucleotide Polymorphisms of the Insulin-Like Growth Factor-I Gene and Their Associations with Growth Traits in Common Carp (Cyprinus carpio L.). International Journal of Molecular Sciences, 15(12), 22471-22482. https://doi.org/10.3390/ijms151222471