When Isolated at Full Receptivity, in Vitro Fertilized Wheat (Triticum aestivum, L.) Egg Cells Reveal [Ca2+]cyt Oscillation of Intracellular Origin


Abstract
:
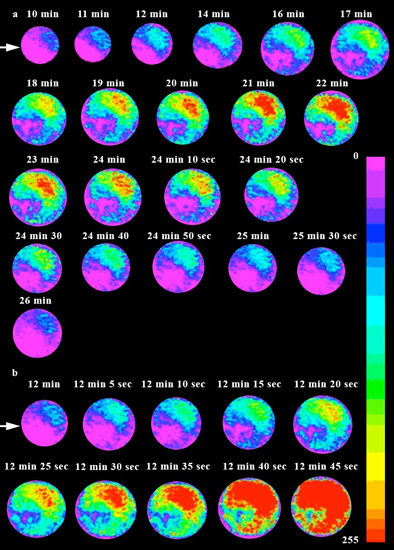{kind=link}
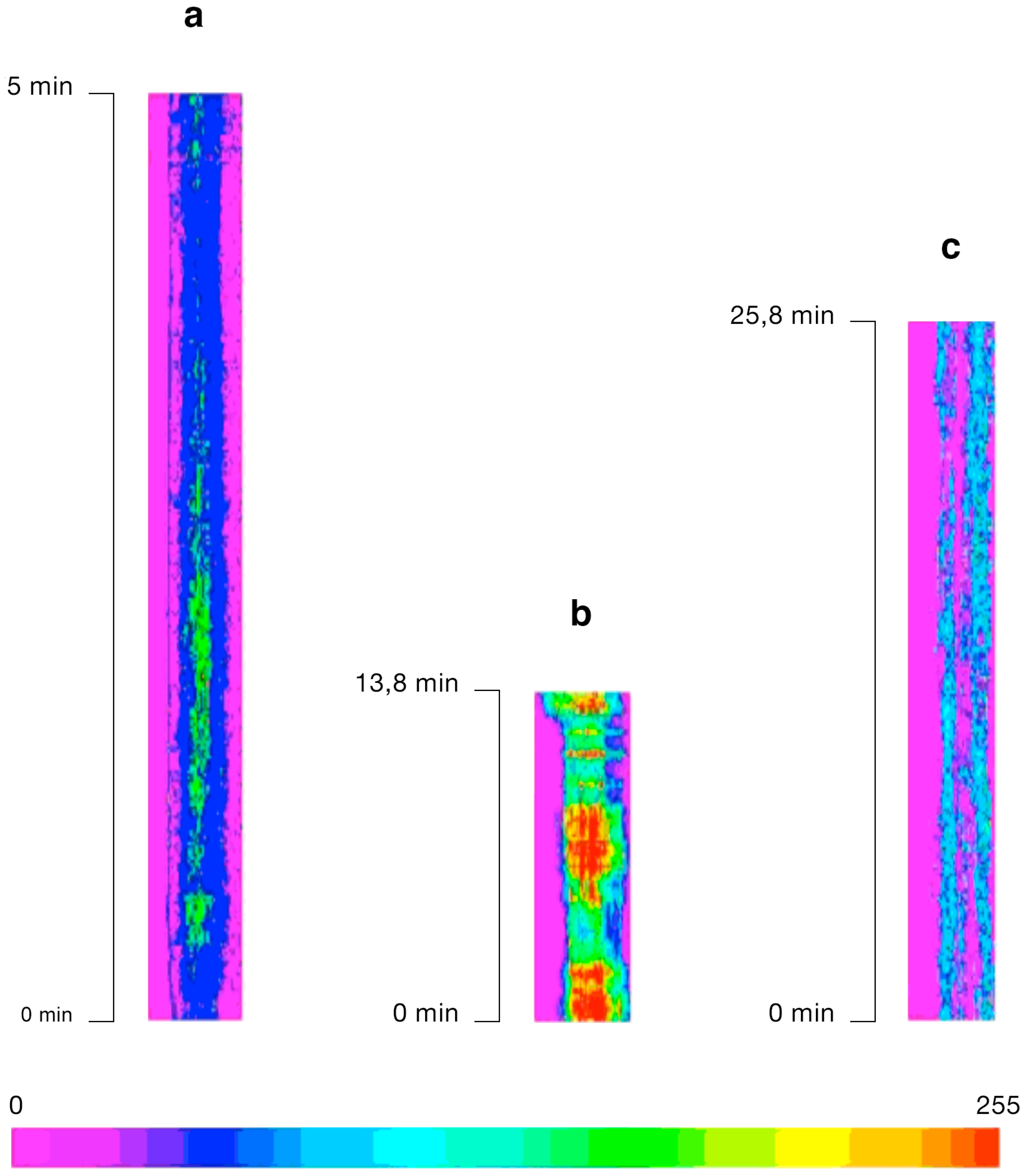{kind=link}
{kind=link}
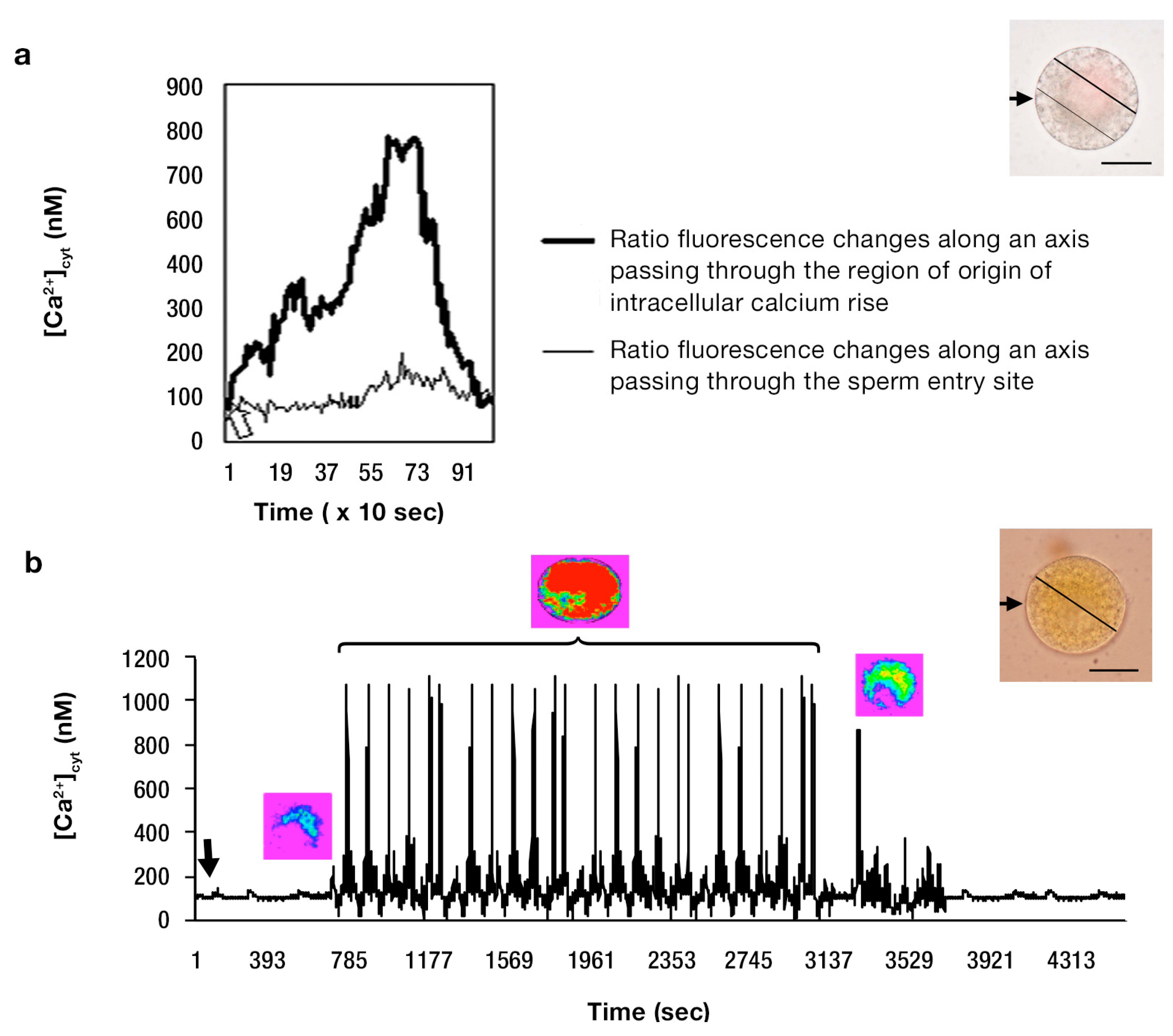{kind=link}
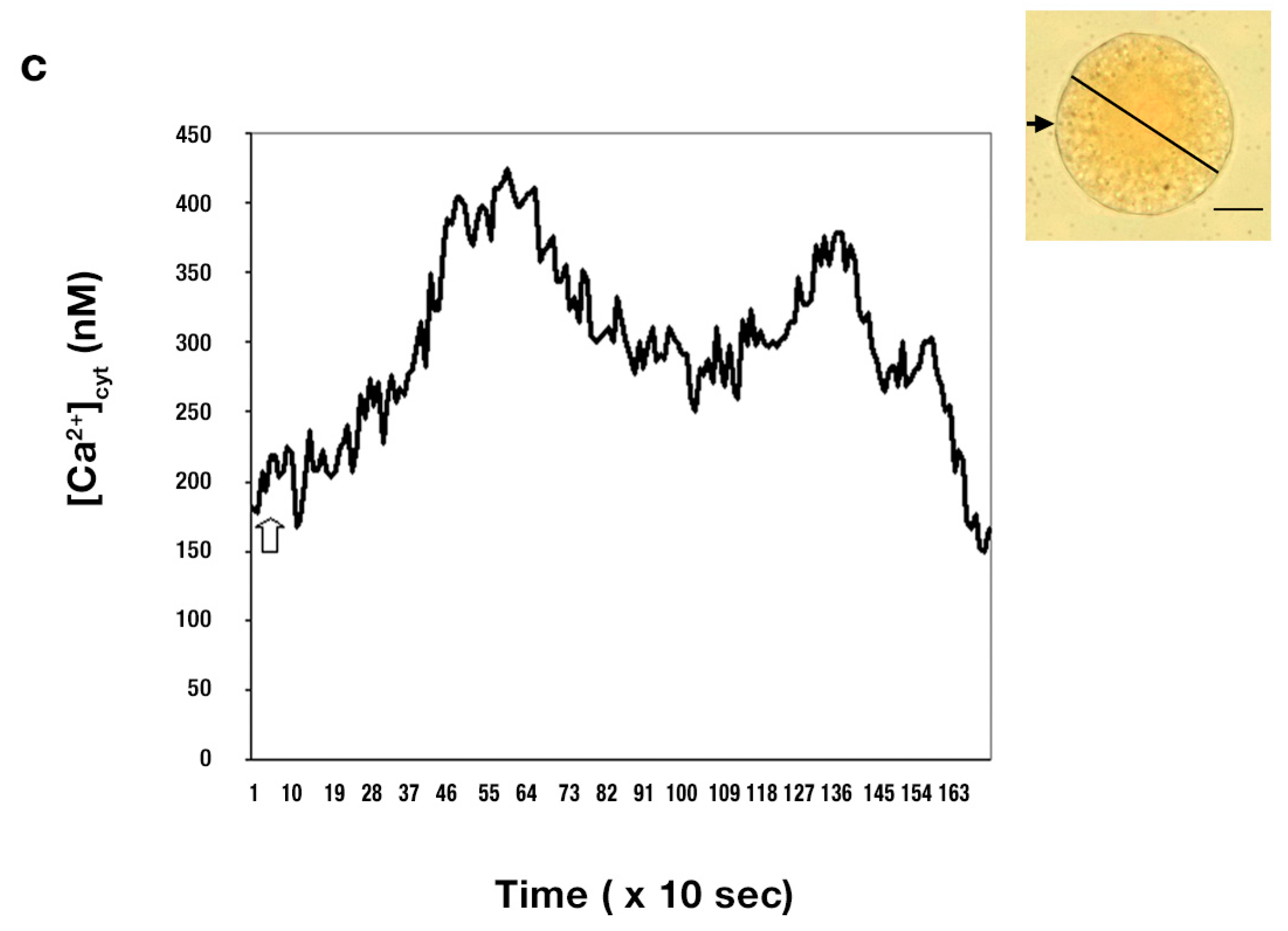{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
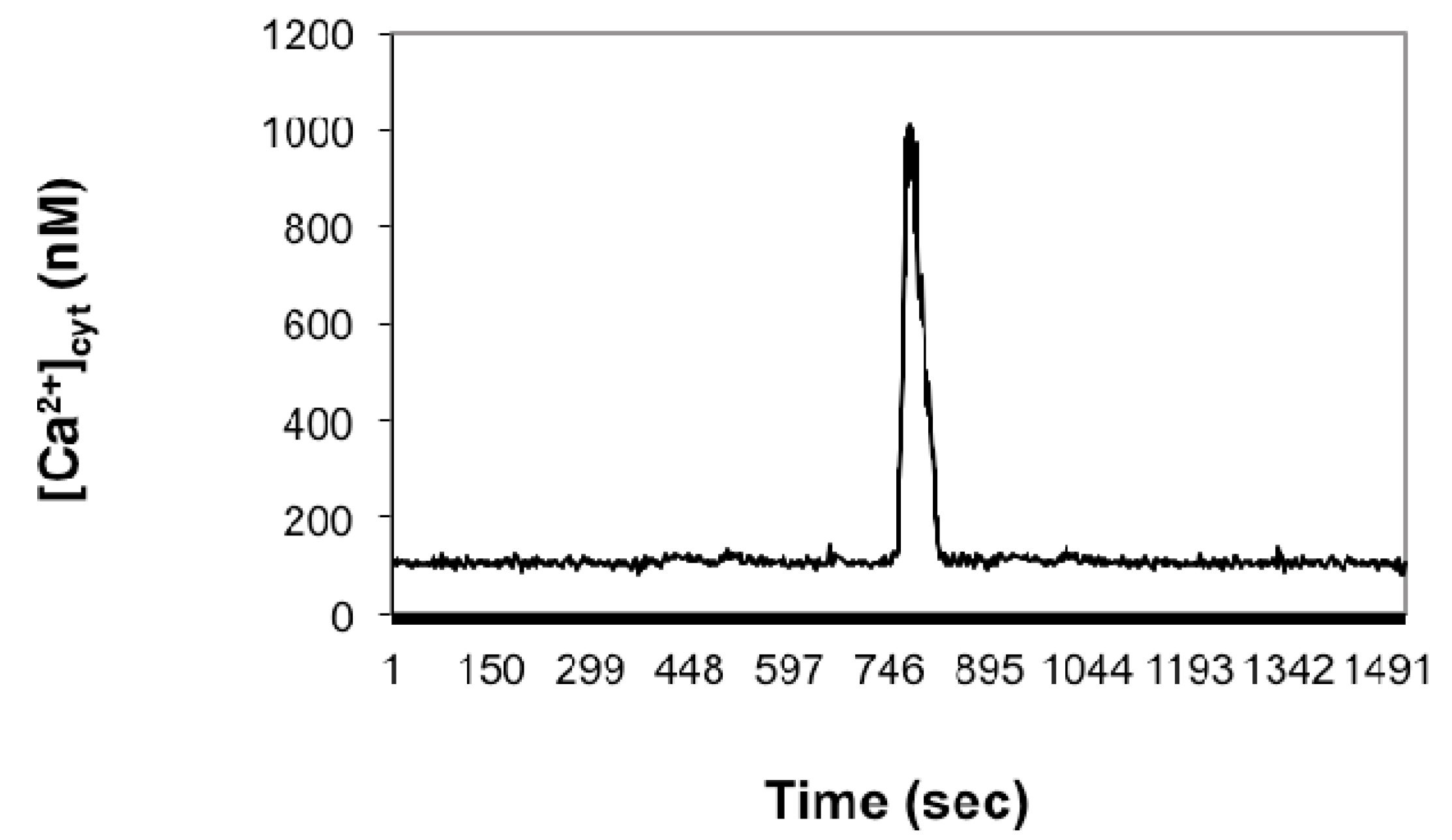
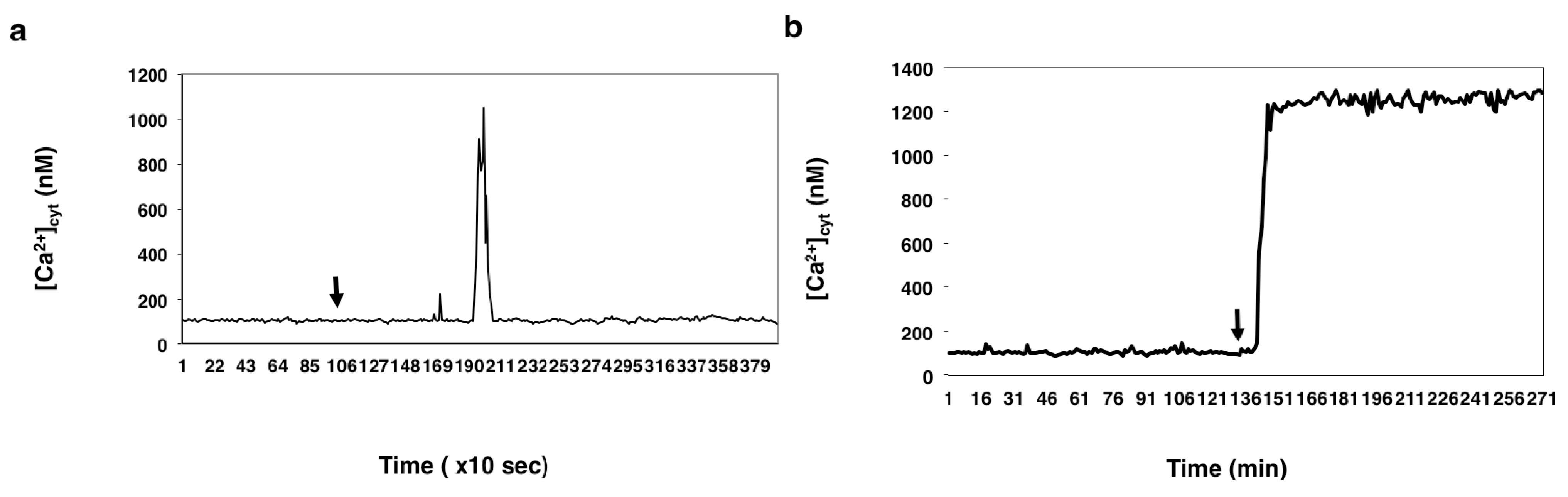
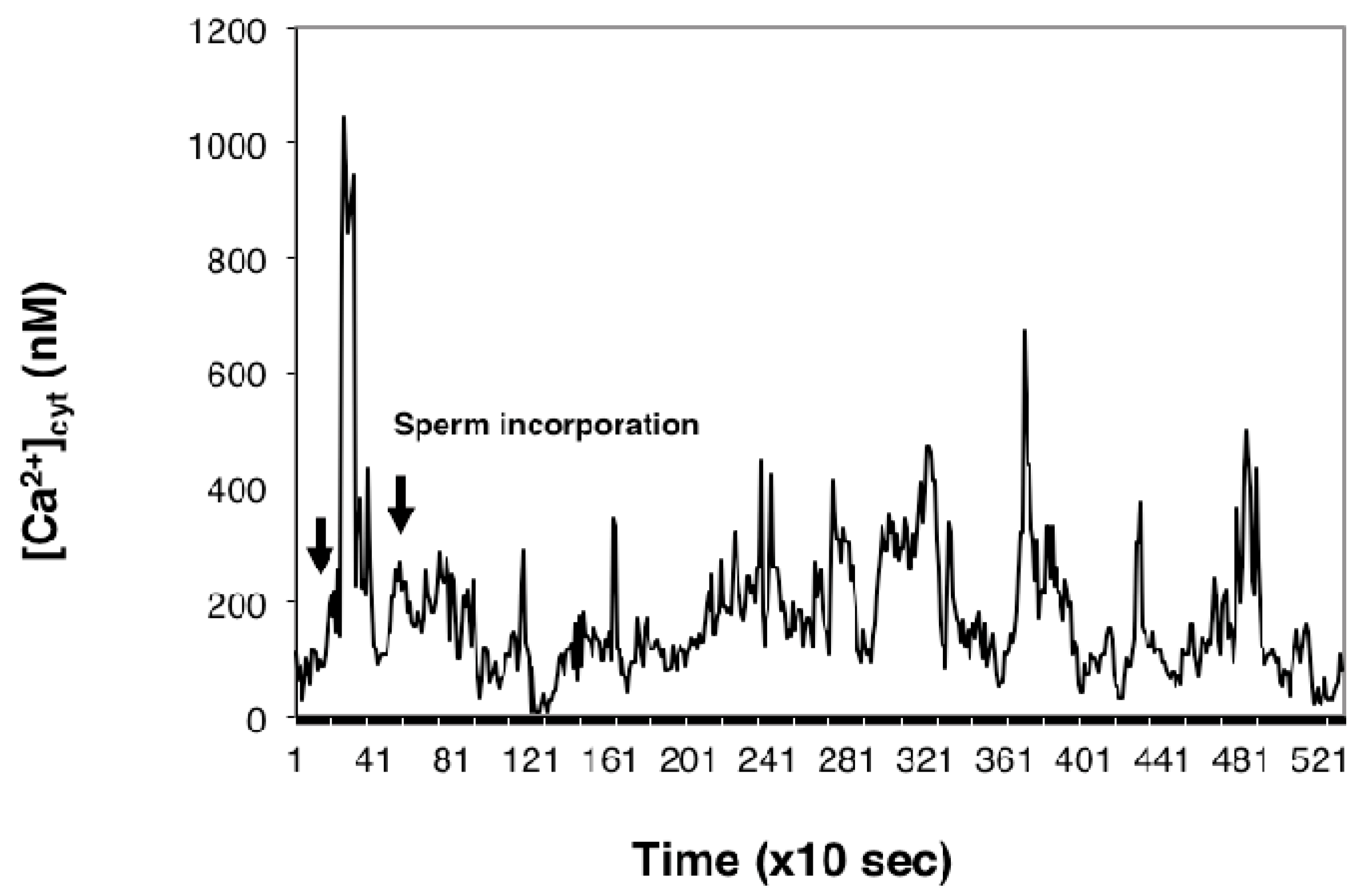
2.1. Imaging [Ca2+]cyt during in Vitro Fertilization (IVF) of Isolated Egg Cells Developed in Situ






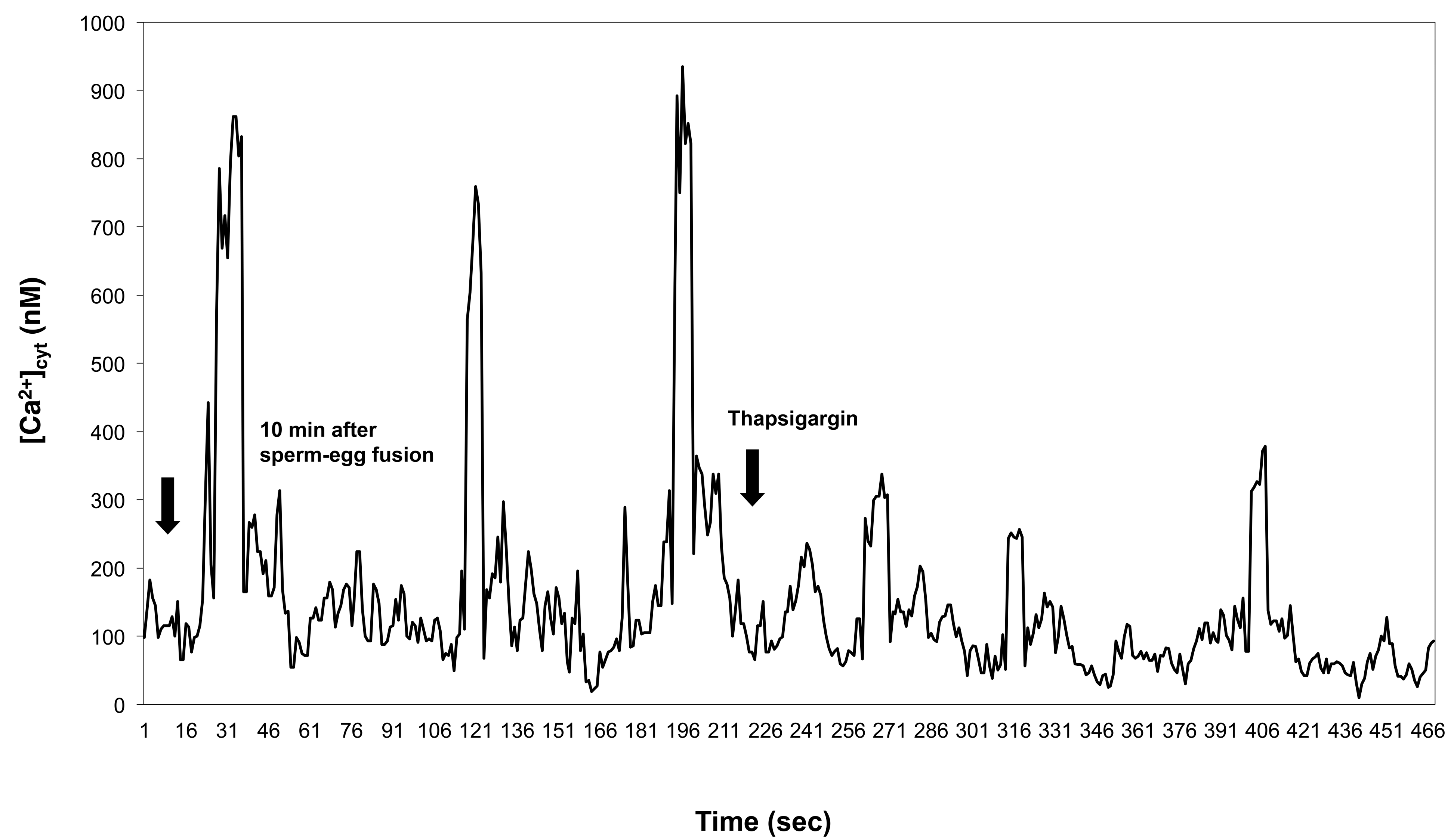
2.2. Effect upon [Ca2+]cyt of Thapsigargin Added to the IVF Medium







3. Discussion
3.1. [Ca2+]cyt Changes during IVF of Wheat Egg Cells
3.2. The Possible Origin of the [Ca2+]cyt Transients
- (1)
- In the unfertilized egg incubated in IVF medium without extracellular Ca2+, thapsigargin at a 10 µM concentration caused a transient increase in [Ca2+]cyt which per se had to originate from an intracellular calcium store.
- (2)
- Since thapsigargin is an irreversible inhibitor of calcium pumps, the single and rapidly decreasing [Ca2+]cyt rise (see Figure 6c,d) suggests that thapsigargin, added at concentrations of 10 and 50 µM, did not interact with the plasma membrane calcium pumps, or if yes, not to the extent that would have prevented them from functioning properly, i.e., pumping the “extra” calcium out of the cytosol; otherwise, the [Ca2+]cyt would have remained at a high level for a much longer time (due to the irreversible depletion of both the ER and PM Ca2+-ATPase). It may be reasoned, however, that other intracellular Ca2+ pumps, such as those located, e.g., in the Golgi apparatus membrane or on vacuole membranes, may have remained unaffected by thapsigargin, which could still facilitate sequestering Ca2+ into the Golgi apparatus or into vacuoles. Indeed, Ordenes et al. [52] identified thapsigargin-sensitive Ca2+ pump activity present in the Golgi apparatus vesicles isolated from the elongation zone of etiolated pea epicotyl.
- (3)
- Nevertheless, imaging of fluorescent thapsigargin-stained egg cells at which the fluorophore was added at a concentration of 10 µM failed to reveal any thapsigargin-binding sites other than those localized in the ER membranes visualized by injecting DiI into the isolated wheat egg cells (see Figure 8a–d). Since imaging Dil injected into wheat egg cells proved to be a reliable and effective method in visualizing specifically the ER membranes in the wheat female gamete [36], this observation argues in favor of our hypothesis. Additionally, Pònya et al. [36] identified the ER by CTC (chlortetracycline) labelling to be the main calcium store in the wheat egg cell. Thus, it seems unlikely that calcium leaking from the ER into the cytoplasm upon the addition of thapsigargin could be sequestered into other cell organelles.
- (4)
- The observation that 10 µM thapsigargin treatment caused Ca2+ release in unfertilized egg cells incubated in IVF medium without or with calcium and that in the latter case, the [Ca2+]cyt transient was not significantly higher compared to that measured when cells were incubated in calcium-free medium (see Figure 6c and Figure 7a) suggests that thapsigargin at this concentration has little effect on the plasma membrane ATPase; otherwise the [Ca2+]cyt rise would have been much higher when extracellular calcium was present in the incubation medium due to the cell’s “succumbing” to the tremendous (20,000-fold: 0.1 µM intracellular versus 2 mM extracellular Ca2+ concentration) “Ca2+ pressure” on the cell membrane. In concert with this assumption, when applied to unfertilized egg cells incubated in IVF medium containing 2 mM CaCl2, thapsigargin at a high concentration (100 µM) caused a rapidly rising increase in [Ca2+]cyt, the peak value of which was higher than that observed in wheat egg cells treated with thapsigargin at 10 µM (compare Figure 7a,b). The plateau reached in [Ca2+]cyt was sustained during [Ca2+]cyt measurement (n = 17) and only slightly diminished due to photobleaching of the calcium-sensitive dye. This finding lends credit to the hypothesis that at this concentration, Ca2+ overload occurs in the cell, most probably due to the inhibitory effect exerted by thapsigargin on the plasma membrane calcium pumps.
4. Experimental Section
4.1. Plant Materials
4.2. Gamete Isolation
4.3. Microinjection of Live Egg Cells and Visualizing the Fluorophores
4.4. The IVF Procedure
4.5. Cell Wall Detection
4.6. Measurement of Fura-2 Dextran Fluorescence
4.7. Image Recording and Processing
4.8. Procedure of 1,1'-Dihexadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate (DiI) Injection
4.9. Thapsigargin Treatment
4.10. Visualization of Thapsigargin-Binding Sites in the Wheat Egg
4.11. Ca2+ Influx Inhibition with Gadolinium
4.12. Culture Procedures
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Steinhardt, R.; Zucker, R.; Schatten, G. Intracellular calcium release at fertilization in the sea urchin egg. Dev. Biol. 1977, 58, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Steinhardt, R.; Zucker, R.; Schatten, G. Intracellular calcium release at fertilization in the ascidian egg. Dev. Biol. 1977, 135, 182–190. [Google Scholar]
- Speksnijder, J.E.; Corson, D.W.; Sardet, C. Free calcium pulses following fertilization in the ascidian egg. Dev. Biol. 1989, 135, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, M.J.; Patel, R. Calcium and cell cycle control. Development 1990, 108, 525–542. [Google Scholar] [PubMed]
- Kline, D.; Kline, J.T. Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse egg. Dev. Biol. 1992, 149, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Stricker, S.A. Comparative biology of calcium signaling during fertilization and egg activation in animals. Dev. Biol. 1999, 211, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, M.J.; Steinhardt, R.A. Ionic regulation of egg activation. Q. Rev. Biophys. 1982, 15, 593–666. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, L.F. Sources of calcium in egg activation: A review and hypothesis. Dev. Biol. 1983, 99, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Nucitelli, R. How do sperm activate eggs? Curr. Top. Dev. Biol. 1991, 25, 1–16. [Google Scholar] [PubMed]
- Swann, K.; Ozil, J.P. Dynamics of the calcium signal that triggers mammalian egg activation. Int. Rev. Cytol. 1994, 152, 183–222. [Google Scholar] [PubMed]
- Eisen, A.; Reynolds, G.T. Source and sinks for the calcium released during fertilization of single sea urchin eggs. J. Cell Biol. 1985, 127, 641–652. [Google Scholar]
- Jaffe, L.F. The role of calcium explosions, waves, and pulses in activating eggs. In Biology of Fertilization; Metz, C.B., Monroy, A., Eds.; Academic Press: Orlando, FL, USA, 1985; Volume 3, pp. 12–165. [Google Scholar]
- Miyazaki, S. Inositol 1,4,5-trisphosphate-induced calcium release and guanine nucleotide-binding protein-mediated periodic calcium rises in golden hamster eggs. J. Cell Biol. 1988, 106, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Kado, R.T.; Jaffe, L.A. Development of calcium release mechanisms during starfish oocyte maturation. Dev. Biol. 1990, 140, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Nakada, K.; Shirakawa, H.; Miyazaki, S. Development of inositol trisphosphate-induced calcium release mechanism during maturation of hamster oocytes. Dev. Biol. 1993, 156, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Mehlmann, L.M.; Kline, D. Regulation of intracellular calcium in the mouse egg: Calcium release in response to sperm or inositol trisphosphate is enhanced after meiotic maturation. Biol. Reprod. 1994, 51, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Swann, K.; Whitaker, M. The part played by inositol triphosphate and calcium in the propagation of the fertilization wave in sea urchin eggs. J. Cell Biol. 1986, 103, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Stricker, S.A.; Centonze, V.E.; Paddock, S.W.; Schatten, G. Confocal microscopy of fertilization-induced calcium dynamics in sea urchin eggs. Dev. Biol. 1992, 149, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Gillot, I.; Whitaker, M. Imaging calcium waves in eggs and embryos. J. Exp. Biol. 1993, 184, 213–219. [Google Scholar]
- Mohri, T.; Ivonnet, P.I.; Chambers, E.L. Effect on sperm-induced activation current and increase of cytosolic Ca2+ by agents that modify the mobilization of [Ca2+]i I. Heparin and pentosan polysulfate. Dev. Biol. 1995, 172, 139–157. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.S. Mechanisms of calcium regulation in sea urchin eggs and their activities during fertilization. Curr. Top. Dev. Biol. 1995, 30, 63–101. [Google Scholar] [PubMed]
- Whitaker, M.; Swann, K. Lighting the fuse at fertilization. Development 1993, 117, 1–12. [Google Scholar]
- Antoine, A.F.; Dumas, C.; Faure, J.E.; Feijò, J.A.; Rougier, M. Egg activation in flowering plants. Sex. Plan. Reprod. 2001, 14, 21–26. [Google Scholar] [CrossRef]
- Russell, S.D. Double fertilization. Int. Rev. Cytol. 1992, 140, 357–388. [Google Scholar]
- Kranz, E.; Dresselhaus, T. In vitro fertilization with isolated higher plant gametes. Trends Plant Sci. 1996, 1, 82–89. [Google Scholar] [CrossRef]
- Digonnet, C.; Aldon, D.; Leduc, N.; Dumas, C.; Rougier, M. First evidence of a calcium transient in flowering plants at fertilization. Development 1997, 124, 2867–2874. [Google Scholar] [PubMed]
- Okamoto, T.; Higuchi, K.; Shinkawa, T.; Isobe, T.; Lörz, H.; Koshiba, T.; Kranz, E. Identification of major proteins in maize egg cells. Plant Cell Physiol. 2004, 10, 1406–1412. [Google Scholar] [CrossRef]
- Roberts, S.K.; Gillot, I.; Brownlee, C. Cytoplasmic calcium and Fucus egg activation. Development 1994, 120, 155–163. [Google Scholar]
- Roberts, S.K.; Brownlee, C. Calcium influx, fertilization potential and egg activation in Fucus serratus. Zygote 1995, 3, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Antoine, A.F.; Faure, J.E.; Cordeiro, S.; Dumas, C.; Rougier, M.; Feijò, J.A. A calcium influx is triggered and propagates in the zygote as a wave front during in vitro fertilization of flowering plants. Proc. Natl. Acad. Sci. USA 2000, 97, 10643–10648. [Google Scholar] [CrossRef] [PubMed]
- Weterings, K.; Russell, S.D. Experimental analysis of the fertilization process. Plant Cell 2004, 16, S107–S118. [Google Scholar] [CrossRef] [PubMed]
- Pónya, Z.; Pv Fehér, F.; Av Mitykó, J.; Dudits, D.; Barnabás, B. Optimisation of introducing foreign genes into egg cells and zygotes of wheat (Tritium aestivum L.) via microinjection. Protoplasma 1999, 208, 163–172. [Google Scholar] [CrossRef]
- Kranz, E.; Bautor, J.; Lörz, H. In vitro fertilization of single, isolated gametes of maize mediated by electrofusion. Sex. Plant Reprod. 1991, 4, 12–16. [Google Scholar]
- Kranz, E.; Lörz, H. In vitro fertilization of maize by single egg and sperm cell protoplast fusion mediated by high calcium and high pH. Zygote 1994, 2, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Pónya, Z.; Tímár, I.; Szabó, L.; Kristóf, Z.; Barnabás, B. Morphological characterisation of wheat (T. aestivum L.) egg cell protoplasts isolated from immature and overaged caryopses. Sex. Plant Reprod. 1999, 11, 357–359. [Google Scholar] [CrossRef]
- Pónya, Z.; Kristòf, Z.; Ciampolin, F.; Faleri, C.; Cresti, M. Structural change in the endoplasmic reticulum during the in situ development and in vitro fertilization of wheat egg cells. Sex. Plant Reprod. 2004, 17, 177–188. [Google Scholar] [CrossRef]
- Thastrup, O.; Cullen, P.J.; Bjørn, K.; Hanley, M.R.; Dawson, A.P. Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2+-ATPase. Proc. Natl. Acad. Sci. USA 1990, 87, 2466–2470. [Google Scholar] [CrossRef] [PubMed]
- Sagara, Y.; Inesi, G. Inhibition of the sarcoplasmic reticulum Ca2+ transport ATPase by thapsigargin at subnanomolar concentrations. J. Biol. Chem. 1991, 266, 13503–13506. [Google Scholar] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of calcium indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar]
- Vissenberg, K.; Feijó, J.A.; Weisenseel, M.H.; Verbelen, J.P. Ion fluxes, auxin and the induction of elongation growth in Nicotiana tabacum cells. J. Exp. Bot. 2001, 52, 2161–2167. [Google Scholar] [PubMed]
- Antoine, A.F.; Faure, J.E.; Dumas, C.; Feijó, J.A. Differential contribution of cytoplasmic Ca2+ and Ca2+ influx to gamete fusion and egg activation in maize. Nat. Cell Biol. 2001, 3, 1120–1123. [Google Scholar] [CrossRef] [PubMed]
- Treiman, M.; Caspersen, C.; Christensen, S.B. A tool coming of age: thapsigargin as an inhibitor of sarco-endoplasmic reticulum Ca2+-ATPases. Trends Pharmacol. Sci. 1998, 19, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Thomson, L.J.; Hall, J.L.; Williams, L.E. A study of the effect of inhibitors of the animal sarcoplasmic/endoplasmic reticulum-type calcium pumps on the primary Ca2+-ATPase of red beet. Plant Physiol. 1994, 104, 1295–1300. [Google Scholar] [PubMed]
- Zhao, J.; Yu, F.L.; Lianf, S.P.; Zhou, C.; Yang, H.Y. Changes of calcium distribution in egg cells, zygotes and two-celled proembryos of rice (Oryza sativa L.). Sex. Plant Reprod. 2002, 14, 331–337. [Google Scholar] [CrossRef]
- Mòl, R.; Matthys-Rochon, E.; Dumas, C. The kinetics of cytological events during double fertilization in Zea mays L. Plant J. 1994, 5, 197–206. [Google Scholar] [CrossRef]
- Meyer, T.; Stryer, I. Molecular model for receptor-stimulated calcium spiking. Proc. Natl. Acad. Sci. USA 1988, 85, 5051–5055. [Google Scholar] [CrossRef] [PubMed]
- Parker, I.; Ivorra, I. Inhibition by Ca2+ of inositol trisphosphate-mediated Ca2+ liberation: A possible mechanism of oscillatory release of [Ca2+]i. Proc. Nat. Acad. Sci. USA 1990, 87, 160–264. [Google Scholar]
- Faure, J.E.; Digonnet, C.; Dumas, C. An in vitro system for adhesion and fusion of maize gametes. Science 1994, 263, 598–1600. [Google Scholar] [CrossRef]
- Jackson, T.R.; Patterson, S.I.; Thastrup, O.; Hanley, M.R. A novel tumour promoter, thapsigargin, transiently increases cytoplasmic free Ca2+ without generation of inositol phosphates in NG115-401L neuronal cells. Biochem. J. 1988, 253, 81–86. [Google Scholar] [PubMed]
- Gouy, H.; Cefai, D.; Christensen, S.B.; Debré, P.; Bismuth, G. Ca2+ influx in human T lymphocytes is induced independently of inositol phosphate production by mobilization of intracellular Ca2+ stores. A study with the Ca2+ endoplasmic reticulum-ATPase inhibitor thapsigargin. Eur. J. Immunol. 1990, 20, 2269–2275. [Google Scholar] [CrossRef] [PubMed]
- Wictome, L.E.; Henderson, I.; Lee, A.G.; East, J.M. Mechanisms of inhibition of the calcium pump of sarcoplasmic reticulum by thapsigargin. Biochem. J. 1992, 283, 525–529. [Google Scholar] [PubMed]
- Ordenes, V.R.; Reyes, C.; Wolff, D.; Orellana, A. A thapsigargin-sensitive Ca2+ pump is present in the pea Golgi apparatus membrane. Plant Physiol. 2002, 129, 1820–1828. [Google Scholar] [CrossRef] [PubMed]
- Chaubal, R.; Reger, B.J. The dynamics of calcium distribution in the synergid cells of wheat after pollination. Sex. Plant Reprod. 1992, 5, 206–213. [Google Scholar]
- Heslop-Harrison, J.; Heslop-Harrison, Y. Evaluation of pollen viability by enzymatically induced fluorescence; intracellular hydrolysis of fluorescein diacetate. Stain Technol. 1970, 45, 115–120. [Google Scholar] [PubMed]
- Poenie, M. Alteration of intracellular fura-2 fluorescence by viscosity: A simple correction. Cell Calcium 1990, 11, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Kao, K.N.; Michayluk, M.R. Nutritional requirements for growth of Vicia hajastana cells and protoplasts at a very low population density in liquid media. Planta 1975, 126, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Parrington, J.; Swann, K.; Shevchenko, V.I.; Sesay, A.K.; Lai, F.A. Calcium oscillations in mammalian eggs triggered by a soluble sperm protein. Nature 1996, 379, 354–368. [Google Scholar] [CrossRef]
- Rudd, J.J.; Franklin-Tong, V.E. Unravelling response-specificity in Ca2+ signalling pathways in plant cells. New Phytol. 2001, 151, 7–33. [Google Scholar] [CrossRef]
- Peng, X.B.; Sun, M.X.; Yang, H.Y. Comparative detection of calcium fluctuations in single female sex cells of tobacco to distinguish calcium signals triggered by in vitro fertilization. J. Integr. Plant Biol. 2009, 51, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.L.; Tian, H.Q.; Russell, S.D. Calcium function and distribution during fertilization in angiosperms. Am. J. Bot. 2007, 94, 1046–1060. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, Y.; Nishimaki, M.; Takeuchi, H.; Geitmann, A.; Kurihara, D.; Higashiyama, T. Live imaging of calcium spikes during double fertilization in Arabidopsis. Nat. Commun. 2014, 5, 1–9. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pónya, Z.; Corsi, I.; Hoffmann, R.; Kovács, M.; Dobosy, A.; Kovács, A.Z.; Cresti, M.; Barnabás, B. When Isolated at Full Receptivity, in Vitro Fertilized Wheat (Triticum aestivum, L.) Egg Cells Reveal [Ca2+]cyt Oscillation of Intracellular Origin. Int. J. Mol. Sci. 2014, 15, 23766-23791. https://doi.org/10.3390/ijms151223766
Pónya Z, Corsi I, Hoffmann R, Kovács M, Dobosy A, Kovács AZ, Cresti M, Barnabás B. When Isolated at Full Receptivity, in Vitro Fertilized Wheat (Triticum aestivum, L.) Egg Cells Reveal [Ca2+]cyt Oscillation of Intracellular Origin. International Journal of Molecular Sciences. 2014; 15(12):23766-23791. https://doi.org/10.3390/ijms151223766
Chicago/Turabian StylePónya, Zsolt, Ilaria Corsi, Richárd Hoffmann, Melinda Kovács, Anikó Dobosy, Attila Zoltán Kovács, Mauro Cresti, and Beáta Barnabás. 2014. "When Isolated at Full Receptivity, in Vitro Fertilized Wheat (Triticum aestivum, L.) Egg Cells Reveal [Ca2+]cyt Oscillation of Intracellular Origin" International Journal of Molecular Sciences 15, no. 12: 23766-23791. https://doi.org/10.3390/ijms151223766
APA StylePónya, Z., Corsi, I., Hoffmann, R., Kovács, M., Dobosy, A., Kovács, A. Z., Cresti, M., & Barnabás, B. (2014). When Isolated at Full Receptivity, in Vitro Fertilized Wheat (Triticum aestivum, L.) Egg Cells Reveal [Ca2+]cyt Oscillation of Intracellular Origin. International Journal of Molecular Sciences, 15(12), 23766-23791. https://doi.org/10.3390/ijms151223766