Correlation between BPI Gene Upstream CpG Island Methylation and mRNA Expression in Piglets


Abstract
:1. Introduction
2. Results and Discussion
2.1. Bioinformatic Analysis


{kind=link}
{kind=link}
{kind=link}
| Matrix Family | Detailed Matrix Information | Start Position | End Position | Matrix Similarity | |||||
|---|---|---|---|---|---|---|---|---|---|
| V$AP2F | Transcription factor AP-2, beta | 26 | 40 | 0.901 | |||||
| V$NFKB | c-Rel | 40 | 54 | 0.910 | |||||
| V$BCDF | Cone-rod homeobox-containing transcription factor/otx-like homeobox gene | 60 | 76 | 0.976 | |||||
| V$HBOX | Homeodomain transcription factor Gsh-2 | 59 | 77 | 0.957 | |||||
| V$XBBF | Regulatory factor X, 5 | 93 | 113 | 0.946 | |||||
| V$XBBF | Regulatory factor X, 4 | 140 | 160 | 0.922 | |||||
| V$PAX3 | Pax-3 paired domain protein | 142 | 160 | 0.943 | |||||
| V$SF1F | SF1 steroidogenic factor 1 | 174 | 188 | 0.996 | |||||
| V$NBRE | Monomers of the nur subfamily of nuclear receptors (nur77, nurr1, nor-1) | 175 | 189 | 0.941 | |||||
| V$RORA | RAR (Retinoic acid receptor)-related orphan receptor alpha1 | 172 | 194 | 0.932 | |||||
| V$CSEN | Downstream regulatory element-antagonist modulator, Ca2+-binding protein of the neuronal calcium sensors family that binds DRE (downstream regulatory element) sites as a tetramer | 180 | 190 | 0.992 | |||||
| V$MEF3 | MEF3 (Myocyte enhancer factor 3) binding site, present in skeletal muscle-specific transcriptional enhancers | 180 | 192 | 0.912 |
2.2. Validation of the CpG Island Fragment Amplification

2.3. Results and Analysis of Methylation Levels
| Methylation Level (%) | 8-Day | 18-Day | 30-Day | 35-Day |
|---|---|---|---|---|
| Overall | 75.94 ± 26.75 a | 61.41 ± 35.14 a,b | 36.04 ± 10.28 b | 61.48 ± 23.22 a,b |
| CpG_1 | 71.43 ± 28.09 | 69.73 ± 16.75 | 42.30 ± 10.44 | 60.13 ± 34.29 |
| CpG_2 | 75.55 ± 36.80 | 62.98 ± 32.53 | 31.90 ± 14.15 | 49.58 ± 33.77 |
| CpG_3 | 85.00 ± 19.15 a | 67.93 ± 26.02 a,b | 27.95 ± 18.85 b | 58.85 ± 24.54 a,b |
| CpG_4 | 87.58 ± 11.36 a | 79.00 ± 16.93 a | 41.18 ± 15.92 b | 73.70 ± 11.09 a |
| CpG_5 | 67.13 ± 35.41 | 60.00 ± 33.91 | 25.00 ± 15.23 | 55.30 ± 25.86 |
| CpG_6 | 79.23 ± 26.22 | 57.80 ± 30.84 | 44.45 ± 18.99 | 67.85 ± 23.69 |
| CpG_7 | 65.38 ± 14.05 | 61.10 ± 27.67 | 36.40 ± 22.31 | 63.93 ± 9.54 |
| CpG_8 | 78.15 ± 25.64 | 55.33 ± 43.18 | 47.73 ± 14.08 | 74.60 ± 25.62 |
| CpG_9 | 73.98 ± 33.66 | 57.85 ± 35.36 | 51.13 ± 5.11 | 62.03 ± 22.05 |
| CpG_10 | 68.58 ± 27.33 a | 44.58 ± 18.04 a | 4.18 ± 8.35 b | 50.18 ± 24.72 a |
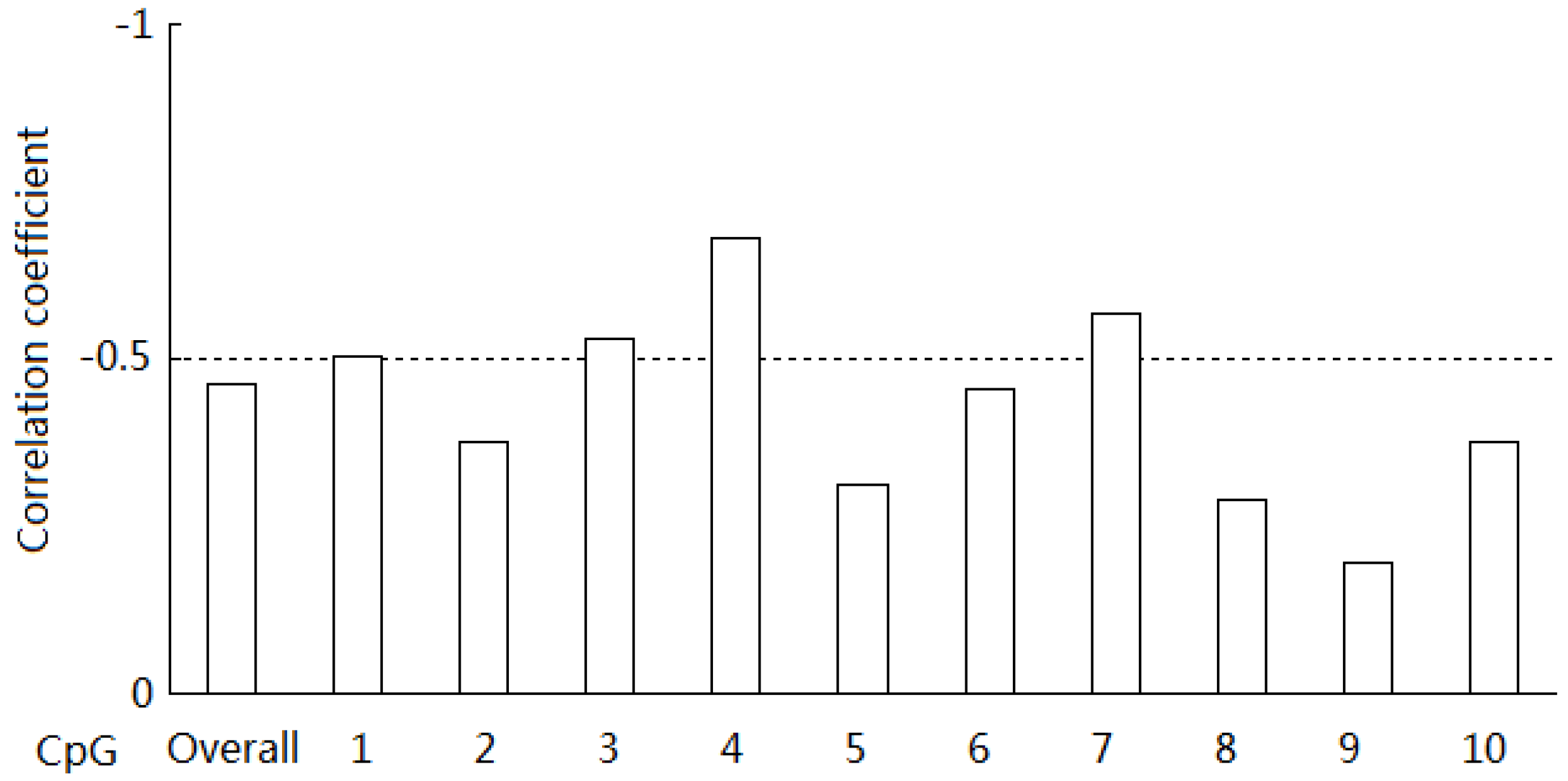
2.4. Correlation between the Methylation Levels and mRNA Expression

2.5. Discussion
3. Experimental Section
3.1. Experimental Animals
3.2. Bioinformatic Analysis
3.3. Methylation Analysis
3.4. Real-Time PCR Analysis
| Primer | Sequence of Primer | Length (bp) |
|---|---|---|
| BPI RT-PCR primer | 5'-ATATCGAATCTGCGCTCCGA-3' | 136 |
| 5'-TTGATGCCAACCATTCTGTCC-3' | ||
| GAPDH RT-PCR primer | 5'-ACATCATCCCTGCTTCTACTGG-3' | 187 |
| 5'-CTCGGACGCCTGCTTCAC-3' |
3.5. Data Processing and Analysis
4. Conclusions
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Balakrishnan, A.; Marathe, S.A.; Joglekar, M.; Chakravortty, D. Bactericidal/permeability increasing protein: A multifaceted protein with functions beyond LPS neutralization. Innate Immun. 2013, 19, 339–347. [Google Scholar] [CrossRef]
- Schultz, H.; Weiss, J.P. The bactericidal/permeability-increasing protein (BPI) in infection and inflammatory disease. Clin. Chim. Acta 2007, 384, 12–23. [Google Scholar] [CrossRef]
- Mao, Y.Z.; Zhou, C.Y.; Zhu, L.; Huang, Y.; Yan, T.R.; Fang, J.G.; Zhu, W. Identification and expression analysis on bactericidal permeability-increasing protein (BPI)/lipopolysaccharide-binding protein (LBP) of ark shell, Scapharca broughtonii. Fish Shellfish Immunol. 2013, 35, 642–652. [Google Scholar] [CrossRef]
- Christopher, K.T.; Thomas, J.S.; Shi, X.W.; Martha, A.M. Genetic Markers for Improved Disease Resistance in Animals (BPI). U.S. Patent 20040234980, 25 November 2004. [Google Scholar]
- Imberechts, H.; Wild, P.; Charlier, G.; de Greve, H.; Lintermans, P.; Pohl, P. Characterization of F18 fimbrial genes fedE and fedF involved in adhesion and length of enterotoxemic Escherichia coli strain 107/86. Microb. Pathog. 1996, 21, 183–192. [Google Scholar]
- Imberechts, H.; de Greve, H.; Schlicker, C.; Bouchet, H.; Pohl, P.; Charlier, G.; Bertschinger, H.; Wild, P.; Vandekerckhove, J.; van Damme, J. Characterization of F107 fimbriae of Escherichia coli 107/86, which causes oedema disease in pigs and nucleotide sequence of F107 major fimbrial sub−unit gene, fedA. Infect. Immun. 1992, 60, 1963–1971. [Google Scholar]
- Zhu, J.; Zi, C.; Wu, Z.C.; Zheng, X.R.; Su, X.M.; Zhu, G.Q.; Huang, X.G.; Wu, S.L.; Bao, W.B. Age-dependent expression of the BPI gene in Sutai piglets. Genet. Mol. Res. 2013, 12, 2120–2126. [Google Scholar] [CrossRef]
- Wang, R.X.; Xu, J.H. Genomic DNA methylation and histone methylation (in Chinese). Yi Chuan 2014, 36, 191–199. [Google Scholar]
- Cartharius, K.; Frech, K.; Grote, K.; Klocke, B.; Haltmeier, M.; Klingenhoff, A.; Frisch, M.; Bayerlein, M.; Werner, T. MatInspector and beyond: Promoter analysis based on transcription factor binding sites. Bioinformatics 2005, 21, 2933–2442. [Google Scholar] [CrossRef]
- Kang, Y.K.; Park, J.S.; Koo, D.B.; Choi, Y.H.; Kim, S.U.; Lee, K.K.; Han, Y.M. Limited demethylation leaves mosaictype methylation states in cloned bovine preimplantation embryos. EMBO J. 2002, 21, 1092–1100. [Google Scholar] [CrossRef]
- Gardiner-Garden, M.; Frommer, M. CpG islands in vertebrate genomes. J. Mol. Biol. 1987, 196, 261–282. [Google Scholar] [CrossRef]
- Antequera, F. Structure, function and evolution of CpG island promoters. Cell. Mol. Life Sci. 2003, 60, 1647–1658. [Google Scholar] [CrossRef]
- Laird, P.W. Principles and challenges of genomewide DNA methylation analysis. Nat. Rev. Genet. 2010, 11, 191–203. [Google Scholar] [CrossRef]
- Wang, J.; Zhuang, J.; Iyer, S.; Lin, X.; Whitfield, T.W.; Greven, M.C.; Pierce, B.G.; Dong, X.; Kundaje, A.; Cheng, Y.; et al. Sequence features and chromatin structure around the genomic regions bound by 119 human transcription factors. Genome Res. 2012, 22, 1798–812. [Google Scholar] [CrossRef]
- Baker-Andresen, D.; Ratnu, V.S.; Bredy, T.W. Dynamic DNA methylation: A prime candidate for genomic metaplasticity and behavioral adaptation. Trends Neurosci. 2013, 36, 3–13. [Google Scholar]
- Laurent, L.; Wong, E.; Li, G.; Huynh, T.; Tsirigos, A.; Ong, C.T.; Low, H.M.; Kin Sung, K.W.; Rigoutsos, I.; Loring, J.; et al. Dynamic changes in the human methylome during differentiation. Genome Res. 2010, 20, 320–331. [Google Scholar] [CrossRef]
- Thurston, A.; Lucas, E.S.; Allegrucci, C.; Steele, W.; Young, L.E. Region-specific DNA methylation in the preimplantation embryo as a target for genomic plasticity. Theriogenology. 2007, 68, S98–S106. [Google Scholar] [CrossRef]
- Haaf, T. Methylation dynamics in the early mammalian embryo: Implications of genome reprogramming defects for development. Curr. Top. Microbiol. Immunol. 2006, 310, 13–22. [Google Scholar]
- Calvanese, V.; Lara, E.; Kahn, A.; Fraga, M.F. The role of epigenetics in aging and age-related diseases. Ageing Res. Rev. 2009, 8, 268–276. [Google Scholar] [CrossRef]
- Bjornsson, H.T.; Sigurdsson, M.I.; Fallin, M.D.; Irizarry, R.A.; Aspelund, T.; Cui, H.; Yu, W.; Rongione, M.A.; Ekström, T.J.; Harris, T.B.; et al. Intra-individual change over time in DNA methylation with familial clustering. JAMA 2008, 299, 2877–2883. [Google Scholar]
- Heyn, H.; Li, N.; Ferreira, H.J.; Moran, S.; Pisano, D.G.; Gomez, A.; Diez, J.; Sanchez-Mut, J.V.; Setien, F.; Carmona, F.J.; et al. Distinct DNA methylomes of newborns and centenarians. Proc. Natl. Acad. Sci. USA 2012, 109, 10522–10527. [Google Scholar] [CrossRef]
- Akin, H.; Tahan, G.; Türe, F.; Eren, F.; Atuğ, O.; Tahan, V.; Hamzaoğlu, I.; Imeryüz, N.; Tözün, N.; Hamzaoglu, H.O. Association between bactericidal/permeability increasing protein (BPI) gene polymorphism (Lys216Glu) and in flammatory bowel disease. J. Crohns Colitis 2011, 5, 14–18. [Google Scholar] [CrossRef]
- Verdonck, F.; Cox, E.; van Gog, K.; van der Stede, Y.; Duchateau, L.; Deprez, P.; Goddeeris, B.M. Different kinetic of antibody responses following infection of newly weaned pigs with an F4 enterotoxigenic Escherichia coli strain or an F18 verotoxigenic Escherichia coli strain. Vaccine 2002, 20, 2995–3004. [Google Scholar] [CrossRef]
- Barrera, V.; Peinado, M.A. Evaluation of single CpG sites as proxies of CpG island methylation states at the genome scale. Nucleic Acids Res. 2012, 40, 11490–11498. [Google Scholar] [CrossRef]
- Xie, X.L.; Yu, Y.; Yuan, Z.F.; Yang, J.; Ma, P.P.; Li, D.C.; Yu, S.K.; An, F.; Feng, X.J.; Zhang, Y. Comparative analysis on content and distribution of CpG sites in milk production traits and mastitis-related genes in dairy cattle (in Chinese). Yi Chuan 2012, 34, 437–444. [Google Scholar] [CrossRef]
- MethPrimer. Available online: http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi (accessed on 15 Novmber 2013).
- MatInspector. Available online: http://www.genomatix.de/products/MatInspector/index.html (accessed on 15 Novmber 2013).
- QUMA. Available online: http://quma.cdb.riken.jp (accessed on 20 March 2014).
- Shaw, A.E.; Reid, S.M.; Ebert, K.; Hutchings, G.H.; Ferris, N.P.; King, D.P. Implementation of a one-step real-time RT-PCR protocol for diagnosis of foot-and-mouth disease. Virol. Methods 2007, 143, 81–85. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, J.; Yin, X.; Sun, L.; Sun, S.; Zi, C.; Zhu, G.; Wu, S.; Bao, W. Correlation between BPI Gene Upstream CpG Island Methylation and mRNA Expression in Piglets. Int. J. Mol. Sci. 2014, 15, 10989-10998. https://doi.org/10.3390/ijms150610989
Wang J, Yin X, Sun L, Sun S, Zi C, Zhu G, Wu S, Bao W. Correlation between BPI Gene Upstream CpG Island Methylation and mRNA Expression in Piglets. International Journal of Molecular Sciences. 2014; 15(6):10989-10998. https://doi.org/10.3390/ijms150610989
Chicago/Turabian StyleWang, Jing, Xuemei Yin, Li Sun, Shouyong Sun, Chen Zi, Guoqiang Zhu, Shenglong Wu, and Wenbin Bao. 2014. "Correlation between BPI Gene Upstream CpG Island Methylation and mRNA Expression in Piglets" International Journal of Molecular Sciences 15, no. 6: 10989-10998. https://doi.org/10.3390/ijms150610989
APA StyleWang, J., Yin, X., Sun, L., Sun, S., Zi, C., Zhu, G., Wu, S., & Bao, W. (2014). Correlation between BPI Gene Upstream CpG Island Methylation and mRNA Expression in Piglets. International Journal of Molecular Sciences, 15(6), 10989-10998. https://doi.org/10.3390/ijms150610989