Transcriptional Analysis of Apoptotic Cerebellar Granule Neurons Following Rescue by Gastric Inhibitory Polypeptide



Abstract
:1. Introduction
2. Results and Discussion
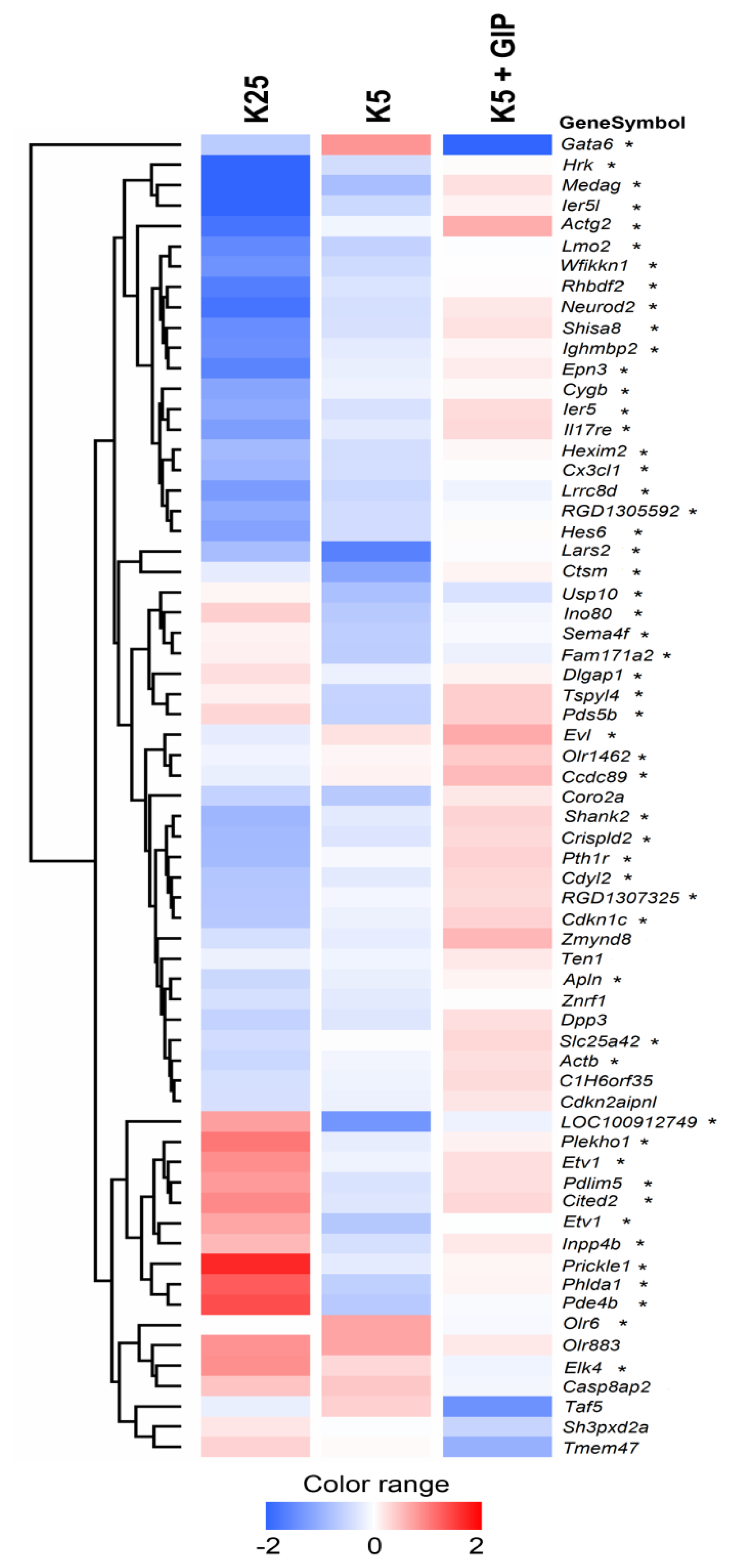
2.1. Whole-Genome Expression Changes Underlying Apoptosis Rescue by Gip
2.2. Deregulated Gene Groups
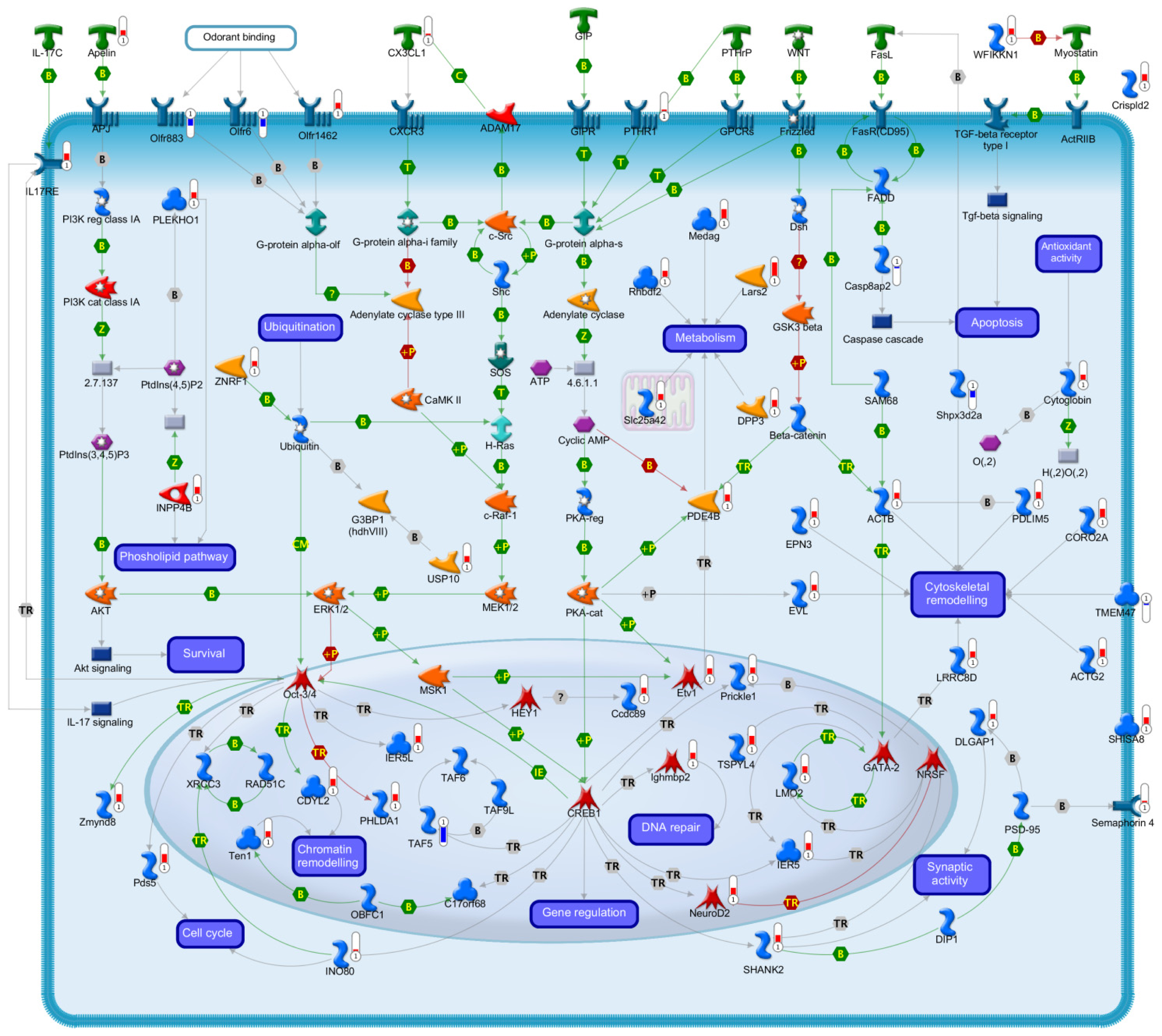
2.2.1. Signal Transduction
2.2.2. Transcription
2.2.3. Cell Cycle
2.2.4. Chromatin Remodelling
2.2.5. Cell Death
2.2.6. Antioxidant Activity
2.2.7. Ubiquitination
2.2.8. Metabolism
2.2.9. Cytoskeletal Organization
3. Experimental Section
3.1. Materials
3.2. Neuronal Cultures
3.3. Microarray Experiments
3.4. Real Time Quantitative PCR
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsB.M. carried out microarray and RT-PCR analysis, and drafted the manuscript; M.T.C. performed all neuronal cultures experiments, P.C. and S.C. conceived and designed the study. S.C. performed microarray data analysis.
References
- Jellinger, K.A. Challenges in neuronal apoptosis. Curr. Alzheimer Res 2006, 3, 377–391. [Google Scholar]
- Arends, M.J.; Wyllie, A.H. Apoptosis: Mechanisms and roles in pathology. Internatl. Rev. Exp. Pathol 1991, 32, 223–254. [Google Scholar]
- Oppenheim, R.W. Cell death during development of the nervous system. Annu. Rev. Neurosci 1991, 14, 453–501. [Google Scholar]
- Pettmann, B.; Henderson, C.E. Neuronal cell death. Neuron 1998, 20, 633–647. [Google Scholar]
- Mattson, M.P. Neuronal life-and-death signaling, apoptosis, and neurodegenerative disorders. Antioxid. Redox Signal 2006, 8, 1997–2006. [Google Scholar]
- D’Mello, S.R.; Chin, P.C. Treating neurodegenerative conditions through the understanding of neuronal apoptosis. Curr. Drug Targets 2005, 4, 3–23. [Google Scholar]
- Contestabile, A. Cerebellar granule cells as a model to study mechanisms of neuronal apoptosis or survival in vivo and in vitro. Cerebellum 2002, 1, 41–55. [Google Scholar]
- D’Mello, S.R.; Galli, C.; Ciotti, T.; Calissano, P. Induction of apoptosis in cerebellar granule neurons by low potassium: Inhibition of death by insulin-like growth factor i and camp. Proc. Natl. Acad. Sci. USA 1993, 90, 10989–10993. [Google Scholar]
- Gallo, V.; Ciotti, M.T.; Coletti, A.; Aloisi, F.; Levi, G. Selective release of glutamate from cerebellar granule cells differentiating in culture. Proc. Natl. Acad. Sci. USA 1982, 79, 7919–7923. [Google Scholar]
- Isaev, N.K.; Stelmashook, E.V.; Halle, A.; Harms, C.; Lautenschlager, M.; Weih, M.; Dirnagl, U.; Victorov, I.V.; Zorov, D.B. Inhibition of Na+, K+-atpase activity in cultured rat cerebellar granule cells prevents the onset of apoptosis induced by low potassium. Neurosci. Lett 2000, 283, 41–44. [Google Scholar]
- Wood, K.A.; Dipasquale, B.; Youle, R.J. In situ labeling of granule cells for apoptosis-associated DNA fragmentation reveals different mechanisms of cell loss in developing cerebellum. Neuron 1993, 11, 621–632. [Google Scholar]
- Tanaka, M.; Marunouchi, T. Immunohistochemical analysis of developmental stage of external granular layer neurons which undergo apoptosis in postnatal rat cerebellum. Neurosci. Lett 1998, 242, 85–88. [Google Scholar]
- Nikolic, M.; Gardner, H.A.; Tucker, K.L. Postnatal neuronal apoptosis in the cerebral cortex: Physiological and pathophysiological mechanisms. Neuroscience 2013, 254, 369–378. [Google Scholar]
- Galli, C.; Meucci, O.; Scorziello, A.; Werge, T.M.; Calissano, P.; Schettini, G. Apoptosis in cerebellar granule cells is blocked by high kcl, forskolin, and igf-1 through distinct mechanisms of action: The involvement of intracellular calcium and rna synthesis. J. Neurosci 1995, 15, 1172–1179. [Google Scholar]
- Cavallaro, S.; Copani, A.; D’Agata, V.; Musco, S.; Petralia, S.; Ventra, C.; Stivala, F.; Travali, S.; Canonico, P.L. Pituitary adenylate cyclase activating polypeptide prevents apoptosis in cultured cerebellar granule neurons. Mol. Pharmacol 1996, 50, 60–66. [Google Scholar]
- Paratore, S.; Ciotti, M.T.; Basille, M.; Vaudry, D.; Gentile, A.; Parenti, R.; Calissano, P.; Cavallaro, S. Gastric inhibitory polypeptide and its receptor are expressed in the central nervous system and support neuronal survival. Cent. Nerv. Syst. Agent Med. Chem 2011, 11, 210–222. [Google Scholar]
- Inagaki, N.; Seino, Y.; Takeda, J.; Yano, H.; Yamada, Y.; Bell, G.I.; Eddy, R.L.; Fukushima, Y.; Byers, M.G.; Shows, T.B.; et al. Gastric inhibitory polypeptide: Structure and chromosomal localization of the human gene. Mol. Endocrinol 1989, 3, 1014–1021. [Google Scholar]
- Tseng, C.C.; Jarboe, L.A.; Landau, S.B.; Williams, E.K.; Wolfe, M.M. Glucose-dependent insulinotropic peptide: Structure of the precursor and tissue-specific expression in rat. Proc. Natl. Acad. Sci. USA 1993, 90, 1992–1996. [Google Scholar]
- Ojima, A.; Matsui, T.; Maeda, S.; Takeuchi, M.; Yamagishi, S. Glucose-dependent insulinotropic polypeptide (gip) inhibits signaling pathways of advanced glycation end products (ages) in endothelial cells via its antioxidative properties. Horm. Metab. Res 2012, 44, 501–505. [Google Scholar]
- Harada, N. Structure and function of incretin receptor. Nihon Rinsho 2011, 69, 813–820. (in Japanese). [Google Scholar]
- Yamada, Y.; Hayami, T.; Nakamura, K.; Kaisaki, P.J.; Someya, Y.; Wang, C.Z.; Seino, S.; Seino, Y. Human gastric inhibitory polypeptide receptor: Cloning of the gene (gipr) and cdna. Genomics 1995, 29, 773–776. [Google Scholar]
- Gremlich, S.; Porret, A.; Hani, E.H.; Cherif, D.; Vionnet, N.; Froguel, P.; Thorens, B. Cloning, functional expression, and chromosomal localization of the human pancreatic islet glucose-dependent insulinotropic polypeptide receptor. Diabetes 1995, 44, 1202–1208. [Google Scholar]
- Hauner, H.; Glatting, G.; Kaminska, D.; Pfeiffer, E.F. Effects of gastric inhibitory polypeptide on glucose and lipid metabolism of isolated rat adipocytes. Ann. Nutr. Metab 1988, 32, 282–288. [Google Scholar]
- Kogire, M.; Inoue, K.; Sumi, S.; Doi, R.; Yun, M.; Kaji, H.; Tobe, T. Effects of gastric inhibitory polypeptide and glucagon on portal venous and hepatic arterial flow in conscious dogs. Digest. Dis. Sci 1992, 37, 1666–1670. [Google Scholar]
- Holst, J.J. On the physiology of gip and glp-1. Horm. Metab. Res 2004, 36, 747–754. [Google Scholar]
- Hansotia, T.; Drucker, D.J. Gip and glp-1 as incretin hormones: Lessons from single and double incretin receptor knockout mice. Regul. Pept 2005, 128, 125–134. [Google Scholar]
- Gault, V.A.; Harriott, P.; Flatt, P.R.; O’Harte, F.P. Cyclic amp production and insulin releasing activity of synthetic fragment peptides of glucose-dependent insulinotropic polypeptide. Biosci. Rep 2002, 22, 523–528. [Google Scholar]
- Ehses, J.A.; Lee, S.S.; Pederson, R.A.; McIntosh, C.H. A new pathway for glucose-dependent insulinotropic polypeptide (gip) receptor signaling: Evidence for the involvement of phospholipase a2 in gip-stimulated insulin secretion. J. Biol. Chem 2001, 276, 23667–23673. [Google Scholar]
- Volz, A.; Goke, R.; Lankat-Buttgereit, B.; Fehmann, H.C.; Bode, H.P.; Goke, B. Molecular cloning, functional expression, and signal transduction of the gip-receptor cloned from a human insulinoma. FEBS Lett 1995, 373, 23–29. [Google Scholar]
- Brubaker, P.L.; Drucker, D.J. Structure-function of the glucagon receptor family of g protein-coupled receptors: The glucagon, gip, glp-1, and glp-2 receptors. Recept. Channels 2002, 8, 179–188. [Google Scholar]
- Kaplan, A.M.; Vigna, S.R. Gastric inhibitory polypeptide (gip) binding sites in rat brain. Peptides 1994, 15, 297–302. [Google Scholar]
- Gault, V.A.; Holscher, C. Protease-resistant glucose-dependent insulinotropic polypeptide agonists facilitate hippocampal ltp and reverse the impairment of ltp induced by beta-amyloid. J. Neurophysiol 2008, 99, 1590–1595. [Google Scholar]
- Figueiredo, C.P.; Pamplona, F.A.; Mazzuco, T.L.; Aguiar, A.S., Jr.; Walz, R.; Prediger, R.D. Role of the glucose-dependent insulinotropic polypeptide and its receptor in the central nervous system: Therapeutic potential in neurological diseases. Behav. Pharmacol 2010, 21, 394–408. [Google Scholar]
- Faivre, E.; Gault, V.A.; Thorens, B.; Holscher, C. Glucose-dependent insulinotropic polypeptide receptor knockout mice are impaired in learning, synaptic plasticity, and neurogenesis. J. Neurophysiol 2011, 105, 1574–1580. [Google Scholar]
- Buhren, B.A.; Gasis, M.; Thorens, B.; Muller, H.W.; Bosse, F. Glucose-dependent insulinotropic polypeptide (gip) and its receptor (gipr): Cellular localization, lesion-affected expression, and impaired regenerative axonal growth. J. Neurosci. Res 2009, 87, 1858–1870. [Google Scholar]
- Nyberg, J.; Jacobsson, C.; Anderson, M.F.; Eriksson, P.S. Immunohistochemical distribution of glucose-dependent insulinotropic polypeptide in the adult rat brain. J. Neurosci. Res 2007, 85, 2099–2119. [Google Scholar]
- Nyberg, J.; Anderson, M.F.; Meister, B.; Alborn, A.M.; Strom, A.K.; Brederlau, A.; Illerskog, A.C.; Nilsson, O.; Kieffer, T.J.; Hietala, M.A.; et al. Glucose-dependent insulinotropic polypeptide is expressed in adult hippocampus and induces progenitor cell proliferation. J. Neurosci 2005, 25, 1816–1825. [Google Scholar]
- Kim, S.J.; Ao, Z.; Warnock, G.; McIntosh, C.H. Incretin-stimulated interaction between beta-cell kv1.5 and kvbeta2 channel proteins involves acetylation/deacetylation by cbp/sirt1. Biochem. J 2013, 451, 227–234. [Google Scholar]
- Yabe, D.; Seino, Y. Two incretin hormones glp-1 and gip: Comparison of their actions in insulin secretion and beta cell preservation. Prog. Biophys. Mol. Biol 2011, 107, 248–256. [Google Scholar]
- Campbell, R.K. Fate of the beta-cell in the pathophysiology of type 2 diabetes. J. Am. Pharm. Assoc 2009, 49, S10–S15. [Google Scholar]
- Maida, A.; Hansotia, T.; Longuet, C.; Seino, Y.; Drucker, D.J. Differential importance of glucose-dependent insulinotropic polypeptide vs. glucagon-like peptide 1 receptor signaling for beta cell survival in mice. Gastroenterology 2009, 137, 2146–2157. [Google Scholar]
- Widenmaier, S.B.; Ao, Z.; Kim, S.J.; Warnock, G.; McIntosh, C.H. Suppression of p38 mapk and jnk via akt-mediated inhibition of apoptosis signal-regulating kinase 1 constitutes a core component of the beta-cell pro-survival effects of glucose-dependent insulinotropic polypeptide. J. Biol. Chem 2009, 284, 30372–30382. [Google Scholar]
- Kim, S.J.; Nian, C.; Widenmaier, S.; McIntosh, C.H. Glucose-dependent insulinotropic polypeptide-mediated up-regulation of beta-cell antiapoptotic bcl-2 gene expression is coordinated by cyclic amp (camp) response element binding protein (creb) and camp-responsive creb coactivator 2. Mol. Cell. Biol 2008, 28, 1644–1656. [Google Scholar]
- Trumper, A.; Trumper, K.; Horsch, D. Mechanisms of mitogenic and anti-apoptotic signaling by glucose-dependent insulinotropic polypeptide in beta(ins-1)-cells. J. Endocrinol 2002, 174, 233–246. [Google Scholar]
- Kim, S.J.; Winter, K.; Nian, C.; Tsuneoka, M.; Koda, Y.; McIntosh, C.H. Glucose-dependent insulinotropic polypeptide (gip) stimulation of pancreatic beta-cell survival is dependent upon phosphatidylinositol 3-kinase (pi3k)/protein kinase b (pkb) signaling, inactivation of the forkhead transcription factor foxo1, and down-regulation of bax expression. J. Biol. Chem 2005, 280, 22297–22307. [Google Scholar]
- Lyssenko, V.; Eliasson, L.; Kotova, O.; Pilgaard, K.; Wierup, N.; Salehi, A.; Wendt, A.; Jonsson, A.; de Marinis, Y.Z.; Berglund, L.M.; et al. Pleiotropic effects of gip on islet function involve osteopontin. Diabetes 2011, 60, 2424–2433. [Google Scholar]
- Tsukiyama, K.; Yamada, Y.; Yamada, C.; Harada, N.; Kawasaki, Y.; Ogura, M.; Bessho, K.; Li, M.; Amizuka, N.; Sato, M.; et al. Gastric inhibitory polypeptide as an endogenous factor promoting new bone formation after food ingestion. Mol. Endocrinol 2006, 20, 1644–1651. [Google Scholar]
- Kubota, A.; Yamada, Y.; Yasuda, K.; Someya, Y.; Ihara, Y.; Kagimoto, S.; Watanabe, R.; Kuroe, A.; Ishida, H.; Seino, Y. Gastric inhibitory polypeptide activates map kinase through the wortmannin-sensitive and -insensitive pathways. Biochem. Biophys. Res. Commun 1997, 235, 171–175. [Google Scholar]
- Ehses, J.A.; Casilla, V.R.; Doty, T.; Pospisilik, J.A.; Winter, K.D.; Demuth, H.U.; Pederson, R.A.; McIntosh, C.H. Glucose-dependent insulinotropic polypeptide promotes beta-(ins-1) cell survival via cyclic adenosine monophosphate-mediated caspase-3 inhibition and regulation of p38 mitogen-activated protein kinase. Endocrinology 2003, 144, 4433–4445. [Google Scholar]
- Yasuda, K.; Inagaki, N.; Yamada, Y.; Kubota, A.; Seino, S.; Seino, Y. Hamster gastric inhibitory polypeptide receptor expressed in pancreatic islets and clonal insulin-secreting cells: Its structure and functional properties. Biochem. Biophys. Res. Commun 1994, 205, 1556–1562. [Google Scholar]
- Zeng, X.J.; Yu, S.P.; Zhang, L.; Wei, L. Neuroprotective effect of the endogenous neural peptide apelin in cultured mouse cortical neurons. Exp. Cell Res 2010, 316, 1773–1783. [Google Scholar]
- O’Donnell, L.A.; Agrawal, A.; Sabnekar, P.; Dichter, M.A.; Lynch, D.R.; Kolson, D.L. Apelin, an endogenous neuronal peptide, protects hippocampal neurons against excitotoxic injury. J. Neurochem 2007, 102, 1905–1917. [Google Scholar]
- Reaux, A.; de Mota, N.; Skultetyova, I.; Lenkei, Z.; El Messari, S.; Gallatz, K.; Corvol, P.; Palkovits, M.; Llorens-Cortes, C. Physiological role of a novel neuropeptide, apelin, and its receptor in the rat brain. J. Neurochem 2001, 77, 1085–1096. [Google Scholar]
- Cook, D.R.; Gleichman, A.J.; Cross, S.A.; Doshi, S.; Ho, W.; Jordan-Sciutto, K.L.; Lynch, D.R.; Kolson, D.L. Nmda receptor modulation by the neuropeptide apelin: Implications for excitotoxic injury. J. Neurochem 2011, 118, 1113–1123. [Google Scholar]
- Kasai, A.; Kinjo, T.; Ishihara, R.; Sakai, I.; Ishimaru, Y.; Yoshioka, Y.; Yamamuro, A.; Ishige, K.; Ito, Y.; Maeda, S. Apelin deficiency accelerates the progression of amyotrophic lateral sclerosis. PLoS One 2011, 6, e23968. [Google Scholar]
- Morganti, J.M.; Nash, K.R.; Grimmig, B.A.; Ranjit, S.; Small, B.; Bickford, P.C.; Gemma, C. The soluble isoform of cx3cl1 is necessary for neuroprotection in a mouse model of parkinson’s disease. J. Neurosci 2012, 32, 14592–14601. [Google Scholar]
- Hao, F.; Zhang, N.N.; Zhang, D.M.; Bai, H.Y.; Piao, H.; Yuan, B.; Zhu, H.Y.; Yu, H.; Xiao, C.S.; Li, A.P. Chemokine fractalkine attenuates overactivation and apoptosis of bv-2 microglial cells induced by extracellular atp. Neurochem. Res 2013, 38, 1002–1012. [Google Scholar]
- Cipriani, R.; Villa, P.; Chece, G.; Lauro, C.; Paladini, A.; Micotti, E.; Perego, C.; de Simoni, M.G.; Fredholm, B.B.; Eusebi, F.; et al. Cx3cl1 is neuroprotective in permanent focal cerebral ischemia in rodents. J. Neurosci 2011, 31, 16327–16335. [Google Scholar]
- Lauro, C.; Cipriani, R.; Catalano, M.; Trettel, F.; Chece, G.; Brusadin, V.; Antonilli, L.; van Rooijen, N.; Eusebi, F.; Fredholm, B.B.; et al. Adenosine a1 receptors and microglial cells mediate cx3cl1-induced protection of hippocampal neurons against glu-induced death. Neuropsychopharmacology 2010, 35, 1550–1559. [Google Scholar]
- Noda, M.; Doi, Y.; Liang, J.; Kawanokuchi, J.; Sonobe, Y.; Takeuchi, H.; Mizuno, T.; Suzumura, A. Fractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons and antioxidant enzyme heme oxygenase-1 expression. J. Biol. Chem 2011, 286, 2308–2319. [Google Scholar]
- Boehme, S.A.; Lio, F.M.; Maciejewski-Lenoir, D.; Bacon, K.B.; Conlon, P.J. The chemokine fractalkine inhibits fas-mediated cell death of brain microglia. J. Immunol 2000, 165, 397–403. [Google Scholar]
- Cook, A.; Hippensteel, R.; Shimizu, S.; Nicolai, J.; Fatatis, A.; Meucci, O. Interactions between chemokines: Regulation of fractalkine/cx3cl1 homeostasis by sdf/cxcl12 in cortical neurons. J. Biol. Chem 2010, 285, 10563–10571. [Google Scholar]
- Catalano, M.; Lauro, C.; Cipriani, R.; Chece, G.; Ponzetta, A.; di Angelantonio, S.; Ragozzino, D.; Limatola, C. Cx3cl1 protects neurons against excitotoxicity enhancing glt-1 activity on astrocytes. J. Neuroimmunol 2013, 263, 75–82. [Google Scholar]
- Yuan, Q.; Chiquet, B.T.; Devault, L.; Warman, M.L.; Nakamura, Y.; Swindell, E.C.; Hecht, J.T. Craniofacial abnormalities result from knock down of nonsyndromic clefting gene, crispld2, in zebrafish. Genesis 2012, 50, 871–881. [Google Scholar]
- Szlama, G.; Kondas, K.; Trexler, M.; Patthy, L. Wfikkn1 and wfikkn2 bind growth factors tgfbeta1, bmp2 and bmp4 but do not inhibit their signalling activity. FEBS J 2010, 277, 5040–5050. [Google Scholar]
- Liu, Y.; Cheng, H.; Zhou, Y.; Zhu, Y.; Bian, R.; Chen, Y.; Li, C.; Ma, Q.; Zheng, Q.; Zhang, Y.; et al. Myostatin induces mitochondrial metabolic alteration and typical apoptosis in cancer cells. Cell Death Dis 2013, 4, e494. [Google Scholar]
- Holzbaur, E.L.; Howland, D.S.; Weber, N.; Wallace, K.; She, Y.; Kwak, S.; Tchistiakova, L.A.; Murphy, E.; Hinson, J.; Karim, R.; et al. Myostatin inhibition slows muscle atrophy in rodent models of amyotrophic lateral sclerosis. Neurobiol. Dis 2006, 23, 697–707. [Google Scholar]
- Filipe, M.; Goncalves, L.; Bento, M.; Silva, A.C.; Belo, J.A. Comparative expression of mouse and chicken shisa homologues during early development. Dev. Dyn 2006, 235, 2567–2573. [Google Scholar]
- Pei, J.; Grishin, N.V. Unexpected diversity in shisa-like proteins suggests the importance of their roles as transmembrane adaptors. Cell Signal 2012, 24, 758–769. [Google Scholar]
- Schultze, W.; Eulenburg, V.; Lessmann, V.; Herrmann, L.; Dittmar, T.; Gundelfinger, E.D.; Heumann, R.; Erdmann, K.S. Semaphorin4f interacts with the synapse-associated protein sap90/psd-95. J. Neurochem 2001, 78, 482–489. [Google Scholar]
- Barrette, B.; Calvo, E.; Vallieres, N.; Lacroix, S. Transcriptional profiling of the injured sciatic nerve of mice carrying the Wld(s) mutant gene: Identification of genes involved in neuroprotection, neuroinflammation, and nerve regeneration. Brain Behav. Immun 2010, 24, 1254–1267. [Google Scholar]
- Li, T.S.; Li, X.N.; Chang, Z.J.; Fu, X.Y.; Liu, L. Identification and functional characterization of a novel interleukin 17 receptor: A possible mitogenic activation through ras/mitogen-activated protein kinase signaling pathway. Cell Signal 2006, 18, 1287–1298. [Google Scholar]
- Moseley, T.A.; Haudenschild, D.R.; Rose, L.; Reddi, A.H. Interleukin-17 family and il-17 receptors. Cytokine Growth Factor Rev 2003, 14, 155–174. [Google Scholar]
- Vidlak, D.; Kielian, T. Differential effects of interleukin-17 receptor signaling on innate and adaptive immunity during central nervous system bacterial infection. J. Neuroinflam 2012, 9, 128. [Google Scholar]
- Bonni, A.; Brunet, A.; West, A.E.; Datta, S.R.; Takasu, M.A.; Greenberg, M.E. Cell survival promoted by the ras-mapk signaling pathway by transcription-dependent and independent mechanisms. Science 1999, 286, 1358–1362. [Google Scholar]
- Holt, E.H.; Broadus, A.E.; Brines, M.L. Parathyroid hormone-related peptide is produced by cultured cerebellar granule cells in response to l-type voltage-sensitive Ca2+ channel flux via a Ca2+/calmodulin-dependent kinase pathway. J. Biol. Chem 1996, 271, 28105–28111. [Google Scholar]
- Ono, T.; Inokuchi, K.; Ogura, A.; Ikawa, Y.; Kudo, Y.; Kawashima, S. Activity-dependent expression of parathyroid hormone-related protein (pthrp) in rat cerebellar granule neurons. Requirement of pthrp for the activity-dependent survival of granule neurons. J. Biol. Chem 1997, 272, 14404–14411. [Google Scholar]
- Chatterjee, O.; Nakchbandi, I.A.; Philbrick, W.M.; Dreyer, B.E.; Zhang, J.P.; Kaczmarek, L.K.; Brines, M.L.; Broadus, A.E. Endogenous parathyroid hormone-related protein functions as a neuroprotective agent. Brain Res 2002, 930, 58–66. [Google Scholar]
- Brines, M.L.; Ling, Z.; Broadus, A.E. Parathyroid hormone-related protein protects against kainic acid excitotoxicity in rat cerebellar granule cells by regulating l-type channel calcium flux. Neurosci. Lett 1999, 274, 13–16. [Google Scholar]
- Gessi, M.; Monego, G.; Calviello, G.; Lanza, P.; Giangaspero, F.; Silvestrini, A.; Lauriola, L.; Ranelletti, F.O. Human parathyroid hormone-related protein and human parathyroid hormone receptor type 1 are expressed in human medulloblastomas and regulate cell proliferation and apoptosis in medulloblastoma-derived cell lines. Acta Neuropathol 2007, 114, 135–145. [Google Scholar]
- Garcia-Esparcia, P.; Schluter, A.; Carmona, M.; Moreno, J.; Ansoleaga, B.; Torrejon-Escribano, B.; Gustincich, S.; Pujol, A.; Ferrer, I. Functional genomics reveals dysregulation of cortical olfactory receptors in parkinson disease: Novel putative chemoreceptors in the human brain. J. Neuropathol. Exp. Neurol 2013, 72, 524–539. [Google Scholar]
- Shati, A.A.; Elsaid, F.G.; Hafez, E.E. Biochemical and molecular aspects of aluminium chloride-induced neurotoxicity in mice and the protective role of crocus sativus l. Extraction and honey syrup. Neuroscience 2011, 175, 66–74. [Google Scholar]
- Juhasz, K.; Zvara, A.; Lipp, A.M.; Nimmervoll, B.; Sonnleitner, A.; Balogi, Z.; Duda, E. Casein kinase 2-interacting protein-1, an inflammatory signaling molecule interferes with tnf reverse signaling in human model cells. Immunol. Lett 2013, 152, 55–64. [Google Scholar]
- Tsunoda, T.; Ota, T.; Fujimoto, T.; Doi, K.; Tanaka, Y.; Yoshida, Y.; Ogawa, M.; Matsuzaki, H.; Hamabashiri, M.; Tyson, D.R.; et al. Inhibition of phosphodiesterase-4 (pde4) activity triggers luminal apoptosis and akt dephosphorylation in a 3-d colonic-crypt model. Mol. Cancer 2012, 11, 46. [Google Scholar]
- Kim, S.W.; Rai, D.; McKeller, M.R.; Aguiar, R.C. Rational combined targeting of phosphodiesterase 4b and syk in dlbcl. Blood 2009, 113, 6153–6160. [Google Scholar]
- Svoboda, N.; Zierler, S.; Kerschbaum, H.H. Camp mediates ammonia-induced programmed cell death in the microglial cell line bv-2. Eur. J. Neurosci 2007, 25, 2285–2295. [Google Scholar]
- Fatemi, S.H.; King, D.P.; Reutiman, T.J.; Folsom, T.D.; Laurence, J.A.; Lee, S.; Fan, Y.T.; Paciga, S.A.; Conti, M.; Menniti, F.S. Pde4b polymorphisms and decreased pde4b expression are associated with schizophrenia. Schizophr. Res 2008, 101, 36–49. [Google Scholar]
- Zeng, W.; Kong, Q.; Li, C.; Mao, B. Xenopus rcor2 (rest corepressor 2) interacts with zmynd8, which is involved in neural differentiation. Biochem. Biophys. Res. Commun 2010, 394, 1024–1029. [Google Scholar]
- Lee, S.H.; Chen, T.; Zhou, J.; Hofmann, J.; Bepler, G. Protein kinase c-beta gene variants, pathway activation, and enzastaurin activity in lung cancer. Clin. Lung Cancer 2010, 11, 169–175. [Google Scholar]
- Lim, S.; Naisbitt, S.; Yoon, J.; Hwang, J.I.; Suh, P.G.; Sheng, M.; Kim, E. Characterization of the shank family of synaptic proteins. Multiple genes, alternative splicing, and differential expression in brain and development. J. Biol. Chem 1999, 274, 29510–29518. [Google Scholar]
- Boeckers, T.M.; Bockmann, J.; Kreutz, M.R.; Gundelfinger, E.D. Prosap/shank proteins—A family of higher order organizing molecules of the postsynaptic density with an emerging role in human neurological disease. J. Neurochem 2002, 81, 903–910. [Google Scholar]
- Park, E.; Na, M.; Choi, J.; Kim, S.; Lee, J.R.; Yoon, J.; Park, D.; Sheng, M.; Kim, E. The shank family of postsynaptic density proteins interacts with and promotes synaptic accumulation of the beta pix guanine nucleotide exchange factor for rac1 and cdc42. J. Biol. Chem 2003, 278, 19220–19229. [Google Scholar]
- Naisbitt, S.; Kim, E.; Weinberg, R.J.; Rao, A.; Yang, F.C.; Craig, A.M.; Sheng, M. Characterization of guanylate kinase-associated protein, a postsynaptic density protein at excitatory synapses that interacts directly with postsynaptic density-95/synapse-associated protein 90. J. Neurosci 1997, 17, 5687–5696. [Google Scholar]
- Boeckers, T.M.; Winter, C.; Smalla, K.H.; Kreutz, M.R.; Bockmann, J.; Seidenbecher, C.; Garner, C.C.; Gundelfinger, E.D. Proline-rich synapse-associated proteins prosap1 and prosap2 interact with synaptic proteins of the sapap/gkap family. Biochem. Biophys. Res. Commun 1999, 264, 247–252. [Google Scholar]
- Moutin, E.; Raynaud, F.; Fagni, L.; Perroy, J. Gkap-dlc2 interaction organizes the postsynaptic scaffold complex to enhance synaptic nmda receptor activity. J. Cell Sci 2012, 125, 2030–2040. [Google Scholar]
- Grabrucker, A.M. A role for synaptic zinc in prosap/shank psd scaffold malformation in autism spectrum disorders. Dev. Neurobiol 2014, 74, 136–146. [Google Scholar]
- Li, J.M.; Lu, C.L.; Cheng, M.C.; Luu, S.U.; Hsu, S.H.; Chen, C.H. Genetic analysis of the dlgap1 gene as a candidate gene for schizophrenia. Psychiatry Res 2013, 205, 13–17. [Google Scholar]
- Liu, C.; Lin, C.; Whitaker, D.T.; Bakeri, H.; Bulgakov, O.V.; Liu, P.; Lei, J.; Dong, L.; Li, T.; Swaroop, A. Prickle1 is expressed in distinct cell populations of the central nervous system and contributes to neuronal morphogenesis. Hum. Mol. Genet 2013, 22, 2234–2246. [Google Scholar]
- Fujimura, L.; Hatano, M. Role of prickle1 and prickle2 in neurite outgrowth in murine neuroblastoma cells. Methods Mol. Biol 2012, 839, 173–185. [Google Scholar]
- Fujimura, L.; Watanabe-Takano, H.; Sato, Y.; Tokuhisa, T.; Hatano, M. Prickle promotes neurite outgrowth via the dishevelled dependent pathway in c1300 cells. Neurosci. Lett 2009, 467, 6–10. [Google Scholar]
- Bassuk, A.G.; Wallace, R.H.; Buhr, A.; Buller, A.R.; Afawi, Z.; Shimojo, M.; Miyata, S.; Chen, S.; Gonzalez-Alegre, P.; Griesbach, H.L.; et al. A homozygous mutation in human prickle1 causes an autosomal-recessive progressive myoclonus epilepsy-ataxia syndrome. Am. J. Hum. Genet 2008, 83, 572–581. [Google Scholar]
- Toyoshima, Y.; Karas, M.; Yakar, S.; Dupont, J.; Lee, H.; LeRoith, D. Tdag51 mediates the effects of insulin-like growth factor i (igf-i) on cell survival. J. Biol. Chem 2004, 279, 25898–25904. [Google Scholar]
- Murata, T.; Sato, T.; Kamoda, T.; Moriyama, H.; Kumazawa, Y.; Hanada, N. Differential susceptibility to hydrogen sulfide-induced apoptosis between phlda1-overexpressing oral cancer cell lines and oral keratinocytes: Role of phlda1 as an apoptosis suppressor. Exp. Cell Res 2014, 320, 247–257. [Google Scholar]
- Park, E.S.; Kim, J.; Ha, T.U.; Choi, J.S.; Soo Hong, K.; Rho, J. Tdag51 deficiency promotes oxidative stress-induced apoptosis through the generation of reactive oxygen species in mouse embryonic fibroblasts. Exp. Mol. Med 2013, 45, e35. [Google Scholar]
- Xi, Z.Q.; Wang, L.Y.; Sun, J.J.; Liu, X.Z.; Zhu, X.; Xiao, F.; Guan, L.F.; Li, J.M.; Wang, L.; Wang, X.F. Tdag51 in the anterior temporal neocortex of patients with intractable epilepsy. Neurosci. Lett 2007, 425, 53–58. [Google Scholar]
- Van Wayenbergh, R.; Taelman, V.; Pichon, B.; Fischer, A.; Kricha, S.; Gessler, M.; Christophe, D.; Bellefroid, E.J. Identification of boip, a novel cdna highly expressed during spermatogenesis that encodes a protein interacting with the orange domain of the hairy-related transcription factor hrt1/hey1 in xenopus and mouse. Dev. Dyn 2003, 228, 716–725. [Google Scholar]
- Jacot, T.A.; Zalenskaya, I.; Mauck, C.; Archer, D.F.; Doncel, G.F. Tspy4 is a novel sperm-specific biomarker of semen exposure in human cervicovaginal fluids; potential use in hiv prevention and contraception studies. Contraception 2013, 88, 387–395. [Google Scholar]
- Kido, T.; Schubert, S.; Schmidtke, J.; Chris Lau, Y.F. Expression of the human tspy gene in the brains of transgenic mice suggests a potential role of this y chromosome gene in neural functions. Yi Chuan Xue Bao 2011, 38, 181–191. (in Chinese). [Google Scholar]
- Williams, M.; Lyu, M.S.; Yang, Y.L.; Lin, E.P.; Dunbrack, R.; Birren, B.; Cunningham, J.; Hunter, K. Ier5, a novel member of the slow-kinetics immediate-early genes. Genomics 1999, 55, 327–334. [Google Scholar]
- Cirelli, C.; Tononi, G. Gene expression in the brain across the sleep-waking cycle. Brain Res 2000, 885, 303–321. [Google Scholar]
- Baye, T.M.; Wilke, R.A.; Olivier, M. Genomic and geographic distribution of private snps and pathways in human populations. Personal. Med 2009, 6, 623–641. [Google Scholar]
- Konishi, Y.; Aoki, T.; Ohkawa, N.; Matsu-Ura, T.; Mikoshiba, K.; Tamura, T. Identification of the C-terminal activation domain of the neurod-related factor (ndrf). Nucleic Acids Res 2000, 28, 2406–2412. [Google Scholar]
- Bormuth, I.; Yan, K.; Yonemasu, T.; Gummert, M.; Zhang, M.; Wichert, S.; Grishina, O.; Pieper, A.; Zhang, W.; Goebbels, S.; et al. Neuronal basic helix-loop-helix proteins neurod2/6 regulate cortical commissure formation before midline interactions. J. Neurosci 2013, 33, 641–651. [Google Scholar]
- Schwab, M.H.; Bartholomae, A.; Heimrich, B.; Feldmeyer, D.; Druffel-Augustin, S.; Goebbels, S.; Naya, F.J.; Zhao, S.; Frotscher, M.; Tsai, M.J.; et al. Neuronal basic helix-loop-helix proteins (nex and beta2/neuro d) regulate terminal granule cell differentiation in the hippocampus. J. Neurosci 2000, 20, 3714–3724. [Google Scholar]
- Konishi, Y.; Matsu-ura, T.; Mikoshiba, K.; Tamura, T. Stimulation of gene expression of neurod-related factor in the mouse brain following pentylenetetrazol-induced seizures. Brain Res 2001, 97, 129–136. [Google Scholar]
- Lin, C.H.; Hansen, S.; Wang, Z.; Storm, D.R.; Tapscott, S.J.; Olson, J.M. The dosage of the neurod2 transcription factor regulates amygdala development and emotional learning. Proc. Natl. Acad. Sci. USA 2005, 102, 14877–14882. [Google Scholar]
- Lin, C.H.; Stoeck, J.; Ravanpay, A.C.; Guillemot, F.; Tapscott, S.J.; Olson, J.M. Regulation of neurod2 expression in mouse brain. Dev. Biol 2004, 265, 234–245. [Google Scholar]
- Olson, J.M.; Asakura, A.; Snider, L.; Hawkes, R.; Strand, A.; Stoeck, J.; Hallahan, A.; Pritchard, J.; Tapscott, S.J. Neurod2 is necessary for development and survival of central nervous system neurons. Dev. Biol 2001, 234, 174–187. [Google Scholar]
- Flames, N.; Hobert, O. Gene regulatory logic of dopamine neuron differentiation. Nature 2009, 458, 885–889. [Google Scholar]
- Abe, H.; Okazawa, M.; Nakanishi, S. The etv1/er81 transcription factor orchestrates activity-dependent gene regulation in the terminal maturation program of cerebellar granule cells. Proc. Natl. Acad. Sci. USA 2011, 108, 12497–12502. [Google Scholar]
- Abe, H.; Okazawa, M.; Nakanishi, S. Gene regulation via excitation and bdnf is mediated by induction and phosphorylation of the etv1 transcription factor in cerebellar granule cells. Proc. Natl. Acad. Sci. USA 2012, 109, 8734–8739. [Google Scholar]
- Tuoc, T.C.; Stoykova, A. Er81 is a downstream target of pax6 in cortical progenitors. BMC Dev. Biol 2008, 8, 23. [Google Scholar]
- Arber, S.; Ladle, D.R.; Lin, J.H.; Frank, E.; Jessell, T.M. Ets gene er81 controls the formation of functional connections between group ia sensory afferents and motor neurons. Cell 2000, 101, 485–498. [Google Scholar]
- Zhang, Z.; Pinto, A.M.; Wan, L.; Wang, W.; Berg, M.G.; Oliva, I.; Singh, L.N.; Dengler, C.; Wei, Z.; Dreyfuss, G. Dysregulation of synaptogenesis genes antecedes motor neuron pathology in spinal muscular atrophy. Proc. Natl. Acad. Sci. USA 2013, 110, 19348–19353. [Google Scholar]
- Chen, Y.; Zou, H.; Yang, L.Y.; Li, Y.; Wang, L.; Hao, Y.; Yang, J.L. Er81-shrna inhibits growth of triple-negative human breast cancer cell line mda-mb-231 in vivo and in vitro. Asian Pac. J. Cancer Prev. 2012, 13, 2385–2392. [Google Scholar]
- Scheer, E.; Delbac, F.; Tora, L.; Moras, D.; Romier, C. Tfiid taf6–taf9 complex formation involves the heat repeat-containing C-terminal domain of taf6 and is modulated by taf5 protein. J. Biol. Chem 2012, 287, 27580–27592. [Google Scholar]
- Wilhelm, E.; Kornete, M.; Targat, B.; Vigneault-Edwards, J.; Frontini, M.; Tora, L.; Benecke, A.; Bell, B. Taf6delta orchestrates an apoptotic transcriptome profile and interacts functionally with p53. BMC Mol. Biol 2010, 11, 10. [Google Scholar]
- Mizuta, T.R.; Fukita, Y.; Miyoshi, T.; Shimizu, A.; Honjo, T. Isolation of cdna encoding a binding protein specific to 5′-phosphorylated single-stranded DNA with g-rich sequences. Nucleic Acids Res 1993, 21, 1761–1766. [Google Scholar]
- Krieger, F.; Elflein, N.; Ruiz, R.; Guerra, J.; Serrano, A.L.; Asan, E.; Tabares, L.; Jablonka, S. Fast motor axon loss in smard1 does not correspond to morphological and functional alterations of the NMJ. Neurobiol. Dis 2013, 54, 169–182. [Google Scholar]
- Tachi, N.; Kikuchi, S.; Kozuka, N.; Nogami, A. A new mutation of ighmbp2 gene in spinal muscular atrophy with respiratory distress type 1. Pediatr. Neurol 2005, 32, 288–290. [Google Scholar]
- Hinks, G.L.; Shah, B.; French, S.J.; Campos, L.S.; Staley, K.; Hughes, J.; Sofroniew, M.V. Expression of lim protein genes lmo1, lmo2, and lmo3 in adult mouse hippocampus and other forebrain regions: Differential regulation by seizure activity. J. Neurosci 1997, 17, 5549–5559. [Google Scholar]
- Herberth, B.; Minko, K.; Csillag, A.; Jaffredo, T.; Madarasz, E. Scl, gata-2 and lmo2 expression in neurogenesis. Int. J. Dev. Neurosci 2005, 23, 449–463. [Google Scholar]
- Kulemzina, I.; Schumacher, M.R.; Verma, V.; Reiter, J.; Metzler, J.; Failla, A.V.; Lanz, C.; Sreedharan, V.T.; Ratsch, G.; Ivanov, D. Cohesin rings devoid of scc3 and pds5 maintain their stable association with the DNA. PLoS Genet 2012, 8, e1002856. [Google Scholar]
- Chan, K.L.; Gligoris, T.; Upcher, W.; Kato, Y.; Shirahige, K.; Nasmyth, K.; Beckouet, F. Pds5 promotes and protects cohesin acetylation. Proc. Natl. Acad. Sci. USA 2013, 110, 13020–13025. [Google Scholar]
- Ren, Q.; Yang, H.; Rosinski, M.; Conrad, M.N.; Dresser, M.E.; Guacci, V.; Zhang, Z. Mutation of the cohesin related gene pds5 causes cell death with predominant apoptotic features in saccharomyces cerevisiae during early meiosis. Mutat. Res 2005, 570, 163–173. [Google Scholar]
- Zhang, B.; Chang, J.; Fu, M.; Huang, J.; Kashyap, R.; Salavaggione, E.; Jain, S.; Kulkarni, S.; Deardorff, M.A.; Uzielli, M.L.; et al. Dosage effects of cohesin regulatory factor pds5 on mammalian development: Implications for cohesinopathies. PLoS One 2009, 4, e5232. [Google Scholar]
- Hur, S.K.; Park, E.J.; Han, J.E.; Kim, Y.A.; Kim, J.D.; Kang, D.; Kwon, J. Roles of human ino80 chromatin remodeling enzyme in DNA replication and chromosome segregation suppress genome instability. Cell. Mol. Life Sci 2010, 67, 2283–2296. [Google Scholar]
- Seeber, A.; Dion, V.; Gasser, S.M. Checkpoint kinases and the ino80 nucleosome remodeling complex enhance global chromatin mobility in response to DNA damage. Genes Dev 2013, 27, 1999–2008. [Google Scholar]
- Park, E.J.; Hur, S.K.; Kwon, J. Human ino80 chromatin-remodelling complex contributes to DNA double-strand break repair via the expression of rad54b and xrcc3 genes. Biochem. J 2010, 431, 179–187. [Google Scholar]
- Fischle, W.; Franz, H.; Jacobs, S.A.; Allis, C.D.; Khorasanizadeh, S. Specificity of the chromodomain y chromosome family of chromodomains for lysine-methylated ark(s/t) motifs. J. Biol. Chem 2008, 283, 19626–19635. [Google Scholar]
- Gu, P.; Min, J.N.; Wang, Y.; Huang, C.; Peng, T.; Chai, W.; Chang, S. Ctc1 deletion results in defective telomere replication, leading to catastrophic telomere loss and stem cell exhaustion. EMBO J 2012, 31, 2309–2321. [Google Scholar]
- Chen, L.Y.; Redon, S.; Lingner, J. The human cst complex is a terminator of telomerase activity. Nature 2012, 488, 540–544. [Google Scholar]
- Kasbek, C.; Wang, F.; Price, C.M. Human ten1 maintains telomere integrity and functions in genome-wide replication restart. J. Biol. Chem 2013, 288, 30139–30150. [Google Scholar]
- Gramatges, M.M.; Bertuch, A.A. Short telomeres: From dyskeratosis congenita to sporadic aplastic anemia and malignancy. Transl. Res 2013, 162, 353–363. [Google Scholar]
- Lee, K.D.; Pai, M.Y.; Hsu, C.C.; Chen, C.C.; Chen, Y.L.; Chu, P.Y.; Lee, C.H.; Chen, L.T.; Chang, J.Y.; Huang, T.H.; et al. Targeted casp8ap2 methylation increases drug resistance in mesenchymal stem cells and cancer cells. Biochem. Biophys. Res. Commun 2012, 422, 578–585. [Google Scholar]
- Kumral, A.; Tuzun, F.; Tugyan, K.; Ozbal, S.; Yilmaz, O.; Yesilirmak, C.D.; Duman, N.; Ozkan, H. Role of epigenetic regulatory mechanisms in neonatal hypoxic-ischemic brain injury. Early Hum. Dev 2013, 89, 165–173. [Google Scholar]
- Imai, Y.; Kimura, T.; Murakami, A.; Yajima, N.; Sakamaki, K.; Yonehara, S. The ced-4-homologous protein flash is involved in fas-mediated activation of caspase-8 during apoptosis. Nature 1999, 398, 777–785. [Google Scholar]
- Yang, X.C.; Burch, B.D.; Yan, Y.; Marzluff, W.F.; Dominski, Z. Flash, a proapoptotic protein involved in activation of caspase-8, is essential for 3′ end processing of histone pre-mrnas. Mol. Cell 2009, 36, 267–278. [Google Scholar]
- Burmester, T.; Ebner, B.; Weich, B.; Hankeln, T. Cytoglobin: A novel globin type ubiquitously expressed in vertebrate tissues. Mol. Biol. Evolut 2002, 19, 416–421. [Google Scholar]
- Ascenzi, P.; Marino, M.; Polticelli, F.; Coletta, M.; Gioia, M.; Marini, S.; Pesce, A.; Nardini, M.; Bolognesi, M.; Reeder, B.J.; et al. Non-covalent and covalent modifications modulate the reactivity of monomeric mammalian globins. Biochim. Biophys. Acta 2013, 1834, 1750–1756. [Google Scholar]
- Tian, S.F.; Yang, H.H.; Xiao, D.P.; Huang, Y.J.; He, G.Y.; Ma, H.R.; Xia, F.; Shi, X.C. Mechanisms of neuroprotection from hypoxia-ischemia (hi) brain injury by up-regulation of cytoglobin (cygb) in a neonatal rat model. J. Biol. Chem 2013, 288, 15988–16003. [Google Scholar]
- Hundahl, C.A.; Elfving, B.; Muller, H.K.; Hay-Schmidt, A.; Wegener, G. A gene-environment study of cytoglobin in the human and rat hippocampus. PLoS One 2013, 8, e63288. [Google Scholar]
- Beltran-Parrazal, L.; Acuna, D.; Ngan, A.M.; Kim, E.; Ngan, A.; Kawakami, K.; Edmond, J.; Lopez, I.A. Neuroglobin, cytoglobin, and transcriptional profiling of hypoxia-related genes in the rat cerebellum after prenatal chronic very mild carbon monoxide exposure (25 ppm). Brain Res 2010, 1330, 61–71. [Google Scholar]
- Araki, T.; Milbrandt, J. Znrf proteins constitute a family of presynaptic e3 ubiquitin ligases. J. Neurosci 2003, 23, 9385–9394. [Google Scholar]
- Yoshida, K.; Watanabe, M.; Hatakeyama, S. Znrf1 interacts with tubulin and regulates cell morphogenesis. Biochem. Biophys. Res. Commun 2009, 389, 506–511. [Google Scholar]
- Saitoh, F.; Araki, T. Proteasomal degradation of glutamine synthetase regulates schwann cell differentiation. J. Neurosci 2010, 30, 1204–1212. [Google Scholar]
- Tidd, D.M.; Broughton, C.M.; Clark, R.E. Cpg oligodeoxynucleotide 5mer-induced apoptosis in molt-4 leukaemia cells does not require caspase 3 or new protein synthesis. Nucleic Acids Res 2003, 31, 2297–2304. [Google Scholar]
- Matsuki, H.; Takahashi, M.; Higuchi, M.; Makokha, G.N.; Oie, M.; Fujii, M. Both g3bp1 and g3bp2 contribute to stress granule formation. Genes Cells 2013, 18, 135–146. [Google Scholar]
- Takahashi, M.; Higuchi, M.; Matsuki, H.; Yoshita, M.; Ohsawa, T.; Oie, M.; Fujii, M. Stress granules inhibit apoptosis by reducing reactive oxygen species production. Mol. Cell. Biol 2013, 33, 815–829. [Google Scholar]
- Maretzky, T.; McIlwain, D.R.; Issuree, P.D.; Li, X.; Malapeira, J.; Amin, S.; Lang, P.A.; Mak, T.W.; Blobel, C.P. Irhom2 controls the substrate selectivity of stimulated adam17-dependent ectodomain shedding. Proc. Natl. Acad. Sci. USA 2013, 110, 11433–11438. [Google Scholar]
- Christova, Y.; Adrain, C.; Bambrough, P.; Ibrahim, A.; Freeman, M. Mammalian irhoms have distinct physiological functions including an essential role in tace regulation. EMBO Rep 2013, 14, 884–890. [Google Scholar]
- Hurtado, O.; Lizasoain, I.; Fernandez-Tome, P.; Alvarez-Barrientos, A.; Leza, J.C.; Lorenzo, P.; Moro, M.A. Tace/adam17-tnf-alpha pathway in rat cortical cultures after exposure to oxygen-glucose deprivation or glutamate. J. Cereb. Blood Flow Metab 2002, 22, 576–585. [Google Scholar]
- Meliopoulos, V.A.; Andersen, L.E.; Brooks, P.; Yan, X.; Bakre, A.; Coleman, J.K.; Tompkins, S.M.; Tripp, R.A. Microrna regulation of human protease genes essential for influenza virus replication. PLoS One 2012, 7, e37169. [Google Scholar]
- Liu, Y.; Kern, J.T.; Walker, J.R.; Johnson, J.A.; Schultz, P.G.; Luesch, H. A genomic screen for activators of the antioxidant response element. Proc. Natl. Acad. Sci. USA 2007, 104, 5205–5210. [Google Scholar]
- Staneviciene, I.; Sadauskiene, I.; Lesauskaite, V.; Ivanoviene, L.; Kasauskas, A.; Ivanov, L. Subacute effects of cadmium and zinc ions on protein synthesis and cell death in mouse liver. Medicina 2008, 44, 131–138. [Google Scholar]
- Munakata, K.; Iwamoto, K.; Bundo, M.; Kato, T. Mitochondrial DNA 3243A > G mutation and increased expression of lars2 gene in the brains of patients with bipolar disorder and schizophrenia. Biol. Psychiatry 2005, 57, 525–532. [Google Scholar]
- Haitina, T.; Lindblom, J.; Renstrom, T.; Fredriksson, R. Fourteen novel human members of mitochondrial solute carrier family 25 (slc25) widely expressed in the central nervous system. Genomics 2006, 88, 779–790. [Google Scholar]
- Fiermonte, G.; Paradies, E.; Todisco, S.; Marobbio, C.M.; Palmieri, F. A novel member of solute carrier family 25 (slc25a42) is a transporter of coenzyme a and adenosine 3′,5′-diphosphate in human mitochondria. J. Biol. Chem 2009, 284, 18152–18159. [Google Scholar]
- Palmieri, F. The mitochondrial transporter family (slc25): Physiological and pathological implications. Pflugers Arch 2004, 447, 689–709. [Google Scholar]
- Zhang, H.; Chen, X.; Sairam, M.R. Novel genes of visceral adiposity: Identification of mouse and human mesenteric estrogen-dependent adipose (meda)-4 gene and its adipogenic function. Endocrinology 2012, 153, 2665–2676. [Google Scholar]
- Stolzing, A.; Grune, T. Neuronal apoptotic bodies: Phagocytosis and degradation by primary microglial cells. FASEB J 2004, 18, 743–745. [Google Scholar]
- Prager-Khoutorsky, M.; Spira, M.E. Neurite retraction and regrowth regulated by membrane retrieval, membrane supply, and actin dynamics. Brain Res 2009, 1251, 65–79. [Google Scholar]
- Klein, M.E.; Younts, T.J.; Castillo, P.E.; Jordan, B.A. Rna-binding protein sam68 controls synapse number and local beta-actin mrna metabolism in dendrites. Proc. Natl. Acad. Sci. USA 2013, 110, 3125–3130. [Google Scholar]
- Ghosh, T.; Soni, K.; Scaria, V.; Halimani, M.; Bhattacharjee, C.; Pillai, B. Microrna-mediated up-regulation of an alternatively polyadenylated variant of the mouse cytoplasmic β-actin gene. Nucleic Acids Res 2008, 36, 6318–6332. [Google Scholar]
- Ferreira, E.; Cronje, M.J. Selection of suitable reference genes for quantitative real-time PCR in apoptosis-induced mcf-7 breast cancer cells. Mol. Biotechnol 2012, 50, 121–128. [Google Scholar]
- Riviere, J.B.; van Bon, B.W.; Hoischen, A.; Kholmanskikh, S.S.; O’Roak, B.J.; Gilissen, C.; Gijsen, S.; Sullivan, C.T.; Christian, S.L.; Abdul-Rahman, O.A.; et al. De novo mutations in the actin genes actb and actg1 cause baraitser-winter syndrome. Nat. Genet 2012, 44, 440–444. [Google Scholar]
- Miwa, T.; Manabe, Y.; Kurokawa, K.; Kamada, S.; Kanda, N.; Bruns, G.; Ueyama, H.; Kakunaga, T. Structure, chromosome location, and expression of the human smooth muscle (enteric type) γ-actin gene: Evolution of six human actin genes. Mol. Cell. Biol 1991, 11, 3296–3306. [Google Scholar]
- Kumar, A.; Crawford, K.; Close, L.; Madison, M.; Lorenz, J.; Doetschman, T.; Pawlowski, S.; Duffy, J.; Neumann, J.; Robbins, J.; et al. Rescue of cardiac alpha-actin-deficient mice by enteric smooth muscle gamma-actin. Proc. Natl. Acad. Sci. USA 1997, 94, 4406–4411. [Google Scholar]
- Li, G.Y.; Kim, M.; Kim, J.H.; Lee, M.O.; Chung, J.H.; Lee, B.H. Gene expression profiling in human lung fibroblast following cadmium exposure. Food Chem. Toxicol 2008, 46, 1131–1137. [Google Scholar]
- Marshall, T.W.; Aloor, H.L.; Bear, J.E. Coronin 2a regulates a subset of focal-adhesion-turnover events through the cofilin pathway. J. Cell Sci 2009, 122, 3061–3069. [Google Scholar]
- Huang, W.; Ghisletti, S.; Saijo, K.; Gandhi, M.; Aouadi, M.; Tesz, G.J.; Zhang, D.X.; Yao, J.; Czech, M.P.; Goode, B.L.; et al. Coronin 2a mediates actin-dependent de-repression of inflammatory response genes. Nature 2011, 470, 414–418. [Google Scholar]
- Franco, D.L.; Rezaval, C.; Caceres, A.; Schinder, A.F.; Ceriani, M.F. Ena/vasp downregulation triggers cell death by impairing axonal maintenance in hippocampal neurons. Mol. Cell. Neurosci 2010, 44, 154–164. [Google Scholar]
- Krause, M.; Leslie, J.D.; Stewart, M.; Lafuente, E.M.; Valderrama, F.; Jagannathan, R.; Strasser, G.A.; Rubinson, D.A.; Liu, H.; Way, M.; et al. Lamellipodin, an ena/vasp ligand, is implicated in the regulation of lamellipodial dynamics. Dev. Cell 2004, 7, 571–583. [Google Scholar]
- Klostermann, A.; Lutz, B.; Gertler, F.; Behl, C. The orthologous human and murine semaphorin 6a-1 proteins (sema6a-1/sema6a-1) bind to the enabled/vasodilator-stimulated phosphoprotein-like protein (evl) via a novel carboxyl-terminal zyxin-like domain. J. Biol. Chem 2000, 275, 39647–39653. [Google Scholar]
- Kubota, K.; Kim, J.Y.; Sawada, A.; Tokimasa, S.; Fujisaki, H.; Matsuda-Hashii, Y.; Ozono, K.; Hara, J. Lrrc8 involved in b cell development belongs to a novel family of leucine-rich repeat proteins. FEBS Lett 2004, 564, 147–152. [Google Scholar]
- Herrick, S.; Evers, D.M.; Lee, J.Y.; Udagawa, N.; Pak, D.T. Postsynaptic pdlim5/enigma homolog binds spar and causes dendritic spine shrinkage. Mol. Cell. Neurosci 2010, 43, 188–200. [Google Scholar]
- Hussain, N.K.; Yamabhai, M.; Bhakar, A.L.; Metzler, M.; Ferguson, S.S.; Hayden, M.R.; McPherson, P.S.; Kay, B.K. A role for epsin n-terminal homology/ap180 n-terminal homology (enth/anth) domains in tubulin binding. J. Biol. Chem 2003, 278, 28823–28830. [Google Scholar]
- Romanitan, M.O.; Popescu, B.O.; Spulber, S.; Bajenaru, O.; Popescu, L.M.; Winblad, B.; Bogdanovic, N. Altered expression of claudin family proteins in alzheimer’s disease and vascular dementia brains. J. Cell. Mol. Med 2010, 14, 1088–1100. [Google Scholar]
- Bruggeman, L.A.; Martinka, S.; Simske, J.S. Expression of tm4sf10, a claudin/emp/pmp22 family cell junction protein, during mouse kidney development and podocyte differentiation. Dev. Dyn 2007, 236, 596–605. [Google Scholar]
- Laketa, V.; Simpson, J.C.; Bechtel, S.; Wiemann, S.; Pepperkok, R. High-content microscopy identifies new neurite outgrowth regulators. Mol. Biol. Cell 2007, 18, 242–252. [Google Scholar]
- Laumet, G.; Petitprez, V.; Sillaire, A.; Ayral, A.M.; Hansmannel, F.; Chapuis, J.; Hannequin, D.; Pasquier, F.; Scarpini, E.; Galimberti, D.; et al. A study of the association between the adam12 and sh3pxd2a (sh3md1) genes and Alzheimer’s disease. Neurosci. Lett 2010, 468, 1–2. [Google Scholar]
- Harold, D.; Jehu, L.; Turic, D.; Hollingworth, P.; Moore, P.; Summerhayes, P.; Moskvina, V.; Foy, C.; Archer, N.; Hamilton, B.A.; et al. Interaction between the adam12 and sh3md1 genes may confer susceptibility to late-onset alzheimer’s disease. Am. J. Med. Genet. B 2007, 144B, 448–452. [Google Scholar]
- Malinin, N.L.; Wright, S.; Seubert, P.; Schenk, D.; Griswold-Prenner, I. Amyloid-β neurotoxicity is mediated by fish adapter protein and adam12 metalloprotease activity. Proc. Natl. Acad. Sci. USA 2005, 102, 3058–3063. [Google Scholar]
- Levi, G.; Aloisi, F.; Ciotti, M.T.; Gallo, V. Autoradiographic localization and depolarization-induced release of acidic amino acids in differentiating cerebellar granule cell cultures. Brain Res 1984, 290, 77–86. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Name | Genbank | K25 | K5 | K5 + GIP | Forward primer | Reverse primer |
|---|---|---|---|---|---|---|
| Early growth response protein 1 (Egr1) | U75397 | −0.94 | 0.11 | 1.58 | 5′-GTTGGAATGCTGTGGTTACC-3′ | 5′-GCCAAACAAGTCACTTTGTTTA-3′ |
| 1475 ± 85 | 3019 ± 108 | 3361 ± 102 | ||||
| NIPA-like domain containing 2 (Nipal2) | NM_001130559 | −1.26 | −0.22 | 0.71 | 5′-ACATGGAGAAGCAACCTCTG-3′ | 5′-CTCCGTAATTGTCAGCAGCT-3′ |
| 667 ± 16 | 2011 ± 91 | 4065 ± 133 | ||||
| Family with sequence similarity 171, member A2 (Fam171a2) | XM_001081512 | 0.11 | −0.63 | −1.58 | 5′-AGGACAACGTGTACCGCAAT-3′ | 5′-TGGGGATCAGGTTGAGGGAA-3′ |
| 2877 ± 128 | 1136 ± 92 | 871 ± 65 | ||||
| DEAD (Asp-Glu-Ala-Asp) box helicase 56 (Ddx56) | NM_0010042112 | 0.35 | −0.53 | 0.18 | 5′-TCTTAGGCTGTCACCGACTT-3′ | 5′-ATTAGCCACTCTCACATCGC-3′ |
| 2493 ± 106 | 163 ± 12 | 2166 ± 77 | ||||
| Zinc finger protein 423 (Zfp423) | XM_001081512 | −1.23 | −0.27 | 0.59 | 5′-GAAGACAGGAACAGCGTGAC-3′ | 5′-GTCGTCATCACCATCTCCAG-3′ |
| 277 ± 31 | 856 ± 35 | 3184 ± 69 | ||||
| Neuronalpentraxin I (Nptx1) | NM_153735 | −1.50 | −0.39 | 1.14 | 5′-GGAGCTGAATGGTTACATGG-3′ | 5′-ATAAGTCCACTGCGCACAGA-3′ |
| 781 ± 32 | 2630 ± 85 | 4502 ± 181 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maino, B.; Ciotti, M.T.; Calissano, P.; Cavallaro, S. Transcriptional Analysis of Apoptotic Cerebellar Granule Neurons Following Rescue by Gastric Inhibitory Polypeptide. Int. J. Mol. Sci. 2014, 15, 5596-5622. https://doi.org/10.3390/ijms15045596
Maino B, Ciotti MT, Calissano P, Cavallaro S. Transcriptional Analysis of Apoptotic Cerebellar Granule Neurons Following Rescue by Gastric Inhibitory Polypeptide. International Journal of Molecular Sciences. 2014; 15(4):5596-5622. https://doi.org/10.3390/ijms15045596
Chicago/Turabian StyleMaino, Barbara, Maria Teresa Ciotti, Pietro Calissano, and Sebastiano Cavallaro. 2014. "Transcriptional Analysis of Apoptotic Cerebellar Granule Neurons Following Rescue by Gastric Inhibitory Polypeptide" International Journal of Molecular Sciences 15, no. 4: 5596-5622. https://doi.org/10.3390/ijms15045596
APA StyleMaino, B., Ciotti, M. T., Calissano, P., & Cavallaro, S. (2014). Transcriptional Analysis of Apoptotic Cerebellar Granule Neurons Following Rescue by Gastric Inhibitory Polypeptide. International Journal of Molecular Sciences, 15(4), 5596-5622. https://doi.org/10.3390/ijms15045596