l-Cystathionine Inhibits the Mitochondria-Mediated Macrophage Apoptosis Induced by Oxidized Low Density Lipoprotein


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
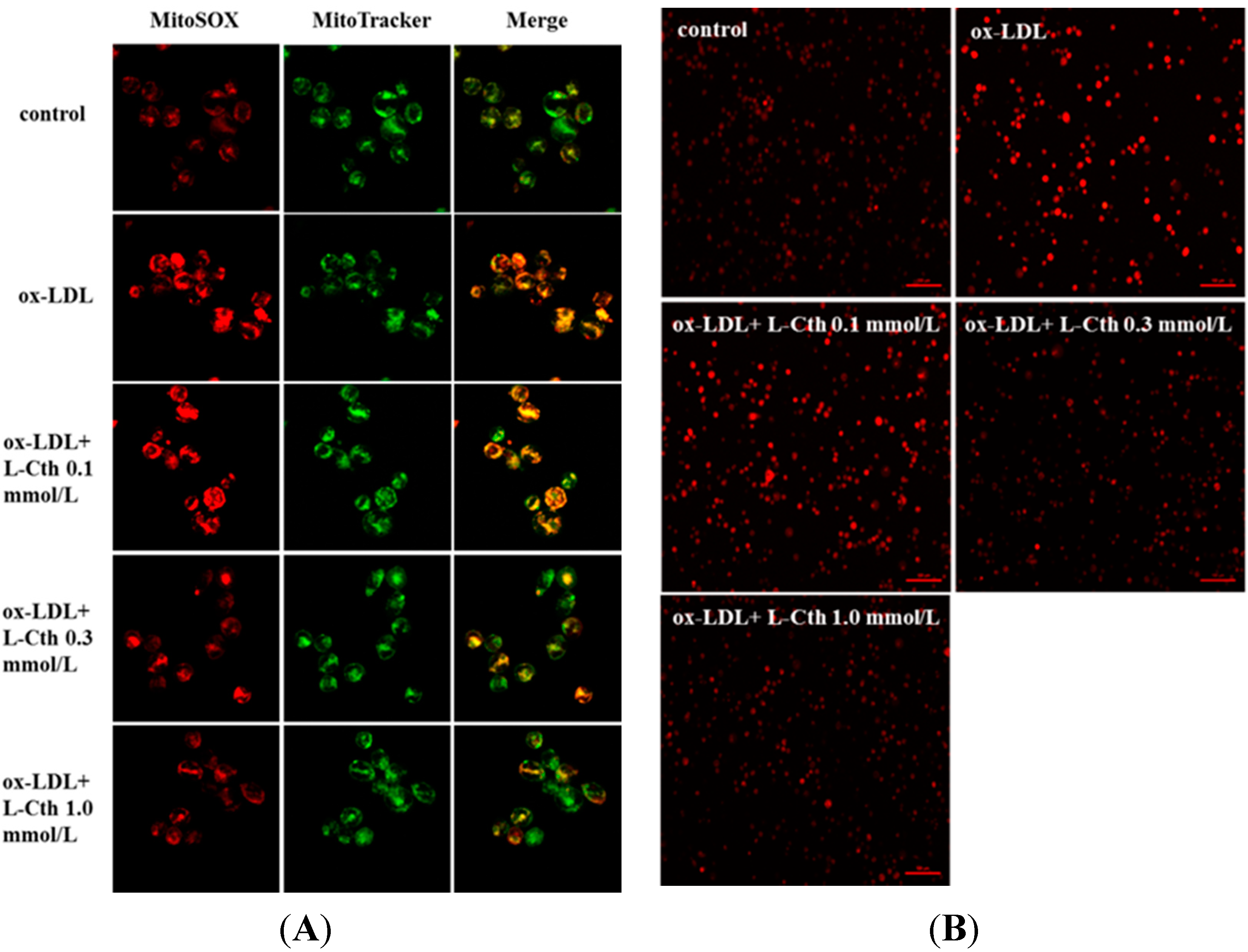
2.1. l-Cystathionine Inhibited Oxidative Stress Induced by ox-LDL in Human Macrophages

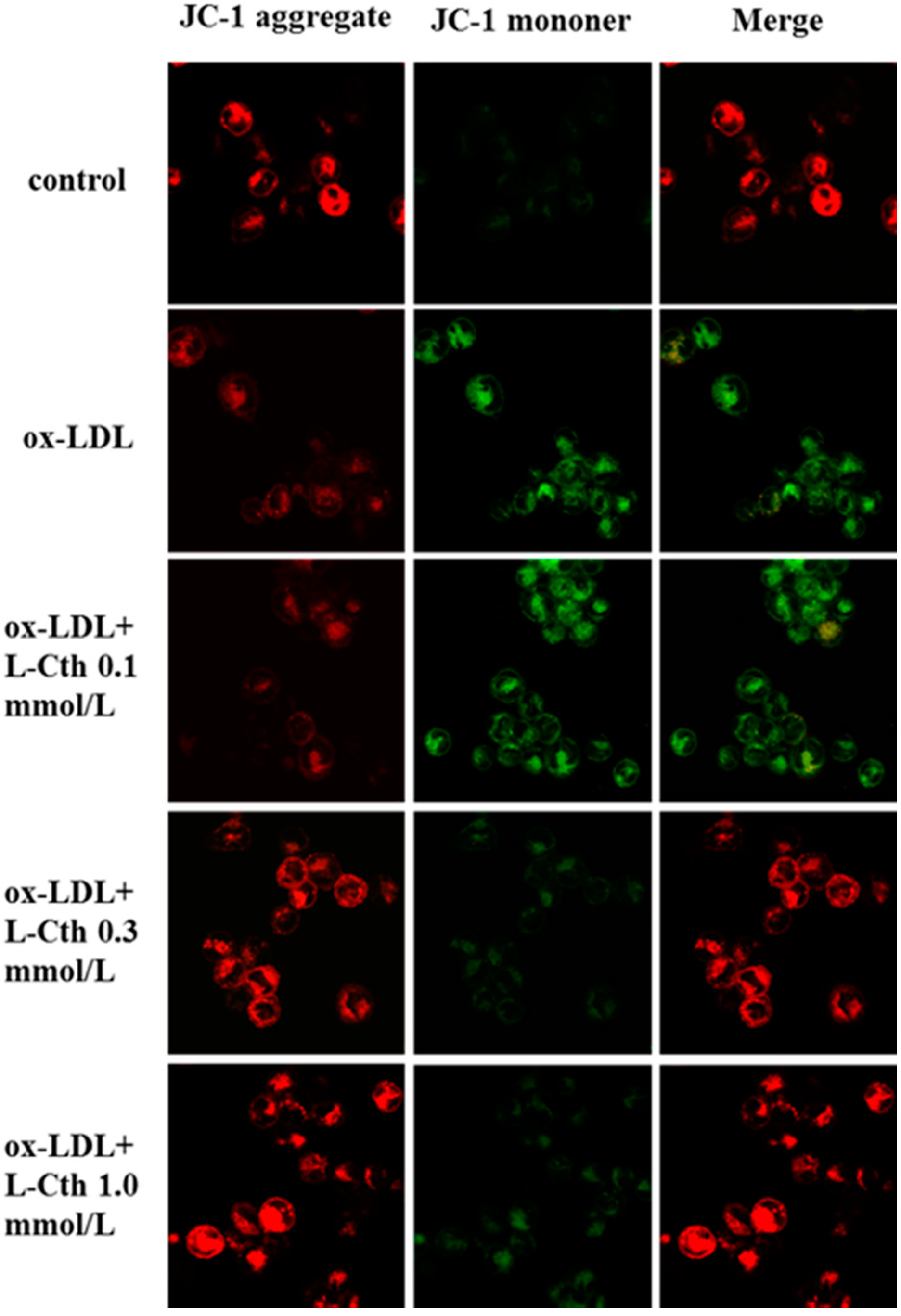
2.2. l-Cystathionine Reversed the Inhibition of Mitochondrial Membrane Potential Induced by ox-LDL in Human Macrophages

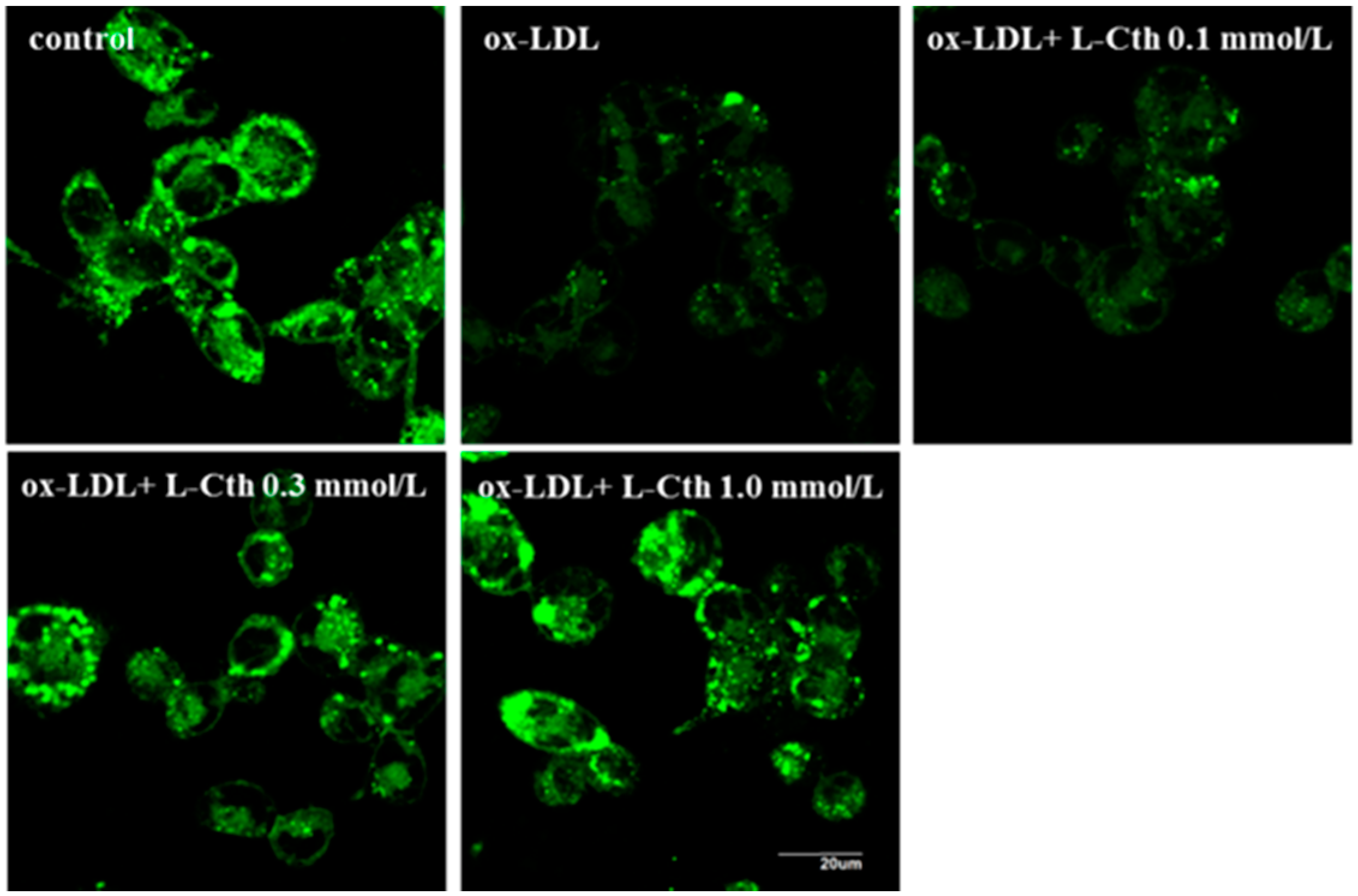
2.3. l-Cystathionine Antagonized Mitochondrial Permeability Transition Pore (MPTP) Opening Induced by ox-LDL in Human Macrophages

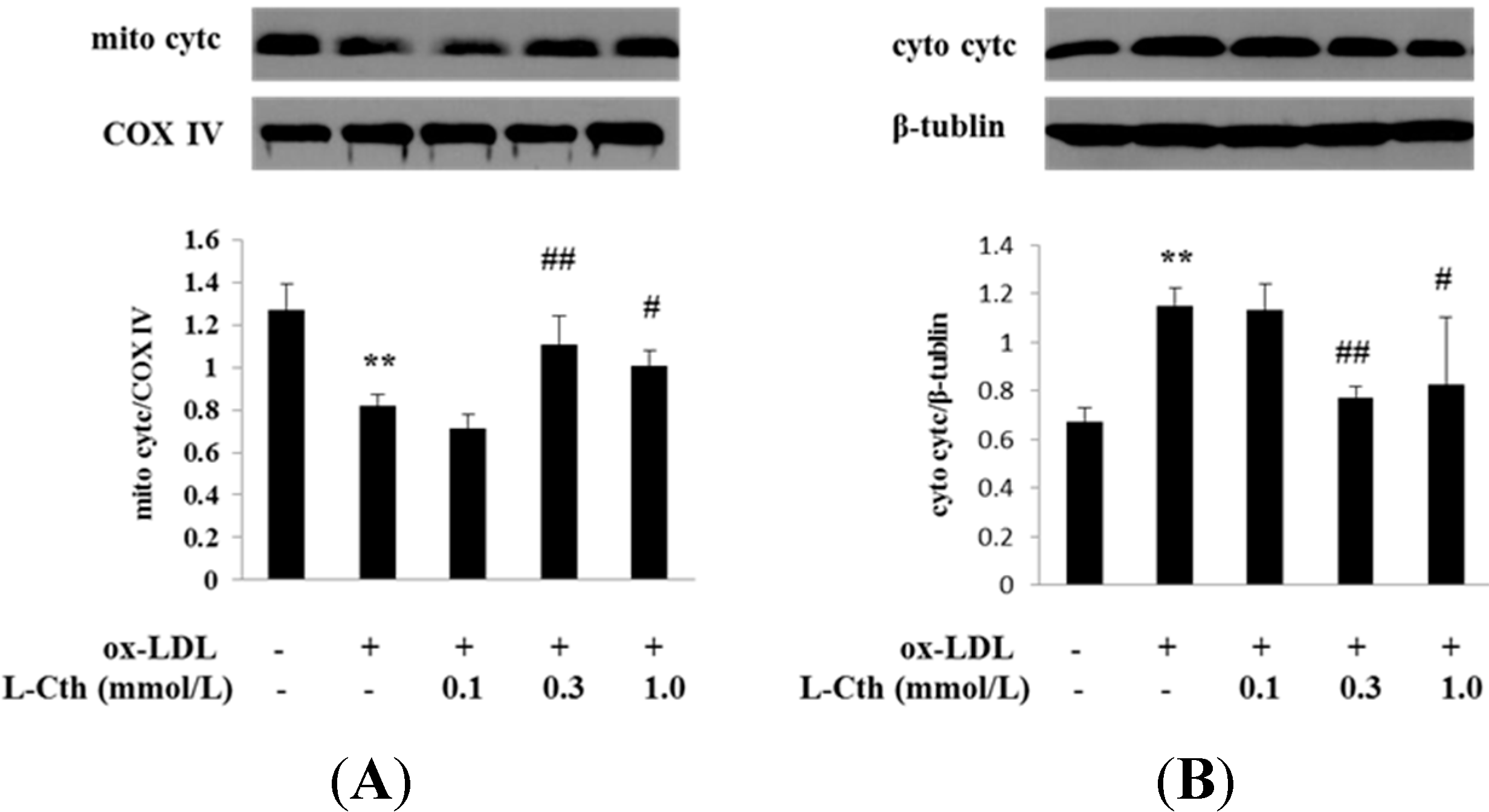
2.4. l-Cystathionine Inhibited the Release of Cytc from the Mitochondrion into the Cytoplasm Induced by ox-LDL in Human Macrophages

2.5. l-Cystathionine Suppressed Cell Apoptosis Induced by ox-LDL in Human Macrophages


3. Discussion

4. Materials and Methods
4.1. THP-1 Monocytes Culture
4.2. Detection of Mitochondrial Superoxide in Human Macrophages by MitoSOX Reagent
4.3. Detection of Superoxide in Human Macrophages by Dihydroethidium (DHE)
4.4. Measurement of Mitochondrial Membrane Potential in Human Macrophages by JC-1
4.5. Measurement of MPTP Opening in Human Macrophages by Cell MPTP Assay Kit
4.6. Detection of the Release of Cytc from the Mitochondrion into the Cytoplasm in Human Macrophages by Western Blot
4.7. Measurement of Caspase-9 Activities in Human Macrophages by Fluorescence and Colorimetric Assay
4.8. Cleavage of Caspase-3 in Human Macrophage Examined by Western Blotting
4.9. Detection of Cell Apoptosis in Human Macrophages by TUNEL Assay
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Seimon, T.; Tabas, I. Mechanisms and consequences of macrophage apoptosis in atherosclerosis. J. Lipid Res. 2009, 50, S382–S387. [Google Scholar] [CrossRef] [PubMed]
- Tsukano, H.; Gotoh, T.; Endo, M.; Miyata, K.; Tazume, H.; Kadomatsu, T.; Yano, M.; Iwawaki, T.; Kohno, K.; Araki, K.; et al. The endoplasmic reticulum stress-C/EBP homologous protein pathway-mediated apoptosis in macrophages contributes to the instability of atherosclerotic plaques. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Thorp, E.; Li, G.; Seimon, T.A.; Kuriakose, G.; Ron, D.; Tabas, I. Reduced apoptosis and plaque necrosis in advanced atherosclerotic lesions of Apoe−/− and LDLR−/− mice lacking CHOP. Cell MeTable 2009, 9, 474–481. [Google Scholar] [CrossRef]
- Imanishi, T.; Akasaka, T. Novel strategies to target inflammatory processes in atherosclerosis. Curr. Pharm. Des. 2013, 19, 1616–1625. [Google Scholar] [PubMed]
- Tabas, I. Consequences and therapeutic implications of macrophage apoptosis in atherosclerosis: The importance of lesion stage and phagocytic efficiency. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Gillies, L.A.; Kuwana, T. Apoptosis regulation at the mitochondrial outer membrane. J. Cell. Biochem. 2014, 115, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Skender, B.; Hofmanová, J.; Slavík, J.; Jelínková, I.; Machala, M.; Moyer, M.P.; Kozubík, A.; Hyršlová Vaculová, A. DHA-mediated enhancement of TRAIL-induced apoptosis in colon cancer cells is associated with engagement of mitochondria and specific alterations in sphingolipid metabolism. Biochim. Biophys. Acta 2014, 1841, 1308–1397. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sato, E.F.; Kira, Y.; Nishikawa, M.; Utsumi, K.; Inoue, M. A possible cooperation of SOD1 and cytochrome c in mitochondria-dependent apoptosis. Free Radic. Biol. Med. 2006, 40, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, V.; Salvi, M.; Toninello, A. Oxidative stress is responsible for mitochondrial permeability transition induction by salicylate in liver mitochondria. J. Biol. Chem. 2005, 280, 33864–33872. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sitasawad, S.L. N-acetylcysteine prevents glucose/glucose oxidase-induced oxidative stress, mitochondrial damage and apoptosis in H9c2 cells. Life Sci. 2009, 84, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Gaull, G.; Sturman, J.A.; Räihä, N.C. Development of mammalian sulfur metabolism: Absence of cystathionase in human fetal tissues. Pediatr. Res. 1972, 6, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, M.; Ma, D.; Sugahara, K.; Kodama, H. Metabolism of cystathionine, N-monoacetylcystathionine, perhydro-1,4-thiazepine-3,5-dicarboxylic acid, and cystathionineketimine in the liver and kidney of d,l-propargylglycine-treated rats. Metabolism 1998, 47, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.M.; Kirsch, J.F. The enzymology of cystathionine biosynthesis: Strategies for the control of substrate and reaction specificity. Arch. Biochem. Biophys. 2005, 433, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sugahara, K.; Sagara, Y.; Fontana, M.; Duprè, S.; Kodama, H. Effect of cystathionine ketimine on the stimulus coupled responses of neutrophils and their modulation by various protein kinase inhibitors. Biochem. Biophys. Res. Commun. 1996, 218, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Zhang, J.; Sugahara, K. Novel priming compounds of cystathionine metabolites on superoxide generation in human neutrophils. Biochem. Biophys. Res. Commun. 2000, 269, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Kamisaki, Y.; Itoh, T. Hepatoprotective effects of cystathionine against acetaminophen-induced necrosis. J. Pharmacol. Exp. Ther. 1989, 250, 667–671. [Google Scholar] [PubMed]
- Kwiecien, N.; Michalska, M.; Wlodek, L. The selective effect of cystathionine on doxorubicin hepatotoxicity in tumor-bearing mice. Eur. J. Pharmacol. 2006, 550, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Maclean, K.N.; Greiner, L.S.; Evans, J.R.; Sood, S.K.; Lhotak, S.; Markham, N.E.; Stabler, S.P.; Allen, R.H.; Austin, R.C.; Balasubramaniam, V.; et al. Cystathionine protects against endoplasmic reticulum stress-induced lipid accumulation, tissue injury, and apoptotic cell death. J. Biol. Chem. 2012, 287, 31994–32005. [Google Scholar] [CrossRef] [PubMed]
- Ghibelli, L.; Fanelli, C.; Rotilio, G.; Lafavia, E.; Coppola, S.; Colussi, C.; Civitareale, P.; Ciriolo, M.R. Rescue of cells from apoptosis by inhibition of active GSH extrusion. FASEB J. 1998, 12, 479–486. [Google Scholar] [PubMed]
- Klein, C.E.; Roberts, B.; Holcenberg, J.; Glode, L.M. Cystathionine metabolism in neuroblastoma. Cancer 1988, 62, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Quéré, I.; Paul, V.; Rouillac, C.; Janbon, C.; London, J.; Demaille, J.; Kamoun, P.; Dufier, J.L.; Abitbol, M.; Chassé, J.F. Spatial and temporal expression of the cystathionine β-synthase gene during early human development. Biochem. Biophys. Res. Commun. 1999, 254, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Robert, K.; Vialard, F.; Thiery, E.; Toyama, K.; Sinet, P.M.; Janel, N.; London, J. Expression of the cystathionine β-synthase (CBS) gene during mouse development and immunolocalization in adult brain. J. Histochem. Cytochem. 2003, 51, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tang, C.S.; Jin, H.F.; Du, J.B. The vasorelaxing effect of hydrogen sulfide on isolated rat aortic rings versus pulmonary artery rings. Acta Pharmacol. Sin. 2011, 32, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Kouichirou, W.; Yoshinori, K.; Kentaro, N.; Tadao, I. Effect of cystathionine as a scavenger of superoxide generated from human leukocytes or derived from xanthine oxidase in vitro. Eur. J. Pharmacol. 1996, 296, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Caroppi, P.; Sinibaldi, F.; Fiorucci, L.; Santucci, R. Apoptosis and human diseases: Mitochondrion damage and lethal role of released cytochrome C as proapoptotic protein. Curr. Med. Chem. 2009, 16, 4058–4065. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.W.; Chen, J.T.; Hong, C.Y.; Lin, Y.L.; Wang, K.T.; Yao, C.J.; Lai, G.M.; Chen, R.M. Honokiol traverses the blood-brain barrier and induces apoptosis of neuroblastoma cells via an intrinsic bax-mitochondrion-cytochrome c-caspase protease pathway. Neurol. Oncol. 2012, 14, 302–314. [Google Scholar] [CrossRef]
- Guo, R.; Su, Y.; Liu, B.; Li, S.; Zhou, S.; Xu, Y. Resveratrol suppresses oxidised low-density lipoprotein-induced macrophage apoptosis through inhibition of intracellular reactive oxygen species generation, LOX-1, and the p38 MAPK pathway. Cell. Physiol. Biochem. 2014, 34, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Auwerx, J. The human leukemia cell line, THP-1: A multifacetted model for the study of monocyte-macrophage differentiation. Experientia 1991, 47, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, X.; Li, Y.; Leu, C.; Guo, L.; Zheng, X.; Zhu, D. 20-Hydroxyeicosatetraenoic acid inhibits the apoptotic responses in pulmonary artery smooth muscle cells. Eur. J. Pharmacol. 2008, 588, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Sun, H.; Liu, G.T. 5-Bromotetrandrine enhances the sensitivity of doxorubicin-induced apoptosis in intrinsic resistant human hepatic cancer Bel7402 cells. Cancer Lett. 2010, 292, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Du, J.; Chen, S.; Liu, A.D.; Holmberg, L.; Chen, Y.; Zhang, C.; Tang, C.; Jin, H. l-Cystathionine Inhibits the Mitochondria-Mediated Macrophage Apoptosis Induced by Oxidized Low Density Lipoprotein. Int. J. Mol. Sci. 2014, 15, 23059-23073. https://doi.org/10.3390/ijms151223059
Zhu M, Du J, Chen S, Liu AD, Holmberg L, Chen Y, Zhang C, Tang C, Jin H. l-Cystathionine Inhibits the Mitochondria-Mediated Macrophage Apoptosis Induced by Oxidized Low Density Lipoprotein. International Journal of Molecular Sciences. 2014; 15(12):23059-23073. https://doi.org/10.3390/ijms151223059
Chicago/Turabian StyleZhu, Mingzhu, Junbao Du, Siyao Chen, Angie Dong Liu, Lukas Holmberg, Yonghong Chen, Chunyu Zhang, Chaoshu Tang, and Hongfang Jin. 2014. "l-Cystathionine Inhibits the Mitochondria-Mediated Macrophage Apoptosis Induced by Oxidized Low Density Lipoprotein" International Journal of Molecular Sciences 15, no. 12: 23059-23073. https://doi.org/10.3390/ijms151223059
APA StyleZhu, M., Du, J., Chen, S., Liu, A. D., Holmberg, L., Chen, Y., Zhang, C., Tang, C., & Jin, H. (2014). l-Cystathionine Inhibits the Mitochondria-Mediated Macrophage Apoptosis Induced by Oxidized Low Density Lipoprotein. International Journal of Molecular Sciences, 15(12), 23059-23073. https://doi.org/10.3390/ijms151223059