Hypoxia Enhances Protective Effect of Placental-Derived Mesenchymal Stem Cells on Damaged Intestinal Epithelial Cells by Promoting Secretion of Insulin-Like Growth Factor-1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
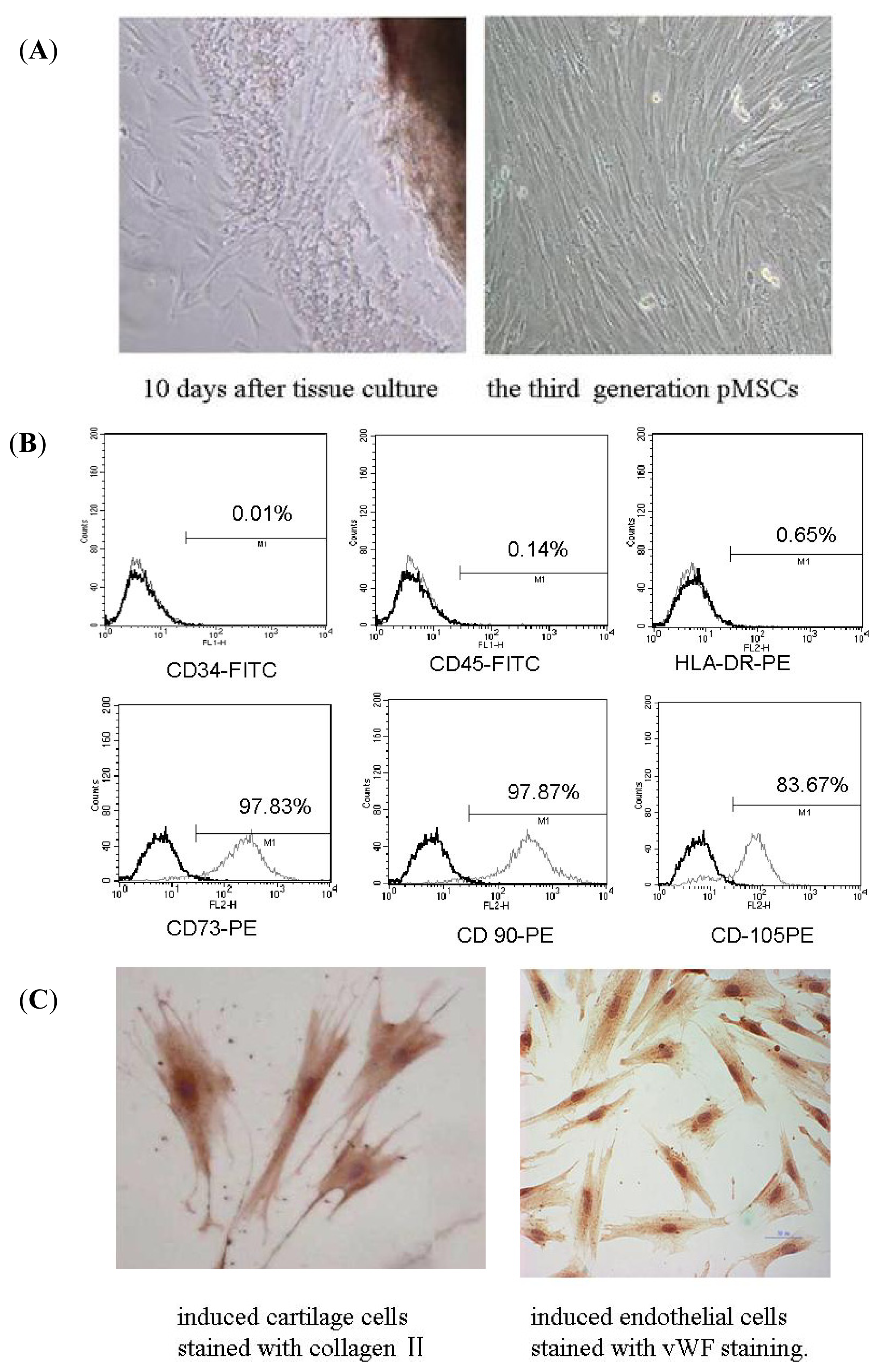
2.1. Isolation and Identification of pMSCs
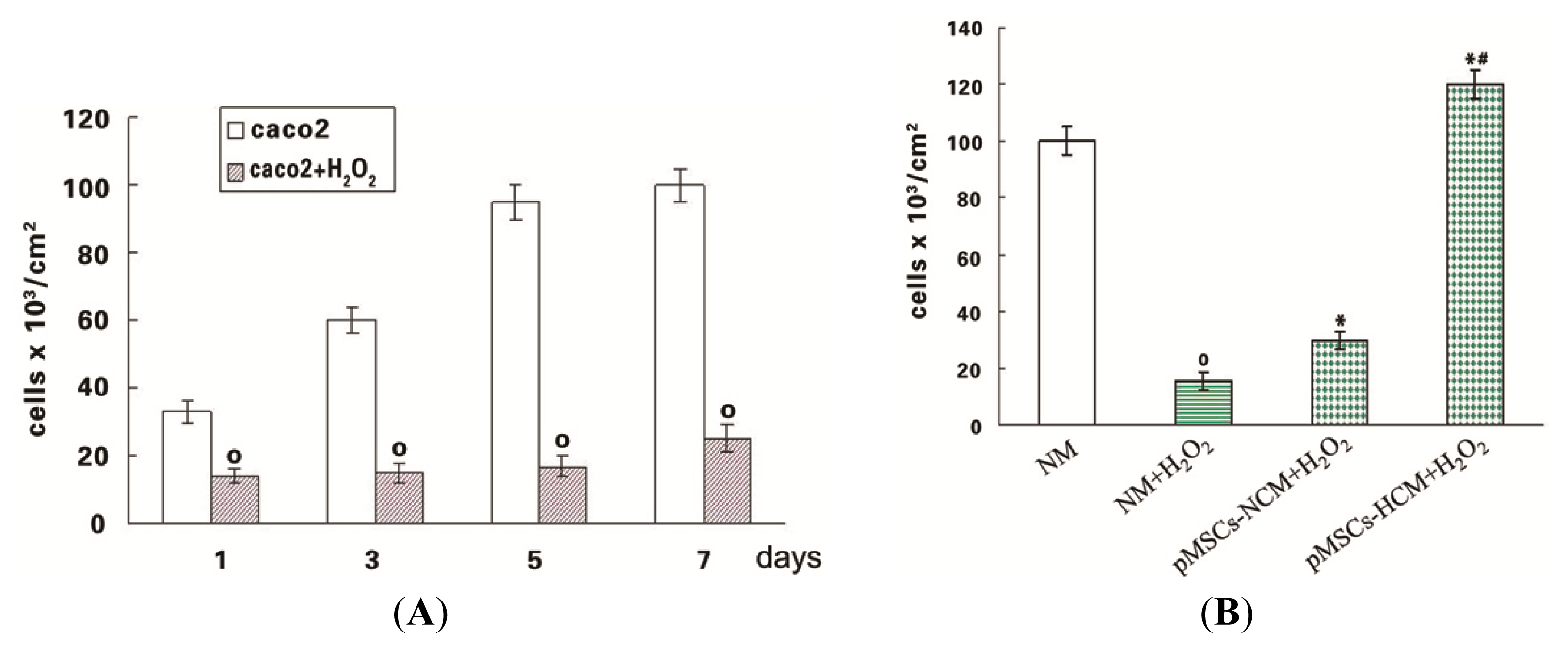
2.2. The pMSCs Hypoxia Culture Medium Is Better Protection for H2O2-treated-caco2 than pMSCs Normoxia Culture Medium
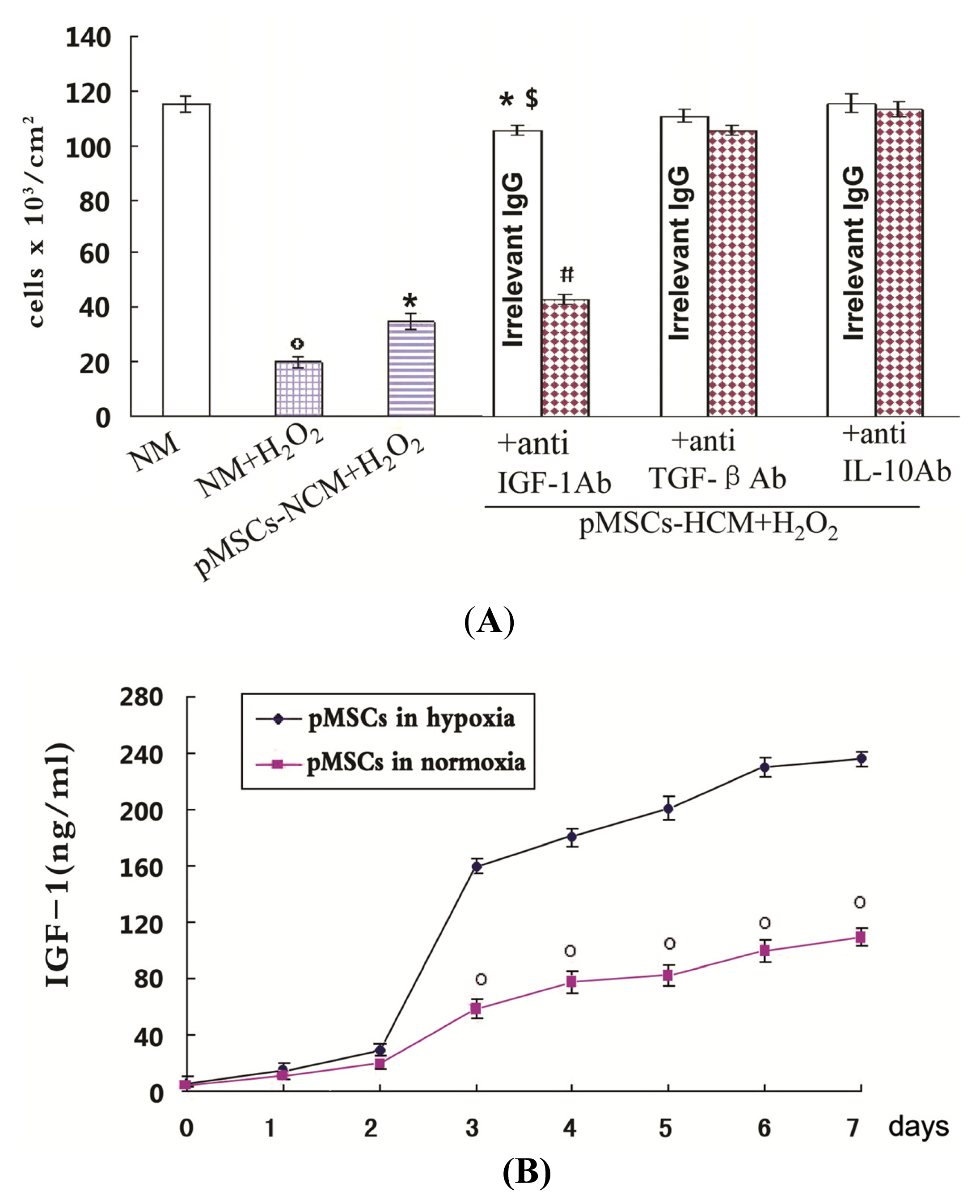
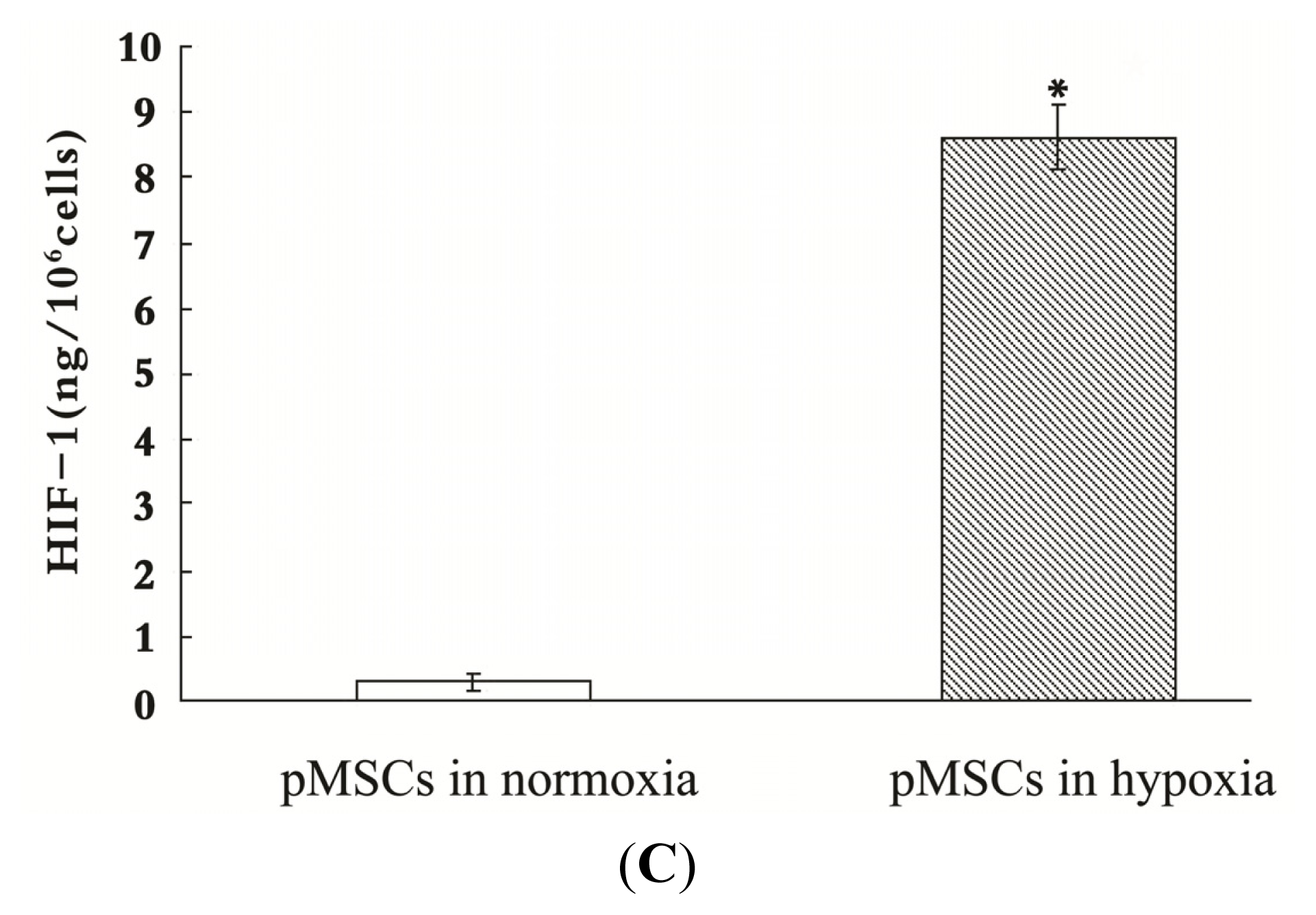
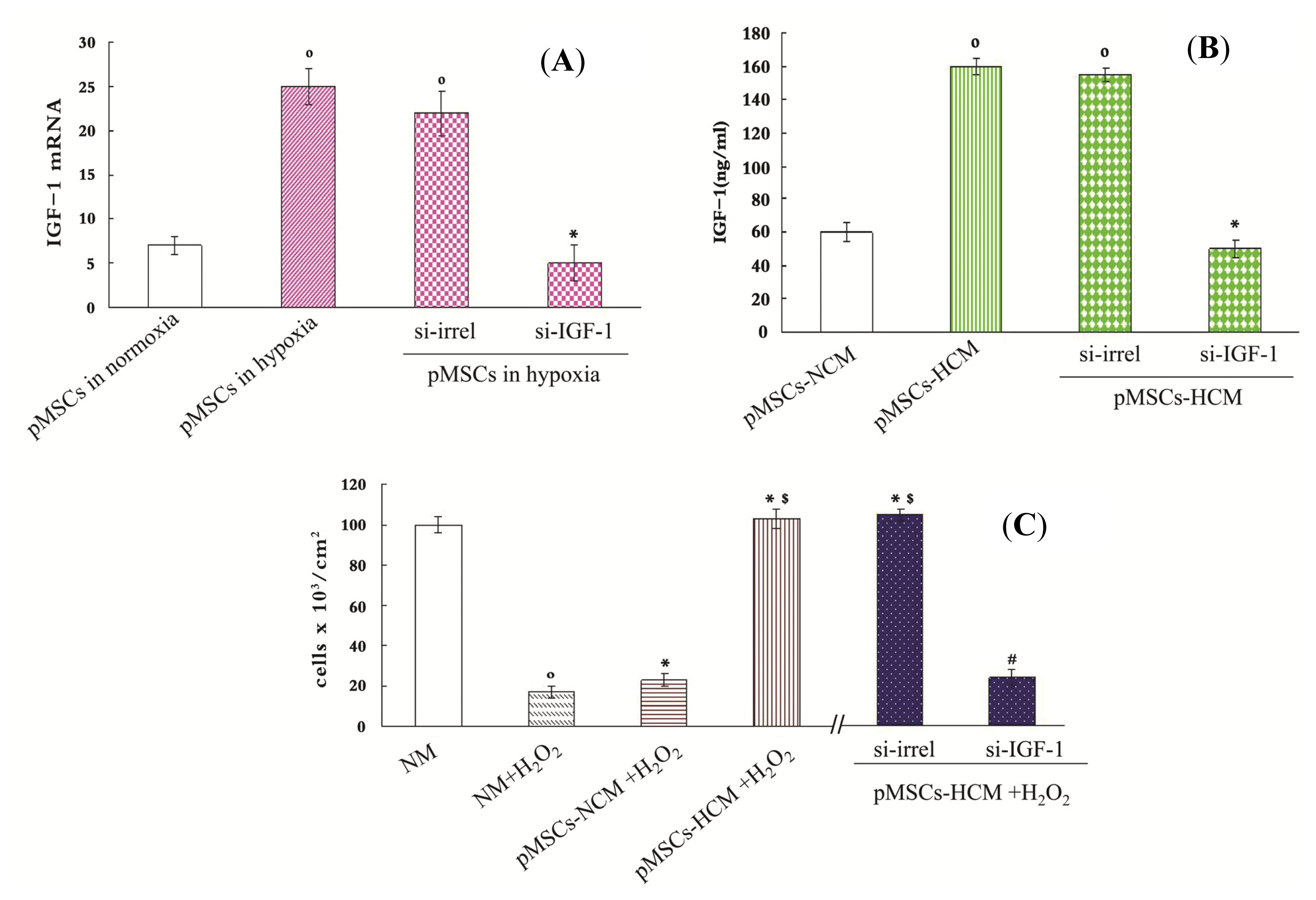
2.3. pMSCs Hypoxia Culture Medium Produces Enhanced Protection for H2O2-treated-caco2 Cells through Increased IGF-1
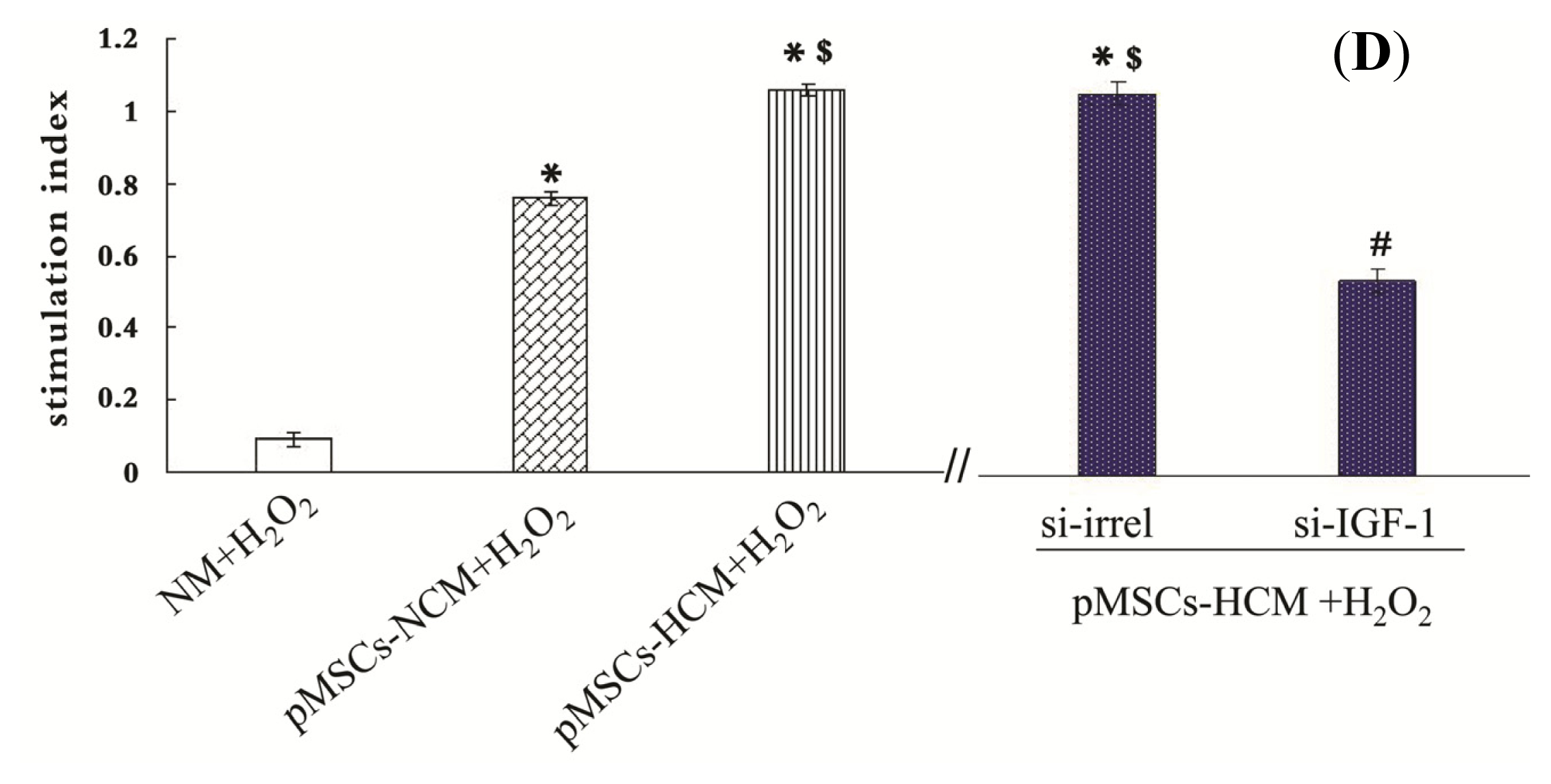
2.4. pMSCs Hypoxia Culture Medium Is More Conducive to Promotion of Proliferation of H2O2-treated-caco2 Cells than pMSCs Normoxia Culture Medium via IGF-1
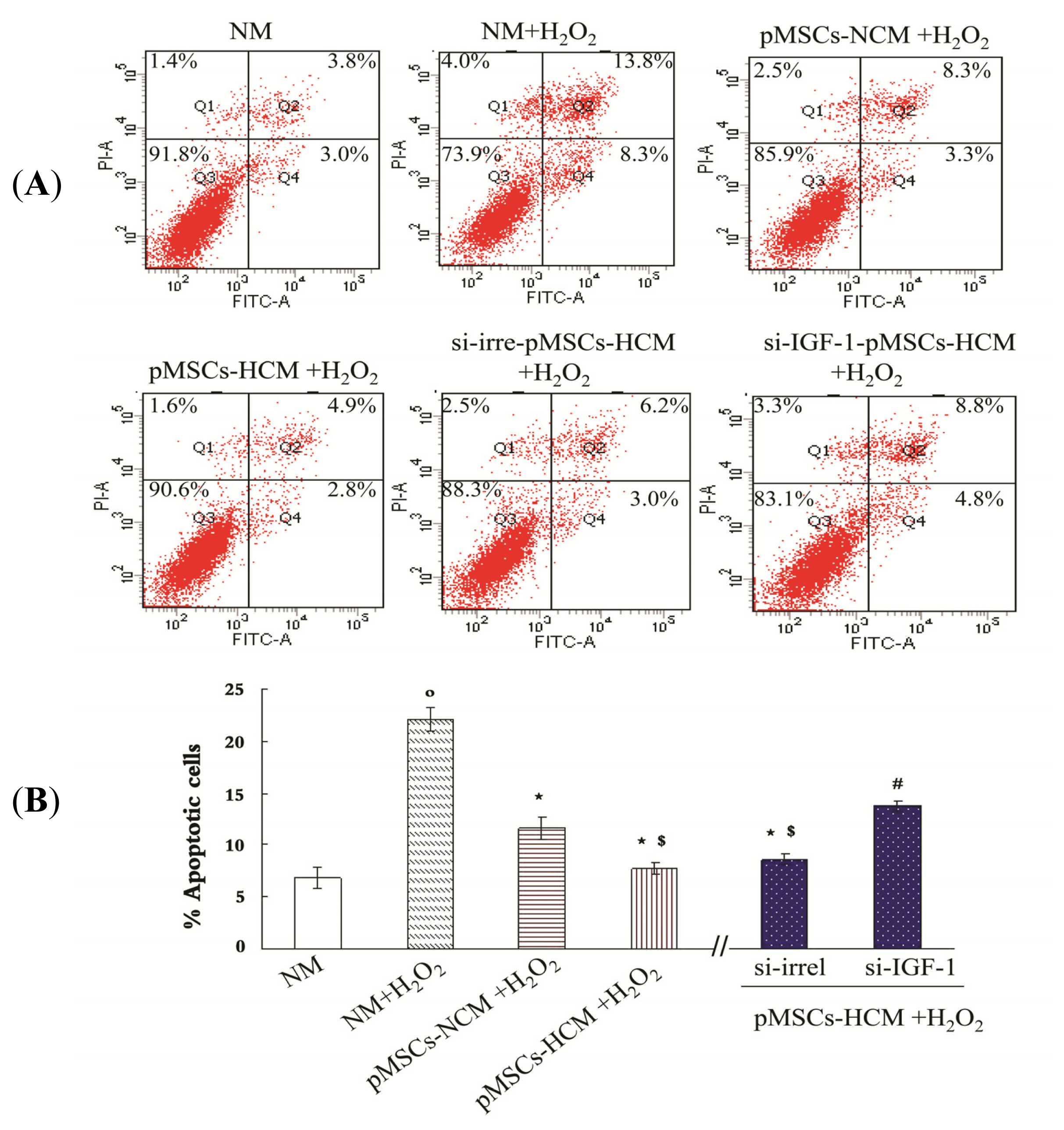
2.5. pMSCs Hypoxia Culture Medium Has Greater Capacity of Limiting H2O2-treated-caco2 Apoptosis via Increased IGF-1 Compared to pMSCs Normoxia Culture Medium
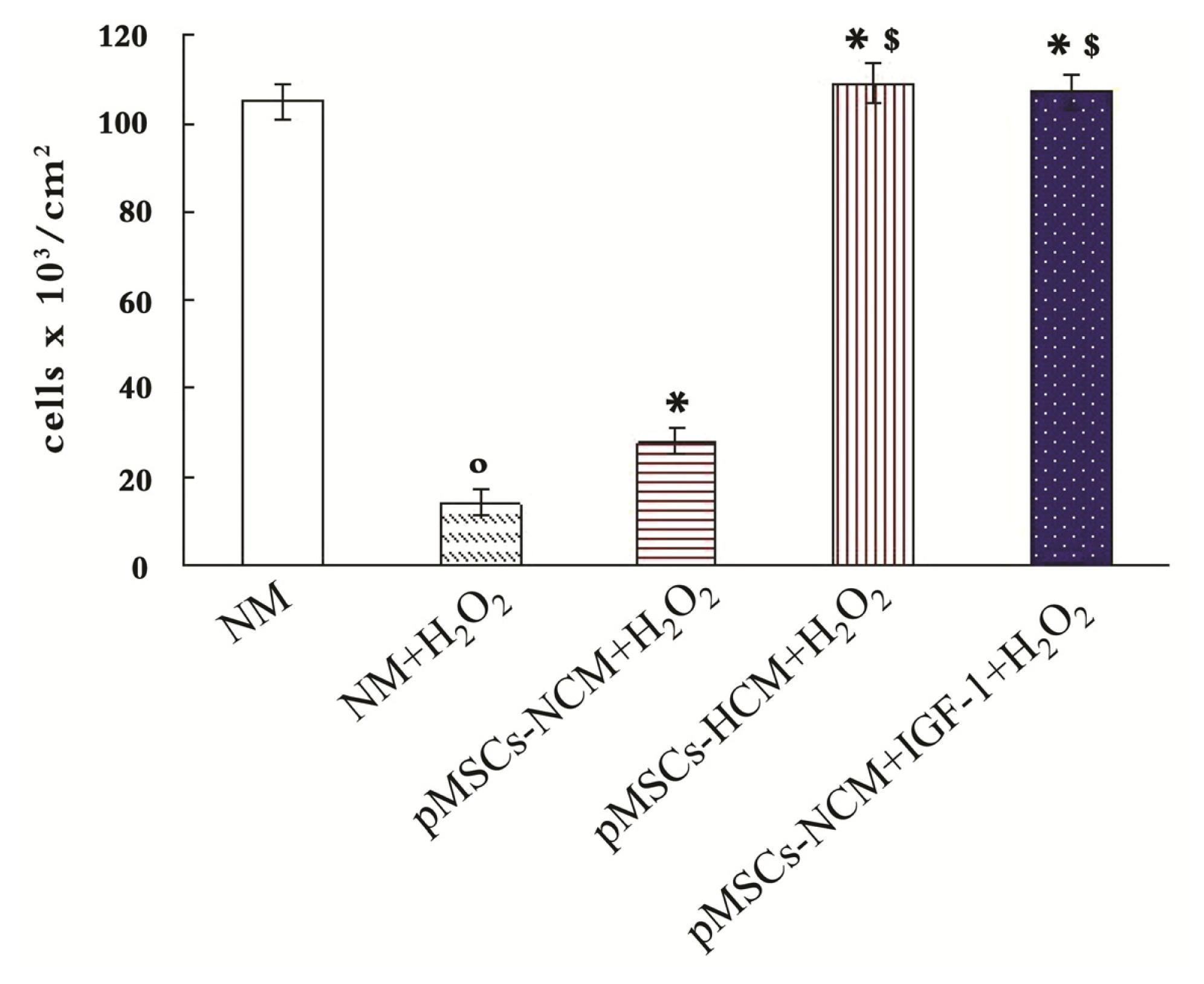
2.6. Addition of IGF-1 Enhances the Protective Effect of pMSCs-NCM on H2O2-treated Caco2 Cells
2.7. Discussion
3. Experimental Section
3.1. Isolation and Passage of Placenta-Derived Mesenchymal Stem Cells
3.2. Trypan Blue Staining
3.3. Cartilage Cells Differentiation and Collagen II Staining
3.4. Endothelial Cells Differentiation and von Willebrand Factor Staining
3.5. Flow Cytometry to Detect Cell Surface Markers and Apoptosis
3.6. ELISA
3.7. RNA Interference
3.8. RT-PCR and Quantitative Real-Time RT-PCR
3.9. Adding the Blocking Antibodies for IGF-1, TGFβ and IL-10
3.10. MTT Determination of Cell Viability
3.11. Cell Immunofluorescence
3.12. Hypoxia Inducible Factor-1 Activity Assay
3.13. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-15-01983-s001.pdfAcknowledgments
Conflicts of Interest
References
- Oldeburg, W.A.; Lau, L.L.; Rodenberg, T.J.; Edmonds, H.J.; Burger, C.D. Acute mesenteric ischemia: A clinical review. Arch. Intern. Med 2004, 164, 1054–1062. [Google Scholar]
- Abu-Elmagd, K.; Reyes, J.; Bond, G.; Mazariegos, G.; Wu, T.; Martin, N.; Sindhi, R. Clinical intestinal transplantation: A decade of experience at a single center. Ann. Surg 2001, 234, 416–417. [Google Scholar]
- Quaedackers, J.S.; Beukm, R.J.; Bennetm, L.; Charlton, A.; Oude Eqbrink, M.G.; Gunn, A.J.; Heineman, E. An evaluation of methods for grading histologic injury following ischemia/reperfusion of the small bowel. Transplant. Proc 2000, 32, 1307–1310. [Google Scholar]
- Khanna, A.; Rossman, J.; Caty, M.G.; Fung, H.L. Beneficial effects of intraluminal nitroglycerin in intestinal ischemia-reperfusion injury in rats. J. Surg. Res 2003, 114, 5–24. [Google Scholar]
- Wang, H.J.; Huang, J. Basic and applicative studies of acidic fibroblast growth factor. Anat. Res 2005, 2, 308–311. [Google Scholar]
- Imberti, B.; Morigi, M.; Tomasoni, S.; Rota, C.; Corna, D.; Longaretti, L. Insulin-like growth factor-1 sustains stem cell mediated renal repair. J. Am. Soc. Nephrol 2007, 18, 2921–2928. [Google Scholar]
- Morigi, M.; Introna, M.; Imberti, B.; Corna, D.; Abbate, M.; Rata, C. Human bone marrow mesenchymal stem cells accelerate recovery of acute renal injury and prolong survival in mice. Stem Cells 2008, 26, 2075–2082. [Google Scholar]
- Xu, Y.X.; Chen, L.; Wang, R. Mesenchymal stem cell therapy for diabetes through paracrine mechanisms. Med. Hypotheses 2008, 71, 390–393. [Google Scholar]
- Zhang, Y.; Li, C.; Jiang, X.; Zhang, S.; Wu, X.; Liu, B.; Tang, P. Human placenta-derived mesenchymal progenitor cells support culture expansion of long-term culture-initiating cells from cord blood CD34+ cells. Exp. Hematol 2004, 32, 657–664. [Google Scholar]
- Fukuchi, Y.; Nakajima, H.; Sugiyama, D.; Hirose, I.; Kitamura, T.; Tsuji, K. Human placenta-derived cells have mesenchyma stem/progenitor cell potential. Stem Cells 2004, 22, 649–658. [Google Scholar]
- Yen, B.L.; Huang, H.I.; Chien, C.C. Isolation of multipotent cells from human term placenta. Stem Cells 2005, 23, 3–9. [Google Scholar]
- Intanker, P.S.; Scherjon, S.A.; Kleijburg-van der-Keur, C. Isolation of mesenchymal stem cells of fetal or maternal origin from human placenta. Stem Cell 2004, 22, 1338–1345. [Google Scholar]
- Manuelpillai, U.; Tchongue, J.; Lourensz, D.; Vaghjini, V.; Samuel, C.S.; Liu, A.; Williams, E.D.; Sievert, W. Transplantation of human amnion epithelial cells reduces hepatic fibrosis in immunocompetent CCl4-treated mice. Cell Transplant 2010, 19, 1157–1168. [Google Scholar]
- Cargnoni, A.; Gibelli, L.; Tosini, A.; Signoroni, P.B.; Nassuato, C.; Arienti, D. Translantation of allogeneic and xenogeneic placenta-derived cells reduces bleomycin-induced lung fibrosis. Cell Transplant 2009, 18, 405–422. [Google Scholar]
- Pan, H.C.; Yang, D.Y.; Chiu, Y.T.; Lai, S.Z.; Wang, Y.C.; Chang, M.H. Enhanced regeneration in injured sciatic nerve by human amniotic mesenchymal stem cell. J. Clin. Neurosci 2006, 13, 570–575. [Google Scholar]
- Minaqawa, T.; Imamura, T.; Iqawa, Y. Differentiation of smooth muscle cells from human amniotic mesenchymal cells implanted in the freeze-injured mouse urinary bladder. Eur. Urol 2010, 58, 299–306. [Google Scholar]
- Wei, J.P.; Nawata, M.; Wakitani, S. Human amniotic mesenchymal cells differentiate into chondrocyte. Cloning Stem Cells 2009, 11, 19–26. [Google Scholar]
- Tsuji, H.; Miyoshi, S.; Ikegami, Y.; Hida, N.; Asada, H.; Togashi, I.; Suzuki, J. Xenografted human amniotic membrane-derived mesenchymal stem cells are immunologically tolerated and transdifferentiated into cardiomyocyte. Circ. Res 2010, 106, 1613–1623. [Google Scholar]
- Kawamichi, Y.; Cui, C.H.; Toyoda, M. Cells of extraembroonic mesodermal origin confer human dystrophin in the mdx model of Duchenne muscular dystrophy. J. Cell. Physiol 2010, 223, 695–702. [Google Scholar]
- Paradowska, E.; Blach-Olszewska, Z.; Gejdel, E. Constitutive and induced cytokine production by human placenta and amniotic membrane at term. Placenta 1997, 18, 441–446. [Google Scholar]
- Liu, D.Q.; Wang, F.C.; Dong, S.W. Experimental study on the implantation and differentiation of donors BMDSCs in the intestinal epithelium of bone marrow-chimeric mice. Chin. J. Gastroenterol. Hepatol 2008, 17, 635–638. [Google Scholar]
- Han, Q.; He, D.N.; Liu, Y.N. Repair of intestinal irradiation injury by Flk-1 + MSCs and SP cells. Jichu Yixue Yu Linchuang 2008, 28, 563–569. [Google Scholar]
- Williams, K.I.; Fuller, C.R.; Fagin, J. Mesenchymal IGF-1 overexpression: paraerine effects in the intestine distinct from endocrine actin. Am. J. Physiol. Gastin. Intest. Liver Physiol 2002, 283, G875–G885. [Google Scholar]
- Das, R.; Jahr, H.; van Osch, G.; Farrell, E. The role of hypoxia in bone marrow-derived mesenchymal stem cells: consideration for regenerative medicine approaches. Tissue Eng. B Rev 2010, 16, 159–168. [Google Scholar]
- Rosova, I.; Dao, M.; Capoccia, B.; Link, D.; Nolta, A. Hypoxic preconditioning results in increased motility and improved therapeutic potential of human mesenchymal stem cell. Stem Cells 2008, 26, 2173–2182. [Google Scholar]
- Jeremie, J.; Matthieu, B.; Joachim, C.; Jean, C.; Pascale, D.; Zoran, I.; Thierry, C.; Laurent, B. Hypoxia-preconditioned mesenchymal stromal cells improve cardiac function in a swine model of chronic myocardial ischaemia. Eur. J. Cardiothorac. Surg 2013, 43, 1050–1057. [Google Scholar]
- Togel, F.; Hu, Z.; Weiss, K.; Isaac, J.; Lange, C.; Westenfelder, C. Administered mesenchymal stem cells protect against ischemic acute renal failure through differentiation-independent mechanisms. Am. J. Physiol. Renal Physiol 2005, 289, F31–F42. [Google Scholar]
- Corthesy-Theulaz, I.; Porta, N.; Prinqault, E. Adhesion of Helicobacter pylori to polarized T84 human intestinal cell monolayers is pH dependent. Infect. Immun 1996, 64, 3827–3832. [Google Scholar]
- Yu, X.; Lu, C.; Liu, H.; Rao, S.; Cai, S.; Liu, S.; Kriegel, A.J. Hypoxic preconditioning with cobalt of bone marrow mesenchymal stem cells improves cell migration and enhances therapy for treatment of ischemic acute kidney injury. PLoS One 2013, 8, e62703. [Google Scholar]
- Liu, S.H.; Huang, J.P.; Lee, R.K. Paracrine factors from human placental multipotent mesenchymal stromal cells protect endothelium from oxidative injury via STAT3 and manganese superoxide dismutase activation. Biol. Reprod 2010, 82, 905–913. [Google Scholar]
- Iliodromiti, Z.; Antonakopoulos, N.; Sifakis, S.; Tsikouras, P.; Daniilidis, A.; Dafopoulos, K. Endocrine, paracrine, and autocrine placental mediators in labor. Hormones 2012, 11, 397–409. [Google Scholar]
- Kong, P.; Xie, X.; Li, F. Placenta mesenchymal stem cell accelerates wound healing by enhancing angiogenesis in diabetic Goto-Kakizaki (GK) rats. Biochem. Biophys. Res. Commun 2013, 438, 410–419. [Google Scholar]
- Ding, H.; Kopple, J.D.; Cohen, A. Recombinant human insulin-like growth factor-accelerates recovery and reduces catabolism in rats with ischemic acute renal failure. J. Clin. Invest 1993, 91, 2281–2287. [Google Scholar]
- Hirschberg, R.; Ding, H. Mechanisms of insulin-like growth factor-1-induced accelerated recovery in experimental ischemic acute renal failure. Miner. Electrolyte Metab 1998, 24, 211–219. [Google Scholar]
- Moses, H.L.; Sena, R. Refulation of differentiation by TGF-β. Curr. Opin. Genet. Dev 1996, 6, 581–586. [Google Scholar]
- Deng, J.; Kohda, Y.; Chiao, H. Interleukin-10 inhiits ischemic and cisplatin-induced acute renal injury. Kidney Int 2001, 60, 2118–2128. [Google Scholar]
- Chen, K.; Okuma, T.; Okamura, K. Insulin-like growth factor-1 prevents gut atrophy and maintains intestinal integrity in septic rats. J. Parenter. Enteral Nutr 1995, 19, 119–124. [Google Scholar]
- Abboud, S.L.; Bethel, C.R.; Aron, D.C. Secretion of insulin-like growth factor 1 and insulin-like growth factor-binding proteins by murine bone marrow stromal cells. J. Clin. Invest 1991, 88, 470–475. [Google Scholar]
- Park, J.H.Y.; Mclusker, R.H.; Vanderhoof, J.A. Secretions of insulin-like growth factor I and IGF-binding protein-II by intestinal epithelial (IEC-6) cell:imlication for autocrine growth regulation. Endocrinology 1992, 131, 1359–1368. [Google Scholar]
- Zhang, X.; Mitsuru, A.; Lgura, K. Mesenchymal progenitor cells derived from chorionic villi of human placenta for cartilage tissue engineering. Biochem. Biophys. Res. Commun 2006, 340, 944–952. [Google Scholar]
- Qian, X.; Ning, H.; Zhang, J.; Hoft, D.F.; Stumpo, D.J.; Blackshear, P.J.; Liu, J. Posttranscriptional regulation of IL-23 expression by IFN-γ through tristetraprolin. J. Immunol 2011, 186, 6454–6464. [Google Scholar]
- Zhang, J.; Qian, X.; Ning, H.; Yang, J.; Xiong, H.; Liu, J. Activation of IL-27 p28 gene transcription by interferon regulatory factor 8 in cooperation with interferon regulatory factor 1. J. Biol. Chem 2010, 285, 21269–21281. [Google Scholar]








© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Du, L.; Yu, Y.; Ma, H.; Lu, X.; Ma, L.; Jin, Y.; Zhang, H. Hypoxia Enhances Protective Effect of Placental-Derived Mesenchymal Stem Cells on Damaged Intestinal Epithelial Cells by Promoting Secretion of Insulin-Like Growth Factor-1. Int. J. Mol. Sci. 2014, 15, 1983-2002. https://doi.org/10.3390/ijms15021983
Du L, Yu Y, Ma H, Lu X, Ma L, Jin Y, Zhang H. Hypoxia Enhances Protective Effect of Placental-Derived Mesenchymal Stem Cells on Damaged Intestinal Epithelial Cells by Promoting Secretion of Insulin-Like Growth Factor-1. International Journal of Molecular Sciences. 2014; 15(2):1983-2002. https://doi.org/10.3390/ijms15021983
Chicago/Turabian StyleDu, Lili, Yanqiu Yu, Haiying Ma, Xiaomei Lu, Ling Ma, Yunan Jin, and Haipeng Zhang. 2014. "Hypoxia Enhances Protective Effect of Placental-Derived Mesenchymal Stem Cells on Damaged Intestinal Epithelial Cells by Promoting Secretion of Insulin-Like Growth Factor-1" International Journal of Molecular Sciences 15, no. 2: 1983-2002. https://doi.org/10.3390/ijms15021983
APA StyleDu, L., Yu, Y., Ma, H., Lu, X., Ma, L., Jin, Y., & Zhang, H. (2014). Hypoxia Enhances Protective Effect of Placental-Derived Mesenchymal Stem Cells on Damaged Intestinal Epithelial Cells by Promoting Secretion of Insulin-Like Growth Factor-1. International Journal of Molecular Sciences, 15(2), 1983-2002. https://doi.org/10.3390/ijms15021983