Modified Low Density Lipoprotein and Lipoprotein-Containing Circulating Immune Complexes as Diagnostic and Prognostic Biomarkers of Atherosclerosis and Type 1 Diabetes Macrovascular Disease

 ,
,  ,
,
Abstract
:1. Introduction
2. Low Density Lipoprotein
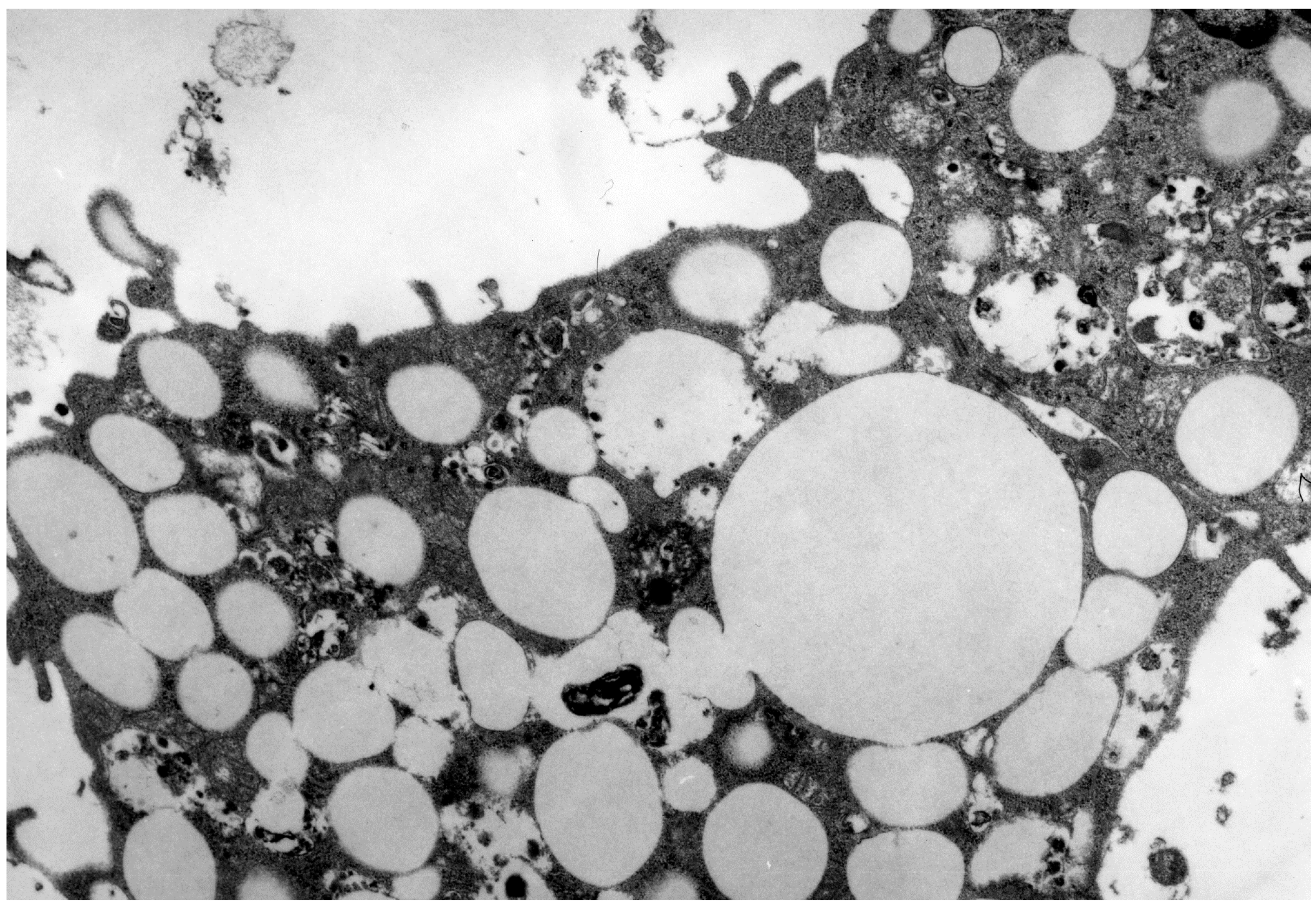
3. LDL in Atherosclerosis
4. LDL Modifications
4.1. Oxidized LDL
4.2. Malondialdehide LDL
4.3. Glycated LDL
4.4. Carbamylated LDL
4.5. Desialylated LDL
5. Naturally Occurring Modified Forms of LDL
6. Autoantibodies against Modified LDL
6.1. Circulating Anti-LDL Antibodies

{kind=link}
{kind=link}
{kind=link}
| LDL Preparation | Affinity Constant (×10−7·М−1) |
|---|---|
| LDL from healthy subjects | 2.4 |
| glycosylated LDL | 2.6 |
| acetylated LDL | 2.8 |
| Cu2+-oxidized LDL | 3.5 |
| LDL from atherosclerotic patients | 11.3 |
| MDA-LDL | 10.9 |
| Desialylated LDL | 89.4 |
6.2. Diagnostic and Prognostic Value of Anti-LDL Self-Antibodies
7. LDL-Containing Circulating Immune Complexes
7.1. Atherogenic and Proinflammatory Properties of LDL-Containing Immune Complexes

7.2. Diagnostic and Prognostic Value of LDL-Containing Immune Complexes

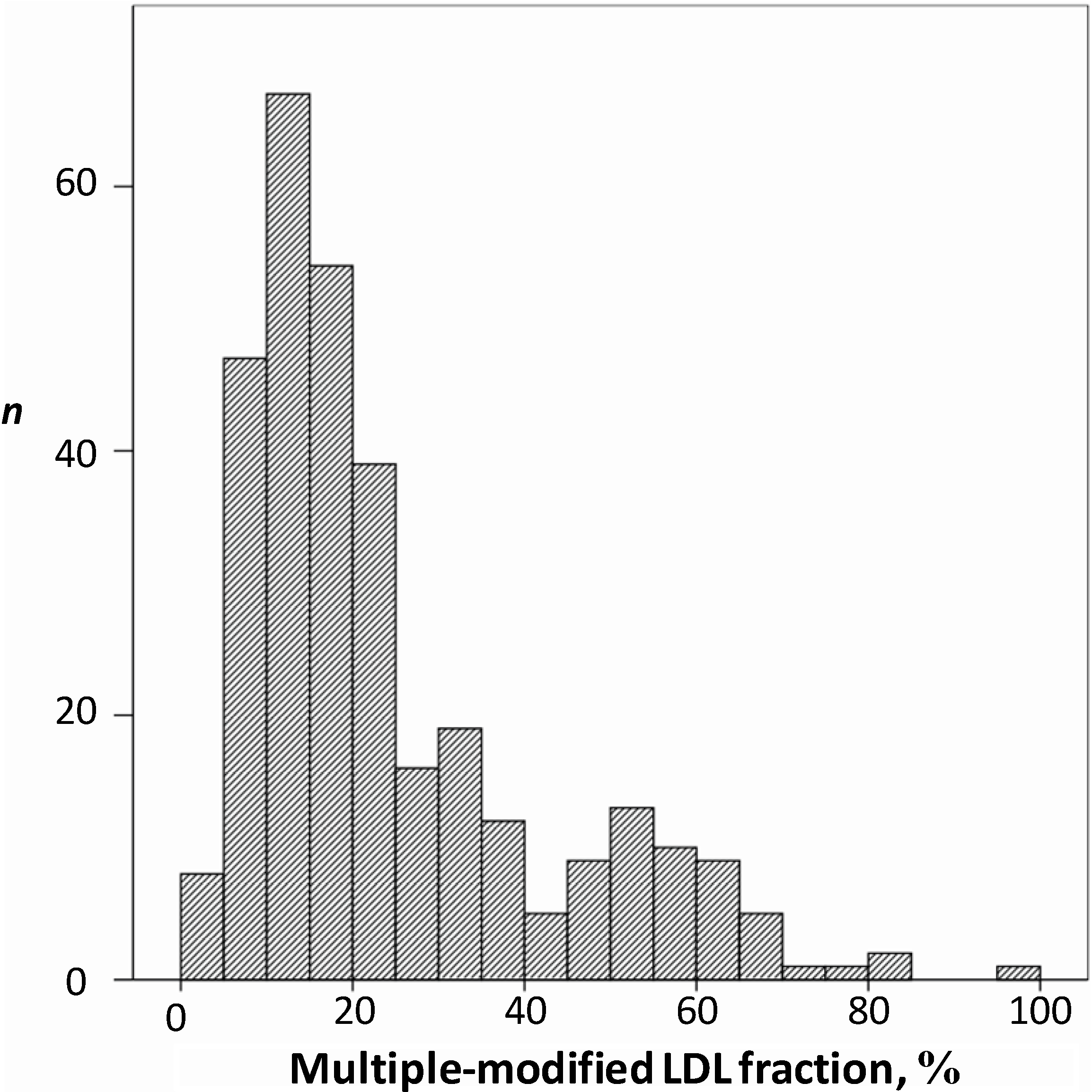
7.3. Diagnostic and Prognostic Potential of Multiple Modified LDL

8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schwartz, S.M.; deBlois, D.; O’Brien, E.R. The intima—Soil for atherosclerosis and restenosis. Circ. Res. 1995, 77, 445–465. [Google Scholar] [CrossRef]
- Andreeva, E.R.; Pugach, I.M.; Orekhov, A.N. Collagen-synthesizing cells in initial and advanced atherosclerotic lesions of human aorta. Atherosclerosis 1997, 130, 133–142. [Google Scholar] [CrossRef]
- Ryu, C.W.; Kwak, H.S.; Jahng, G.H.; Lee, H.N. High-resolution MRI of intracranial atherosclerotic disease. Neurointervention 2014, 9, 9–20. [Google Scholar] [CrossRef]
- Michel, J.B.; Martin-Ventura, J.L.; Nicoletti, A.; Ho-Tin-Noé, B. Pathology of human plaque vulnerability: Mechanisms and consequences of intraplaque haemorrhages. Atherosclerosis 2014, 234, 311–319. [Google Scholar] [CrossRef]
- Watts, G.F.; Gidding, S.; Wierzbicki, A.S.; Toth, P.P.; Alonso, R.; Brown, W.V.; Bruckert, E.; Defesche, J.; Lin, K.K.; Livingston, M.; et al. Integrated guidance on the care of familial hypercholesterolaemia from the International FH Foundation. Int. J. Cardiol. 2014, 171, 309–325. [Google Scholar] [CrossRef]
- Noyes, A.M.; Thompson, P.D. A systematic review of the time course of atherosclerotic plaque regression. Atherosclerosis 2014, 234, 75–84. [Google Scholar] [CrossRef]
- Afonso, L.; Veeranna, V.; Zalawadiya, S.; Ramesh, K.; Niraj, A.; Panaich, S. Predictors of residual cardiovascular risk in patients on statin therapy for primary prevention. Cardiology 2011, 119, 187–190. [Google Scholar] [CrossRef]
- Bedi, U.S.; Singh, M.; Singh, P.P.; Bhuriya, R.; Bahekar, A.; Molnar, J.; Khosla, S.; Arora, R. Effects of statins on progression of carotid atherosclerosis as measured by carotid intimal-medial thickness: A meta-analysis of randomized controlled trials. J. Cardiovasc. Pharmacol. Ther. 2010, 15, 268–273. [Google Scholar] [CrossRef]
- Whayne, T.F., Jr. Problems and possible solutions for therapy with statins. Int. J. Angiol. 2013, 22, 75–82. [Google Scholar] [CrossRef]
- Di Angelantonio, E.; Gao, P.; Pennells, L.; Kaptoge, S.; Caslake, M.; Thompson, A.; Butterworth, A.S.; Sarwar, N.; Wormser, D.; Saleheen, D.; et al. Lipid-related markers and cardiovascular disease prediction. JAMA 2012, 307, 2499–2506. [Google Scholar]
- Benitez, R.M.; Vogel, R.A. Assessment of subclinical atherosclerosis and cardiovascular risk. Clin. Cardiol. 2001, 24, 642–650. [Google Scholar] [CrossRef]
- Belcaro, G.; Nicolaides, A.N.; Ramaswami, G.; Cesarone, M.R.; de Sanctis, M.; Incandela, L.; Ferrari, P.; Geroulakos, G.; Barsotti, A.; Griffin, M.; et al. Carotid and femoral ultrasound morphology screening and cardiovascular events in low risk subjects: A 10-year follow-up study (the CAFES-CAVE study). Atherosclerosis 2001, 156, 379–387. [Google Scholar] [CrossRef]
- Grewal, J.; Anand, S.; Islam, S.; Lonn, E. Prevalence and predictors of subclinical atherosclerosis among asymptomatic “low risk” individuals in a multiethnic population. Atherosclerosis 2008, 197, 435–442. [Google Scholar] [CrossRef]
- Castellon, X.; Bogdanova, V. Screening for subclinical atherosclerosis by noninvasive methods in asymptomatic patients with risk factors. Clin. Interv. Aging 2013, 8, 573–580. [Google Scholar] [CrossRef]
- Bobryshev, Y.V. Novel targets and biomarkers for anti-atherosclerotic therapy. Curr. Pharm. Des. 2013, 19, 5837–5839. [Google Scholar] [CrossRef]
- Ross, R. Cell biology of atherosclerosis. Annu. Rev. Physiol. 1995, 57, 791–804. [Google Scholar] [CrossRef]
- Tertov, V.V.; Bittolo-Bon, G.; Sobenin, I.A.; Cazzolato, G.; Orekhov, A.N.; Avogaro, P. Naturally occurring modified low density lipoproteins are similar if not identical: More electronegative and desialylated lipoprotein subfractions. Exp. Mol. Pathol. 1995, 62, 166–172. [Google Scholar] [CrossRef]
- Pentikäinen, M.O.; Oörni, K.; Ala-Korpela, M.; Kovanen, P.T. Modified LDL–trigger of atherosclerosis and inflammation in the arterial intima. J. Intern. Med. 2000, 247, 359–370. [Google Scholar] [CrossRef]
- Mahley, R.W.; Innerarity, T.L.; Rall, S.C., Jr.; Weisgraber, K.H. Plasma lipoproteins: Apolipoprotein structure and function. J. Lipid Res. 1984, 25, 1277–1294. [Google Scholar]
- Cho, N.H.; Seong, S.Y. Apolipoproteins inhibit the innate immunity activated by necrotic cells or bacterial endotoxin. Immunology 2009, 128, e479–e586. [Google Scholar] [CrossRef]
- Fisher, W.R. The structure of the lower-density lipoproteins of human plasma: Newer concepts derived from studies with the analytical ultracentrifuge. Ann. Clin. Lab. Sci. 1972, 2, 198–208. [Google Scholar]
- Williams, P.T.; Vranizan, K.M.; Krauss, R.M. Correlations of plasma lipoproteins with LDL subfractions by particle size in men and women. J. Lipid Res. 1992, 33, 765–774. [Google Scholar]
- Brown, M.S.; Kovanen, P.T.; Goldstein, J.L. Regulation of plasma cholesterol by lipoprotein receptors. Science 1981, 212, 628–635. [Google Scholar]
- Hegele, R.A. Plasma lipoproteins: Genetic influences and clinical implications. Nat. Rev. Genet. 2009, 10, 109–121. [Google Scholar] [CrossRef]
- Hevonoja, T.; Pentikäinen, M.O.; Hyvonen, M.T.; Kovanen, P.T.; Ala-Korpela, M. Structure of low density lipoprotein (LDL) particles: Basis for understanding molecular changes in modified LDL. Biochim. Biophys. Acta 2000, 1488, 189–210. [Google Scholar] [CrossRef]
- Segrest, J.P.; Jones, M.K.; de Loof, H.; Dashti, N. Structure of apolipoprotein B-100 in low density lipoproteins. J. Lipid Res. 2001, 42, 1346–1367. [Google Scholar]
- Kane, J.P. Apoprotein B: Structural and metabolic heterogeneity. Annu. Rev. Physiol. 1983, 45, 637–650. [Google Scholar] [CrossRef]
- Yang, C.Y.; Gu, Z.W.; Weng, S.A.; Kim, T.W.; Chen, S.H.; Pownall, H.J.; Sharp, P.M.; Liu, S.W.; Li, W.H.; Gotto, A.M., Jr. Structure of apolipoprotein B-100 of human low density lipoproteins. Arteriosclerosis 1989, 9, 96–108. [Google Scholar] [CrossRef]
- Segrest, J.P.; Jones, M.K.; Mishra, V.K.; Anantharamaiah, G.M.; Garber, D.W. ApoB-100 has a pentapartite structure composed of three amphipathic alpha-helical domains alternating with two amphipathic beta-strand domains. Detection by the computer program LOCATE. Arterioscler. Thromb. 1994, 14, 1674–1685. [Google Scholar] [CrossRef]
- Segrest, J.P.; Jones, M.K.; Mishra, V.K.; Pierotti, V.; Young, S.H.; Boren, J.; Innerarity, T.L.; Dashti, N. Apolipoprotein B-100: Conservation of lipid-associating amphipathic secondary structural motifs in nine species of vertebrates. J. Lipid Res. 1998, 39, 85–102. [Google Scholar]
- Powell, L.M.; Wallis, S.C.; Pease, R.J.; Edwards, Y.H.; Knott, T.J.; Scott, J. A novel form of tissue-specific RNA processing produces apolipoprotein-B48 in intestine. Cell 1987, 50, 831–840. [Google Scholar] [CrossRef]
- Kumar, V.; Butcher, S.J.; Öörni, K.; Engelhardt, P.; Heikkonen, J.; Kaski, K.; Ala-Korpela, M.; Kovanen, P.T. Three-dimensional cryoEM reconstruction of native LDL particles to 16Å resolution at physiological body temperature. PLoS One 2011, 6, e18841. [Google Scholar] [CrossRef]
- Forgez, P.; Gregory, H.; Young, J.A.; Knott, T.; Scott, J.; Chapman, M.J. Identification of surface-exposed segments of apolipoprotein B-100 in the LDL particle. Biochem. Biophys. Res. Commun. 1986, 140, 250–257. [Google Scholar] [CrossRef]
- Segrest, J.P.; Jones, M.K.; Dashti, N. N-terminal domain of apolipoprotein B has structural homology to lipovitellin and microsomal triglyceride transfer protein: A “lipid pocket” model for self-assembly of apob-containing lipoprotein particles. J. Lipid Res. 1999, 40, 1401–1416. [Google Scholar]
- Schneider, W.J. The low density lipoprotein receptor. Biochim. Biophys. Acta 1989, 988, 303–317. [Google Scholar] [CrossRef]
- Bobryshev, Y.V. Monocyte recruitment and foam cell formation in atherosclerosis. Micron 2006, 37, 208–222. [Google Scholar] [CrossRef]
- Rezaee, F.; Casetta, B.; Levels, J.H.; Speijer, D.; Meijers, J.C. Proteomic analysis of high-density lipoprotein. Proteomics 2006, 6, 721–730. [Google Scholar] [CrossRef]
- Queiroz, K.C.; Tio, R.A.; Zeebregts, C.J.; de Vries, M.; Mahmoudi, M.; Peppelenbosch, M.P.; Rezaee, F. Human plasma very low density lipoprotein carries Indian hedgehog. J. Proteome Res. 2010, 9, 6052–6059. [Google Scholar] [CrossRef]
- Dashty, M.; Motazacker, M.M.; Levels, J.; de Vries, M.; Mahmoudi, M.; Peppelenbosch, M.P.; Rezaee, F. Proteome of human plasma very low-density lipoprotein and low-density lipoprotein exhibits a link with coagulation and lipid metabolism. Thromb. Haemost. 2014, 111, 518–530. [Google Scholar] [CrossRef]
- Yuasa, Y.; Osaki, T.; Makino, H.; Iwamoto, N.; Kishimoto, I.; Usami, M.; Minamino, N.; Harada-Shiba, M. Proteomic analysis of proteins eliminated by low-density lipoprotein apheresis. Ther. Apher. Dial. 2014, 18, 93–102. [Google Scholar] [CrossRef]
- Karlsson, H.; Leanderson, P.; Tagesson, C.; Lindahl, M. Lipoproteomics I: Mapping of proteins in low-density lipoprotein using two-dimensional gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 551–565. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Nieddu, G.; Zinellu, E.; de Muro, P.; Piredda, F.; Guarino, A.; Spirito, R.; Carta, F.; Turrini, F.; Formato, M. Proteomic analysis of plasma-purified VLDL, LDL, and HDL fractions from atherosclerotic patients undergoing carotid endarterectomy: Identification of serum amyloid A as a potential marker. Oxid. Med. Cell Longev. 2013, 2013, 385214. [Google Scholar]
- Dong, Z.; Wu, T.; Qin, W.; An, C.; Wang, Z.; Zhang, M.; Zhang, Y.; Zhang, C.; An, F. Serum amyloid a directly accelerates the progression of atherosclerosis in apolipoprotein E-deficient mice. Mol. Med. 2011, 17, 1357–1364. [Google Scholar]
- Song, C.; Hsu, K.; Yamen, E.; Yan, W.; Fock, J.; Witting, P.K.; Geczy, C.L.; Freedman, S.B. Serum amyloid A induction of cytokines in monocytes/macrophages and lymphocytes. Atherosclerosis 2009, 207, 374–383. [Google Scholar] [CrossRef]
- Prassl, R.; Laggner, P. Lipoprotein structure and dynamics: Low density lipoprotein viewed as a highly dynamic and flexible nanoparticle. InTech 2012. [Google Scholar] [CrossRef]
- Bancells, C.; Benítez, S.; Villegas, S.; Jorba, O.; Ordóñez-Llanos, J.; Sánchez-Quesada, J.L. Novel phospholipolytic activities associated with electronegative low-density lipoprotein are involved in increased self-aggregation. Biochemistry 2008, 47, 8186–8194. [Google Scholar] [CrossRef]
- Sweet, M.J.; Hume, D.A. Endotoxin signal transduction in macrophages. J. Leukoc. Biol. 1996, 60, 8–26. [Google Scholar]
- Liu, Y.H.; Atkinson, D. Enhancing the contrast of ApoB to locate the surface components in the 3D density map of human LDL. J. Mol. Biol. 2011, 405, 274–283. [Google Scholar] [CrossRef]
- Deckelbaum, R.J.; Shipley, G.G.; Small, D.M.; Lees, R.S.; George, P.K. Thermal transitions in human plasma low density lipoproteins. Science 1975, 190, 392–394. [Google Scholar] [CrossRef]
- Deckelbaum, R.J.; Shipley, G.G.; Small, D.M. Structure and interactions of lipids in human plasma low density lipoproteins. J. Biol. Chem. 1977, 252, 744–754. [Google Scholar]
- Reis, A.; Rudnitskaya, A.; Blackburn, G.J.; Mohd Fauzi, N.; Pitt, A.R.; Spickett, C.M. A comparison of five lipid extraction solvent systems for lipidomic studies of human LDL. J. Lipid Res. 2013, 54, 1812–1824. [Google Scholar] [CrossRef]
- Kanno, K.; Wu, M.K.; Agate, D.A.; Fanelli, B.K.; Wagle, N.; Scapa, E.F.; Ukomadu, C.; Cohen, D.E. Interacting proteins dictate function of the minimal START domain phosphatidylcholine transfer protein/StarD2. J. Biol. Chem. 2007, 282, 30728–30736. [Google Scholar]
- Yao, Z.M.; Vance, D.E. The active synthesis of phosphatidylcholine is required for very low density lipoprotein secretion from rat hepatocytes. J. Biol. Chem. 1988, 263, 2998–3004. [Google Scholar]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Serrano, D.; Bhowmick, T.; Chadha, R.; Garnacho, C.; Muro, S. Intercellular adhesion molecule 1 engagement modulates sphingomyelinase and ceramide, supporting uptake of drug carriers by the vascular endothelium. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1178–1185. [Google Scholar] [CrossRef]
- Merrill, A.H., Jr.; Lingrell, S.; Wang, E.; Nikolova-Karakashian, M.; Vales, T.R.; Vance, D.E. Sphingolipid biosynthesis de novo by rat hepatocytes in culture. Ceramide and sphingomyelin are associated with, but not required for, very-low density lipoprotein (VLDL) secretion. J. Biol. Chem. 1995, 270, 13834–13841. [Google Scholar]
- Nikolova-Karakashian, M.N. Ceramide in serum lipoproteins: Function and regulation of metabolism. Ceramide Signaling; Futerman, A.H., Ed.; Kluwer Academic/Plenum: Dordrecht, The Netherlands, 2002. Available online: http://www.landesbioscience.com/pdf/Futerman15nikolova.pdf (accessed on 26 June 2014).
- Hannun, Y.A.; Luberto, C.; Argraves, K.M. Enzymes of sphingolipid metabolism: From modular to integrative signaling. Biochemistry 2001, 40, 4893–4903. [Google Scholar] [CrossRef]
- Schissel, S.L.; Keesler, G.A.; Schuchman, E.H.; Williams, K.J.; Tabas, I. The cellular trafficking and zinc dependence of secretory and lysosomal sphingomyelinase, two products of the acid sphingomyelinase gene. J. Biol. Chem. 1998, 273, 18250–18259. [Google Scholar]
- Marathe, S.; Kuriakose, G.; Williams, K.J.; Tabas, I. Sphingomyelinase, an enzyme implicated in atherogenesis, is present in atherosclerotic lesions and binds to specific components of the subendothelial extracellular matrix. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2648–2658. [Google Scholar] [CrossRef]
- Marathe, S.; Choi, Y.; Leventhal, A.R.; Tabas, I. Sphingomyelinase converts lipoproteins from apolipoprotein E knockout mice into potent inducers of macrophage foam cell formation. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2607–2613. [Google Scholar] [CrossRef]
- Boyanovsky, B.; Karakashian, A.; King, K.; Giltiay, N.; Nikolova-Karakashian, M. Uptake and metabolism of low density lipoproteins with elevated ceramide content by human microvascular endothelial cells: Implications for the regulation of apoptosis. J. Biol. Chem. 2003, 278, 26992–26999. [Google Scholar]
- Matsunaga, T.; Kotamraju, S.; Kalivendi, S.V.; Dhanasekaran, A.; Joseph, J.; Kalyanaraman, B. Ceramide-induced intracellular oxidant formation, iron signaling, and apoptosis in endothelial cells: Protective role of endogenous nitric oxide. J. Biol. Chem. 2004, 279, 28614–28624. [Google Scholar]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell. Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Chandru, H.; Boggaram, V. The role of sphingosine 1-phosphate in the TNF-α induction ofIL-8 gene expression in lung epithelial cells. Gene 2007, 391, 150–160. [Google Scholar] [CrossRef]
- Sánchez-Quesada, J.L.; Benítez, S.; Ordóñez-Llanos, J. Electronegative low-density lipoprotein. Curr. Opin. Lipidol. 2004, 15, 329–335. [Google Scholar] [CrossRef]
- Benítez, S.; Camacho, M.; Arcelus, R.; Vila, L.; Bancells, C.; Ordóñez-Llanos, J.; Sánchez-Quesada, J.L. Increased lysophosphatidylcholine and non-esterified fatty acid content in LDL induces chemokine release in endothelial cells. Relationship with electronegative LDL. Atherosclerosis 2004, 177, 299–305. [Google Scholar]
- Oörni, K.; Pentikäinen, M.O.; Ala-Korpela, M.; Kovanen, P.T. Aggregation, fusion, and vesicle formation of modified low density lipoprotein particles: Molecular mechanisms and effects on matrix interactions. J. Lipid Res. 2000, 41, 1703–1714. [Google Scholar]
- Hajjar, D.P.; Pomerantz, K.B. Signal transduction in atherosclerosis: Integration of cytokines and the eicosanoid network. FASEB J. 1992, 6, 2933–2941. [Google Scholar]
- Orekhov, A.N. Direct anti-atherosclerotic therapy; development of natural anti-atherosclerotic drugs preventing cellular cholesterol retention. Curr. Pharm. Des. 2013, 19, 5909–5928. [Google Scholar] [CrossRef]
- Kannel, W.B.; Castelli, W.P.; Gordon, T. Cholesterol in the prediction of atherosclerotic disease. New perspectives based on the Framingham study. Ann. Intern. Med. 1979, 90, 85–91. [Google Scholar] [CrossRef]
- Brunzell, J.D.; Davidson, M.; Furberg, C.D.; Goldberg, R.B.; Howard, B.V.; Stein, J.H.; Witztum, J.L. Lipoprotein management in patients with cardiometabolic risk. J. Am. Coll. Cardiol. 2008, 51, 1512–1524. [Google Scholar] [CrossRef]
- Ramjee, V.; Sperling, L.S.; Jacobson, T.A. Non-high-density lipoprotein cholesterol versus apolipoprotein B in cardiovascular risk stratification: Do the math. J. Am. Coll. Cardiol. 2011, 58, 457–463. [Google Scholar] [CrossRef]
- Robinson, J.G.; Wang, S.; Jacobson, T.A. Meta-analysis of comparison of effectiveness of lowering apolipoprotein B versus low-density lipoprotein cholesterol and nonhigh-density lipoprotein cholesterol for cardiovascular risk reduction in randomized trials. Am. J. Cardiol. 2012, 110, 1468–1476. [Google Scholar] [CrossRef]
- Milne, R.W.; Weech, P.K.; Marcel, Y.L. Heterogeneity of apolipoprotein B containing lipoproteins. Adv. Exp. Med. Biol. 1987, 210, 1–5. [Google Scholar] [CrossRef]
- Martin, S.S.; Qasim, A.N.; Wolfe, M.; St Clair, C.; Schwartz, S.; Iqbal, N.; Schutta, M.; Bagheri, R.; Mehta, N.N.; Rader, D.J.; et al. Comparison of high-density lipoprotein cholesterol to apolipoprotein A-I and A-II to predict coronary calcium and the effect of insulin resistance. Am. J. Cardiol. 2011, 107, 393–398. [Google Scholar] [CrossRef]
- Grundy, S.M. Use of emerging lipoprotein risk factors in assessment of cardiovascular risk. JAMA 2012, 307, 2540–2542. [Google Scholar] [CrossRef]
- Itabe, H.; Obama, T.; Kato, R. The dynamics of oxidized LDL during atherogenesis. J. Lipids 2011, 2011, 418313. [Google Scholar]
- Kim, J.A.; Territo, M.C.; Wayner, E.; Carlos, T.M.; Parhami, F.; Smith, C.W.; Haberland, M.E.; Fogelman, A.M.; Berliner, J.A. Partial characterization of leukocyte binding molecules on endothelial cells induced by minimally oxidized LDL. Arterioscler. Thromb. 1994, 14, 427–433. [Google Scholar] [CrossRef]
- Tanner, F.C.; Boulanger, C.M.; Lüscher, T.F. Endothelium-derived nitric oxide, endothelin, and platelet vessel wall interaction: Alterations in hypercholesterolemia and atherosclerosis. Semin. Thromb. Hemost. 1993, 19, 167–175. [Google Scholar] [CrossRef]
- Liao, J.K. Inhibition of Gi proteins by low density lipoprotein attenuates bradykinin-stimulated release of endothelial-derived nitric oxide. J. Biol. Chem. 1994, 269, 12987–12992. [Google Scholar]
- Singh, R.B.; Mengi, S.A.; Xu, Y.J.; Arneja, A.S.; Dhalla, N.S. Pathogenesis of atherosclerosis: A multifactorial process. Exp. Clin. Cardiol. 2002, 7, 40–53. [Google Scholar]
- Jaakkola, O.; Solakivi, T.; Tertov, V.V.; Orekhov, A.N.; Miettinen, T.A.; Nikkari, T. Characteristics of low-density lipoprotein subfractions from patients with coronary artery disease. Coron. Artery Dis. 1993, 4, 379–385. [Google Scholar] [CrossRef]
- Tertov, V.V.; Orekhov, A.N. Metabolism of native and naturally occurring multiple modified low density lipoprotein in smooth muscle cells of human aortic intima. Exp. Mol. Pathol. 1997, 64, 127–145. [Google Scholar] [CrossRef]
- Botham, K.M.; Moore, E.H.; de Pascale, C.; Bejta, F. The induction of macrophage foam cell formation by chylomicron remnants. Biochem. Soc. Trans. 2007, 35, 454–458. [Google Scholar] [CrossRef]
- Marchant, C.E.; van der Veen, C.; Law, N.S.; Hardwick, S.J.; Carpenter, K.L.; Mitchinson, M.J. Oxidation of low-density lipoprotein by human monocyte-macrophages results in toxicity to the oxidising culture. Free Radic. Res. 1996, 24, 333–342. [Google Scholar] [CrossRef]
- Tsimikas, S.; Miller, Y.I. Oxidative modification of lipoproteins: Mechanisms, role in inflammation and potential clinical applications in cardiovascular disease. Curr. Pharm. Des. 2011, 17, 27–37. [Google Scholar] [CrossRef]
- Kacharava, A.G.; Tertov, V.V.; Orekhov, A.N. Autoantibodies against low-density lipoprotein and atherogenic potential of blood. Ann. Med. 1993, 25, 551–555. [Google Scholar]
- Gounopoulos, P.; Merki, E.; Hansen, L.F.; Choi, S.H.; Tsimikas, S. Antibodies to oxidized low density lipoprotein: Epidemiological studies and potential clinical applications in cardiovascular disease. Minerva Cardioangiol. 2007, 55, 821–837. [Google Scholar]
- Burut, D.F.; Karim, Y.; Ferns, G.A. The role of immune complexes in atherogenesis. Angiology 2010, 61, 679–689. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Chistiakov, D.A.; Bobryshev, Y.V.; Orekhov, A.N. Blood atherogenicity as a target for anti-atherosclerotic therapy. Curr. Pharm. Des. 2013, 19, 5954–5962. [Google Scholar] [CrossRef]
- Iughetti, L.; Volta, C.; Maggi, E.; Palladini, G.; Perugini, C.; Bellomo, G.; Bernasconi, S. Circulating antibodies recognizing oxidatively modified low-density lipoprotein in children. Pediatr. Res. 1999, 45, 94–99. [Google Scholar] [CrossRef]
- Wang, J.; Niu, D.; Meng, Y.; Han, A.; Li, K.; Zhang, C. Plasma oxidized lipoprotein(a) and its immune complexes are present in newborns and children. Clin. Chim. Acta 2009, 407, 1–5. [Google Scholar] [CrossRef]
- Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989, 320, 915–924. [Google Scholar] [CrossRef]
- Yoshida, H.; Kisugi, R. Mechanisms of LDL oxidation. Clin. Chim. Acta 2010, 411, 1875–1882. [Google Scholar] [CrossRef]
- Delporte, C.; van Antwerpen, P.; Vanhamme, L.; Roumeguère, T.; Zouaoui Boudjeltia, K. Low-density lipoprotein modified by myeloperoxidase in inflammatory pathways and clinical studies. Mediat. Inflamm. 2013, 2013, 971579. [Google Scholar]
- Delporte, C.; Boudjeltia, K.Z.; Noyon, C.; Furtmüller, P.G.; Nuyens, V.; Slomianny, M.C.; Madhoun, P.; Desmet, J.M.; Raynal, P.; Dufour, D.; et al. Impact of myeloperoxidase-LDL interactions on enzyme activity and subsequent posttranslational oxidative modifications of apoB-100. J. Lipid Res. 2014, 55, 747–757. [Google Scholar] [CrossRef]
- Tertov, V.V.; Kaplun, V.V.; Sobenin, I.A.; Orekhov, A.N. Low-density lipoprotein modification occurring in human plasma. Possible mechanism of in vivo lipoprotein desialylation as a primary step of atherogenic modification. Atherosclerosis 1998, 138, 183–195. [Google Scholar] [CrossRef]
- Ermak, N.; Lacour, B.; Drüeke, T.B.; Vicca, S. Role of reactive oxygen species and Bax in oxidized low density lipoprotein-induced apoptosis of human monocytes. Atherosclerosis 2008, 200, 247–256. [Google Scholar] [CrossRef]
- Sun, L.; Ishida, T.; Yasuda, T.; Kojima, Y.; Honjo, T.; Yamamoto, Y.; Yamamoto, H.; Ishibashi, S.; Hirata, K.; Hayashi, Y. RAGE mediates oxidized LDL-induced pro-inflammatory effects and atherosclerosis in non-diabetic LDL receptor-deficient mice. Cardiovasc. Res. 2009, 82, 371–381. [Google Scholar]
- Pirillo, A.; Catapano, A.L. Soluble lectin-like oxidized low density lipoprotein receptor-1 as a biochemical marker for atherosclerosis-related diseases. Dis. Markers 2013, 35, 413–418. [Google Scholar] [CrossRef]
- Claise, C.; Edeas, M.; Chalas, J.; Cockx, A.; Abella, A.; Capel, L.; Lindenbaum, A. Oxidized low-density lipoprotein induces the production of interleukin-8 by endothelial cells. FEBS Lett. 1996, 398, 223–227. [Google Scholar] [CrossRef]
- Ryoo, S.W.; Kim, D.U.; Won, M.; Chung, K.S.; Jang, Y.J.; Oh, G.T.; Park, S.K.; Maeng, P.J.; Yoo, H.S.; Hoe, K.L. Native LDL induces interleukin-8 expression via H2O2, p38 Kinase, and activator protein-1 in human aortic smooth muscle cells. Cardiovasc. Res. 2004, 62, 185–193. [Google Scholar] [CrossRef]
- Wallenfeldt, K.; Fagerberg, B.; Wikstrand, J.; Hulthe, J. Oxidized low-density lipoprotein in plasma is a prognostic marker of subclinical atherosclerosis development in clinically healthy men. J. Intern. Med. 2004, 256, 413–420. [Google Scholar] [CrossRef]
- Chen, H.W.; Kuo, C.L.; Huang, C.S.; Kuo, S.J.; Liu, C.S. Oxidized low-density lipoproteins, autoantibodies against oxidized low-density lipoproteins and carotid intima media thickness in a clinically healthy population. Cardiology 2008, 110, 252–259. [Google Scholar] [CrossRef]
- Lobbes, M.B.; Lutgens, E.; Heeneman, S.; Cleutjens, K.B.; Kooi, M.E.; van Engelshoven, J.M.; Daemen, M.J.; Nelemans, P.J. Is there more than C-reactive protein and fibrinogen? The prognostic value of soluble CD40 ligand, interleukin-6 and oxidized low-density lipoprotein with respect to coronary and cerebral vascular disease. Atherosclerosis 2006, 187, 18–25. [Google Scholar] [CrossRef]
- Ehara, S.; Ueda, M.; Naruko, T.; Haze, K.; Itoh, A.; Otsuka, M.; Komatsu, R.; Matsuo, T.; Itabe, H.; Takano, T.; et al. Elevated levels of oxidized low density lipoprotein show a positive relationship with the severity of acute coronary syndromes. Circulation 2001, 103, 1955–1960. [Google Scholar] [CrossRef]
- Suzuki, T.; Kohno, H.; Hasegawa, A.; Toshima, S.; Amaki, T.; Kurabayashi, M.; Nagai, R. Diagnostic implications of circulating oxidized low density lipoprotein levels as a biochemical risk marker of coronary artery disease. Clin. Biochem. 2002, 35, 347–353. [Google Scholar] [CrossRef]
- Fraley, A.E.; Tsimikas, S. Clinical applications of circulating oxidized low-density lipoprotein biomarkers in cardiovascular disease. Curr. Opin. Lipidol. 2006, 17, 502–509. [Google Scholar] [CrossRef]
- Fraley, A.E.; Schwartz, G.G.; Olsson, A.G.; Kinlay, S.; Szarek, M.; Rifai, N.; Libby, P.; Ganz, P.; Witztum, J.L.; Tsimikas, S. Relationship of oxidized phospholipids and biomarkers of oxidized low-density lipoprotein with cardiovascular risk factors, inflammatory biomarkers, and effect of statin therapy in patients with acute coronary syndromes: Results from the MIRACL (Myocardial Ischemia Reduction With Aggressive Cholesterol Lowering) trial. J. Am. Coll. Cardiol. 2009, 53, 2186–2196. [Google Scholar] [CrossRef]
- Pryor, W.A.; Stanley, J.P. Letter: A suggested mechanism for the production of malondialdehyde during the autoxidation of polyunsaturated fatty acids. Nonenzymatic production of prostaglandin endoperoxides during autoxidation. J. Org. Chem. 1975, 40, 3615–3617. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Haberland, M.E.; Fogelman, A.M.; Edwards, P.A. Specificity of receptor-mediated recognition of malondialdehyde-modified low density lipoproteins. Proc. Natl. Acad. Sci. USA 1982, 79, 1712–1716. [Google Scholar] [CrossRef]
- Kreuzer, J.; White, A.L.; Knott, T.J.; Jien, M.L.; Mehrabian, M.; Scott, J.; Young, S.G.; Haberland, M.E. Amino terminus of apolipoprotein B suffices to produce recognition of malondialdehyde-modified low density lipoprotein by the scavenger receptor of human monocyte-macrophages. J. Lipid Res. 1997, 38, 324–342. [Google Scholar]
- Tanaga, K.; Bujo, H.; Inoue, M.; Mikami, K.; Kotani, K.; Takahashi, K.; Kanno, T.; Saito, Y. Increased circulating malondialdehyde-modified LDL levels in patients with coronary artery diseases and their association with peak sizes of LDL particles. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 662–666. [Google Scholar] [CrossRef]
- Miyazaki, T.; Shimada, K.; Sato, O.; Kotani, K.; Kume, A.; Sumiyoshi, K.; Sato, Y.; Ohmura, H.; Watanabe, Y.; Mokuno, H.; et al. Circulating malondialdehyde-modified LDL and atherogenic lipoprotein profiles measured by nuclear magnetic resonance spectroscopy in patients with coronary artery disease. Atherosclerosis 2005, 179, 139–145. [Google Scholar] [CrossRef]
- Carmena, R.; Duriez, P.; Fruchart, J.-C. Atherosclerosis: Evolving vascular biology and clinical implications—Atherogenic lipoprotein particles in atherosclerosis. Circulation 2004, 109, 2–7. [Google Scholar] [CrossRef]
- Takahashi, R.; Imamura, A.; Yoshikane, M.; Suzuki, M.; Cheng, X.W.; Numaguchi, Y.; Ikeda, N.; Murohara, T.; Okumura, K. Circulating malondialdehyde-modified low-density lipoprotein is strongly associated with very small low-density lipoprotein cholesterol concentrations in healthy men. Clin. Chim. Acta 2009, 399, 74–78. [Google Scholar] [CrossRef]
- Sánchez-Quesada, J.L.; Pèrez, A. Modified lipoproteins as biomarkers of cardiovascular risk in diabetes mellitus. Endocrinol. Nutr. 2013, 60, 518–528. [Google Scholar] [CrossRef]
- Cohen, M.P.; Lautenslager, G.; Shea, E. Glycated LDL concentrations in non-diabetic and diabetic subjects measured with monoclonal antibodies reactive with glycated apolipoprotein B epitopes. Eur. J. Clin. Chem. Clin. Biochem. 1993, 31, 707–713. [Google Scholar]
- Younis, N.N.; Soran, H.; Pemberton, P.; Charlton-Menys, V.; Elseweidy, M.M.; Durrington, P.N. Small dense LDL is more susceptible to glycation than more buoyant LDL in type 2 diabetes. Clin. Sci. 2013, 124, 343–349. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Tertov, V.V.; Orekhov, A.N. Characterization of chemical composition of native and modified low density lipoprotein occurring in the blood of diabetic patients. Int. Angiol. 1994, 13, 78–83. [Google Scholar]
- Sobenin, I.A.; Tertov, V.V.; Orekhov, A.N. Atherogenic modified LDL in diabetes. Diabetes 1996, 45, S35–S39. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Tertov, V.V.; Koschinsky, T.; Bünting, C.E.; Slavina, E.S.; Dedov, I.I.; Orekhov, A.N. Modified low density lipoprotein from diabetic patients causes cholesterol accumulation in human intimal aortic cells. Atherosclerosis 1993, 100, 41–54. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Tertov, V.V.; Orekhov, A.N.; Smirnov, V.N. Synergetic effect of desialylated and glycated low density lipoproteins on cholesterol accumulation in cultured smooth muscle intimal cells. Atherosclerosis 1991, 89, 151–154. [Google Scholar] [CrossRef]
- Younis, N.; Sharma, R.; Soran, H.; Charlton-Menys, V.; Elseweidy, M.; Durrington, P.N. Glycation as an atherogenic modification of LDL. Curr. Opin. Lipidol. 2008, 19, 378–384. [Google Scholar] [CrossRef]
- Del Turco, S.; Basta, G. An update on advanced glycation endproducts and atherosclerosis. Biofactors 2012, 38, 266–274. [Google Scholar] [CrossRef]
- De Michele, G.; Correale, M.; de Michele, O.; Guerra, V.; Mazzarelli, R.; Misciagna, G. Evaluation of serum biomarkers in nutritional disorders: Glycated apolipoprotein B, fasting serum glucose, fructosamine, stable and labile glycated hemoglobin in diabetic and non-diabetic subjects. Immunopharmacol. Immunotoxicol. 2008, 30, 925–936. [Google Scholar] [CrossRef]
- Misciagna, G.; Logroscino, G.; de Michele, G.; Guerra, V.; Cisternino, A.M.; Caruso, M.G.; Trevisan, M. Glycated apolipoprotein B and myocardial infarction. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 6–12. [Google Scholar] [CrossRef]
- Hayashi, Y.; Okumura, K.; Matsui, H.; Imamura, A.; Miura, M.; Takahashi, R.; Murakami, R.; Ogawa, Y.; Numaguchi, Y.; Murohara, T. Impact of low-density lipoprotein particle size on carotid intima-media thickness in patients with type 2 diabetes mellitus. Metabolism 2007, 56, 608–613. [Google Scholar] [CrossRef]
- Liu, M.L.; Ylitalo, K.; Nuotio, I.; Salonen, R.; Salonen, J.T.; Taskinen, M.R. Association between carotid intima-media thickness and low-density lipoprotein size and susceptibility of low-density lipoprotein to oxidation in asymptomatic members of familial combined hyperlipidemia families. Stroke 2002, 33, 1255–1260. [Google Scholar] [CrossRef]
- Inukai, T.; Yamamoto, R.; Suetsugu, M.; Matsumoto, S.; Wakabayashi, S.; Inukai, Y.; Matsutomo, R.; Takebayashi, K.; Aso, Y. Small low-density lipoprotein and small low-density lipoprotein/total low-density lipoprotein are closely associated with intima-media thickness of the carotid artery in Type 2 diabetic patients. J. Diabetes Complicat. 2005, 19, 269–275. [Google Scholar] [CrossRef]
- Basnakian, A.G.; Shah, S.V.; Ok, E.; Altunel, E.; Apostolov, E.O. Carbamylated LDL. Adv. Clin. Chem. 2010, 51, 25–52. [Google Scholar] [CrossRef]
- Wang, Z.; Nicholls, S.J.; Rodriguez, E.R.; Kummu, O.; Hörkkö, S.; Barnard, J.; Reynolds, W.F.; Topol, E.J.; DiDonato, J.A.; Hazen, S.L. Protein carbamylation links inflammation, smoking, uremia and atherogenesis. Nat. Med. 2007, 13, 1176–1184. [Google Scholar] [CrossRef]
- Shiu, S.W.; Xiao, S.M.; Wong, Y.; Chow, W.S.; Lam, K.S.; Tan, K.C. Carbamylation of LDL and its relationship with myeloperoxidase in type 2 diabetes mellitus. Clin. Sci. 2014, 126, 175–181. [Google Scholar] [CrossRef]
- Gonen, B.; Cole, T.; Hahm, K.S. The interaction of carbamylated low-density lipoprotein with cultured cells—Studies with human fibroblasts, rat peritoneal macrophages and human monocyte-derived macrophages. Biochim. Biophys. Acta 1983, 754, 201–207. [Google Scholar] [CrossRef]
- Roxborough, H.E.; Young, I.S. Carbamylation of proteins and atherogenesis in renal failure. Med. Hypotheses 1995, 45, 125–128. [Google Scholar] [CrossRef]
- Kumar, J.; Shah, S.V. Kidney disease as an independent risk factor for cardiovascular events. J. Ren. Nutr. 2005, 15, 99–104. [Google Scholar] [CrossRef]
- Hörkkö, S.; Huttunen, K.; Kervinen, K.; Kesāniemi, Y.A. Decreased clearance of uraemic and mildly carbamylated low-density lipoprotein. Eur. J. Clin. Investig. 1994, 24, 105–113. [Google Scholar] [CrossRef]
- Ok, E.; Basnakian, A.G.; Apostolov, E.O.; Barri, Y.M.; Shah, S.V. Carbamylated low-density lipoprotein induces death of endothelial cells: A link to atherosclerosis in patients with kidney disease. Kidney Int. 2005, 68, 173–178. [Google Scholar] [CrossRef]
- Apostolov, E.O.; Ok, E.; Burns, S.; Nawaz, S.; Savenka, A.; Shah, S.; Basnakian, A.G. Carbamylated-oxidized LDL: Proatherosclerotic effects on endothelial cells and macrophages. J. Atheroscler. Thromb. 2013, 20, 878–892. [Google Scholar] [CrossRef]
- Speer, T.; Owala, F.O.; Holy, E.W.; Zewinger, S.; Frenzel, F.L.; Stähli, B.E.; Razavi, M.; Triem, S.; Cvija, H.; Rohrer, L.; et al. Carbamylated low-density lipoprotein induces endothelial dysfunction. Eur. Heart J. 2014. [Google Scholar] [CrossRef]
- Asci, G.; Basci, A.; Shah, S.V.; Basnakian, A.; Toz, H.; Ozkahya, M.; Duman, S.; Ok, E. Carbamylated low-density lipoprotein induces proliferation and increases adhesion molecule expression of human coronary artery smooth muscle cells. Nephrology 2008, 13, 480–486. [Google Scholar] [CrossRef]
- Apostolov, E.O.; Shah, S.V.; Ok, E.; Basnakian, A.G. Carbamylated low-density lipoprotein induces monocyte adhesion to endothelial cells through intercellular adhesion molecule-1 and vascular cell adhesion molecule-1. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 826–832. [Google Scholar] [CrossRef]
- Carracedo, J.; Merino, A.; Briceno, C.; Soriano, S.; Buendía, P.; Calleros, L.; Rodriguez, M.; Martín-Malo, A.; Aljama, P.; Ramírez, R. Carbamylated low-density lipoprotein induces oxidative stress and accelerated senescence in human endothelial progenitor cells. FASEB J. 2011, 25, 1314–1322. [Google Scholar] [CrossRef]
- Kummu, O.; Turunen, S.P.; Wang, C.; Lehtimäki, J.; Veneskoski, M.; Kastarinen, H.; Koivula, M.K.; Risteli, J.; Kesäniemi, Y.A.; Hrkkö, S. Carbamyl adducts on low-density lipoprotein induce IgG response in LDLR−/− mice and bind plasma autoantibodies in humans under enhanced carbamylation. Antioxid. Redox Signal. 2013, 19, 1047–1062. [Google Scholar] [CrossRef]
- Kummu, O.; Turunen, S.P.; Prus, P.; Lehtimäki, J.; Veneskoski, M.; Wang, C.; Hörkkö, S. Human monoclonal Fab and human plasma antibodies to carbamyl-epitopes cross-react with malondialdehyde-adducts. Immunology 2014, 141, 416–430. [Google Scholar] [CrossRef]
- Tertov, V.V.; Orekhov, A.N.; Sobenin, I.A.; Morrisett, J.D.; Gotto, A.M., Jr.; Guevara, J.G., Jr. Carbohydrate composition of protein and lipid components in sialic acid-rich and -poor low density lipoproteins from subjects with and without coronary artery disease. J. Lipid Res. 1993, 34, 365–375. [Google Scholar]
- Attie, A.D.; Weinstein, D.B.; Freeze, H.H.; Pittman, R.C.; Steinberg, D. Unaltered catabolism of desialylated low-density lipoprotein in the pig and in cultured rat hepatocytes. Biochem. J. 1979, 180, 647–654. [Google Scholar]
- Bartlett, A.L.; Stanley, K.K. All low density lipoprotein particles are partially desialylated in plasma. Atherosclerosis 1998, 138, 237–245. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Tertov, V.V.; Mukhin, D.N. Desialylated low density lipoprotein–naturally occurring modified lipoprotein with atherogenic potency. Atherosclerosis 1991, 86, 153–161. [Google Scholar] [CrossRef]
- Tertov, V.V.; Sobenin, I.A.; Orekhov, A.N. Modified (desialylated) low-density lipoprotein measured in serum by lectin-sorbent assay. Clin. Chem. 1995, 41, 1018–1021. [Google Scholar]
- Tertov, V.V.; Sobenin, I.A.; Orekhov, A.N. Characterization of desialylated low-density lipoproteins which cause intracellular lipid accumulation. Int. J. Tissue React. 1992, 14, 155–162. [Google Scholar]
- Tertov, V.V.; Orekhov, A.N.; Kacharava, A.G.; Sobenin, I.A.; Perova, N.V.; Smirnov, V.N. Low density lipoprotein-containing circulating immune complexes and coronary atherosclerosis. Exp. Mol. Pathol. 1990, 52, 300–308. [Google Scholar] [CrossRef]
- Grewal, T.; Bartlett, A.; Burgess, J.W.; Packer, N.H.; Stanley, K.K. Desialylated LDL uptake in human and mouse macrophages can be mediated by a lectin receptor. Atherosclerosis 1996, 121, 151–163. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Tertov, V.V.; Mukhin, D.N.; Mikhailenko, I.A. Modification of low density lipoprotein by desialylation causes lipid accumulation in cultured cells: Discovery of desialylated lipoprotein with altered cellular metabolism in the blood of atherosclerotic patients. Biochem. Biophys. Res. Commun. 1989, 162, 206–211. [Google Scholar] [CrossRef]
- Ruelland, A.; Gallou, G.; Legras, B.; Paillard, F.; Cloarec, L. LDL sialic acid content in patients with coronary artery disease. Clin. Chim. Acta 1993, 221, 127–133. [Google Scholar] [CrossRef]
- Garner, B.; Harvey, D.J.; Royle, L.; Frischmann, M.; Nigon, F.; Chapman, M.J.; Rudd, P.M. Characterization of human apolipoprotein B100 oligosaccharides in LDL subfractions derived from normal and hyperlipidemic plasma: Deficiency of α-N-acetylneuraminyllactosyl-ceramide in light and small dense LDL particles. Glycobiology 2001, 11, 791–802. [Google Scholar] [CrossRef]
- Dousset, N.; Dousset, J.C.; Taus, M.; Ferretti, G.; Curatola, G.; Soléra, M.L.; Valdiguié, P. Effect of desialylation on low density lipoproteins: Comparative study before and after oxidative stress. Biochem. Mol. Biol. Int. 1994, 32, 555–563. [Google Scholar]
- Tertov, V.V.; Sobenin, I.A.; Kaplun, V.V.; Orekhov, A.N. Antioxidant content in low density lipoprotein and lipoprotein oxidation in vivo and in vitro. Free Radic. Res. 1998, 29, 165–173. [Google Scholar]
- Tertov, V.V.; Kaplun, V.V.; Sobenin, I.A.; Boytsova, E.Y.; Bovin, N.V.; Orekhov, A.N. Human plasma trans-sialidase causes atherogenic modification of low density lipoprotein. Atherosclerosis 2001, 159, 103–115. [Google Scholar] [CrossRef]
- Tanaka, K.; Tokumaru, S.; Kojo, S. Possible involvement of radical reactions in desialylation of LDL. FEBS Lett. 1997, 413, 202–204. [Google Scholar] [CrossRef]
- Harada, L.M.; Carvalho, M.D.; Passarelli, M.; Quintão, E.C. Lipoprotein desialylation simultaneously enhances the cell cholesterol uptake and impairs the reverse cholesterol transport system: In vitro evidences utilizing neuraminidase-treated lipoproteins and mouse peritoneal macrophages. Atherosclerosis 1998, 139, 65–75. [Google Scholar] [CrossRef]
- Bartlett, A.L.; Grewal, T.; de Angelis, E.; Myers, S.; Stanley, K.K. Role of the macrophage galactose lectin in the uptake of desialylated LDL. Atherosclerosis 2000, 153, 219–230. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Tertov, V.V.; Kabakov, A.E.; Adamova, I.Yu.; Pokrovsky, S.N.; Smirnov, V.N. Autoantibodies against modified low density lipoprotein—Nonlipid factor of blood plasma that stimulates foam cell formation. Arterioscler. Thromb. 1991, 11, 316–326. [Google Scholar] [CrossRef]
- Tertov, V.V.; Sobenin, I.A.; Gabbasov, Z.A.; Popov, E.G.; Jaakkola, O.; Solakivi, T.; Nikkari, T.; Smirnov, V.N.; Orekhov, A.N. Multiple-modified desialylated low density lipoproteins that cause intracellular lipid accumulation. Isolation, fractionation and characterization. Lab. Investig. 1992, 67, 665–675. [Google Scholar]
- Tertov, V.V.; Sobenin, I.A.; Orekhov, A.N.; Jaakkola, O.; Solakivi, T.; Nikkari, T. Characteristics of low density lipoprotein isolated from circulating immune complexes. Atherosclerosis 1996, 122, 191–199. [Google Scholar] [CrossRef]
- Avogaro, P.; Bittolo-Bon, G.; Cazzolato, G. Presence of a modified low density lipoprotein in humans. Arteriosclerosis 1988, 8, 79–87. [Google Scholar] [CrossRef]
- Krauss, R.M. Lipoprotein subfractions and cardiovascular disease risk. Curr. Opin. Lipidol. 2010, 21, 305–311. [Google Scholar] [CrossRef]
- La Belle, M.; Krauss, R.M. Differences in carbohydrate content of low density lipoproteins associated with low density lipoprotein subclass patterns. J. Lipid Res. 1990, 31, 1577–1588. [Google Scholar]
- Lopes-Virella, F.M.; Virella, G. Pathogenic role of modified LDL antibodies and immune complexes in atherosclerosis. J. Atherosler. Thromb. 2013, 20, 743–754. [Google Scholar] [CrossRef]
- Mironova, M.; Virella, G.; Lopes-Virella, M.F. Isolation and characterization of human antioxidized LDL autoantibodies. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 222–229. [Google Scholar] [CrossRef]
- Faria-Neto, J.R.; Chyu, K.Y.; Li, X.; Dimayuga, P.C.; Ferreira, C.; Yano, J.; Cercek, B.; Shah, P.K. Passive immunization with monoclonal IgM antibodies against phosphorylcholine reduces accelerated vein graft atherosclerosis in apolipoprotein E-null mice. Atherosclerosis 2006, 189, 83–90. [Google Scholar] [CrossRef]
- Virella, G.; Thorpe, S.R.; Alderson, N.L.; Derrick, M.B.; Chassereau, C.; Rhett, J.M.; Lopes-Virella, M.F. Definition of the immunogenic forms of modified human LDL recognized by human autoantibodies and by rabbit hyperimmune antibodies. J. Lipid Res. 2004, 45, 1859–1867. [Google Scholar] [CrossRef]
- Virella, G.; Lopes-Virella, M.F. Lipoprotein autoantibodies: Measurement and significance. Clin. Diagn. Lab. Immunol. 2003, 10, 499–505. [Google Scholar]
- Palinski, W.; Rosenfeld, M.E.; Ylä-Herttuala, S.; Gurtner, G.C.; Socher, S.S.; Butler, S.W.; Parthasarathy, S.; Carew, T.E.; Steinberg, D.; Witztum, J.L. Low density lipoprotein undergoes oxidative modification in vivo. Proc. Natl. Acad. Sci. USA 1989, 86, 1372–137. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Jukka, T.; Salonen, J.T.; Zhelankin, A.V.; Alexandra, A.; Melnichenko, A.A.; Kaikkonen, J.; Bobryshev, Y.V.; Orekhov, A.N. Low density lipoprotein-containing circulating immune complexes: Role in atherosclerosis and diagnostic value. BioMed Res. Int. 2014, 2014, 205697. [Google Scholar]
- Doo, Y.C.; Han, S.J.; Lee, J.H.; Cho, G.Y.; Hong, K.S.; Han, K.R.; Lee, N.H.; Oh, D.J.; Ryu, K.H.; Rhim, C.Y.; et al. Associations among oxidized low-density lipoprotein antibody, C-reactive protein, interleukin-6, and circulating cell adhesion molecules in patients with unstable angina pectoris. Am. J. Cardiol. 2004, 93, 554–558. [Google Scholar] [CrossRef]
- Papathanasiou, A.I.; Lourida, E.S.; Tsironis, L.D.; Goudevenos, J.A.; Tselepis, A.D. Short- and long-term elevation of autoantibody titers against oxidized LDL in patients with acute coronary syndromes—Role of the lipoprotein-associated phospholipase A2 and the effect of atorvastatin treatment. Atherosclerosis 2008, 196, 289–297. [Google Scholar] [CrossRef]
- Laczik, R.; Szodoray, P.; Veres, K.; Szomák, E.; Csipo, I.; Sipka, S., Jr.; Shoenfeld, Y.; Szekanecz, Z.; Soltész, P. Assessment of IgG antibodies to oxidized LDL in patients with acute coronary syndrome. Lupus 2011, 20, 730–735. [Google Scholar] [CrossRef]
- Ravandi, A.; Boekholdt, S.M.; Mallat, Z.; Talmud, P.J.; Kastelein, J.J.; Wareham, N.J.; Miller, E.R.; Benessiano, J.; Tedgui, A.; Witztum, J.L.; et al. Relationship of IgG and IgM autoantibodies and immune complexes to oxidized LDL with markers of oxidation and inflammation and cardiovascular events: Results from the EPIC-Norfolk Study. J. Lipid Res. 2011, 52, 1829–1836. [Google Scholar] [CrossRef]
- Karvonen, J.; Pāivänsalo, M.; Kesäniemi, Y.A.; Hörkkö, S. Immunoglobulin M type of autoantibodies to oxidized low-density lipoprotein has an inverse relation to carotid artery atherosclerosis. Circulation 2003, 108, 2107–2112. [Google Scholar] [CrossRef]
- De Geest, B.; Collen, D. Antibodies against oxidized LDL for non-invasive diagnosis of atherosclerotic vascular disease. Eur. Heart J. 2001, 22, 1517–1518. [Google Scholar] [CrossRef]
- Seccia, M.; Albano, E.; Maggi, E.; Bellomo, G. Circulating autoantibodies recognizing peroxidase-oxidized low density lipoprotein. Evidence for new antigenic epitopes formed in vivo independently from lipid peroxidation. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 134–140. [Google Scholar] [CrossRef]
- Shaw, P.X.; Hörkkö, S.; Chang, M.K.; Curtiss, L.K.; Palinski, W.; Silverman, G.J.; Witztum, J.L. Natural antibodies with the T15 idiotype may act in atherosclerosis, apoptotic clearance, and protective immunity. J. Clin. Investig. 2000, 105, 1731–1740. [Google Scholar] [CrossRef]
- Chou, M.Y.; Fogelstrand, L.; Hartvigsen, K.; Hansen, L.F.; Woelkers, D.; Shaw, P.X.; Choi, J.; Perkmann, T.; Bäckhed, F.; Miller, Y.I.; et al. Oxidation-specific epitopes are dominant targets of innate natural antibodies in mice and humans. J. Clin. Investig. 2009, 119, 1335–1349. [Google Scholar] [CrossRef]
- Orchard, T.J.; Virella, G.; Forrest, K.Y.; Evans, R.W.; Becker, D.J.; Lopes-Virella, M.F. Antibodies to oxidized LDL predict coronary artery disease in type 1 diabetes: A nested case-control study from the Pittsburgh Epidemiology of Diabetes Complications Study. Diabetes 1999, 48, 1454–1458. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Virella, G.; Orchard, T.J.; Koskinen, S.; Evans, R.W.; Becker, D.J.; Forrest, K.Y. Antibodies to oxidized LDL and LDL-containing immune complexes as risk factors for coronary artery disease in diabetes mellitus. Clin. Immunol. 1999, 90, 165–172. [Google Scholar] [CrossRef]
- Bertoia, M.L.; Pai, J.K.; Lee, J.H.; Taleb, A.; Joosten, M.M.; Mittleman, M.A.; Yang, X.; Witztum, J.L.; Rimm, E.B.; Tsimikas, S.; et al. Oxidation-specific biomarkers and risk of peripheral artery disease. J. Am. Coll. Cardiol. 2013, 61, 2169–2179. [Google Scholar] [CrossRef]
- Klimov, A.N.; Denisenko, A.D.; Popov, A.V.; Nagornev, V.A.; Pleskov, V.M.; Vinogradov, A.G.; Denisenko, T.V.; Magracheva, E.Ya.; Kheifes, G.M.; Kuznetzov, A.S. Lipoprotein-antibody immune complexes. Their catabolism and role in foam cell formation. Atherosclerosis 1985, 58, 1–15. [Google Scholar] [CrossRef]
- Klimov, A.N.; Denisenko, A.D.; Vinogradov, A.G.; Nagornev, V.A.; Pivovarova, Y.I.; Sitnikova, O.D.; Pleskov, V.M. Accumulation of cholesteryl esters in macrophages incubated with human lipoprotein-antibody autoimmune complex. Atherosclerosis 1988, 74, 41–46. [Google Scholar] [CrossRef]
- Griffith, R.L.; Virella, G.T.; Stevenson, H.C.; Lopes-Virella, M.F. Low density lipoprotein metabolism by human macrophages activated with low density lipoprotein immune complexes—A possible mechanism of foam cell formation. J. Exp. Med. 1988, 168, 1041–1059. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Griffith, R.L.; Shunk, K.A.; Virella, G.T. Enhanced uptake and impaired intracellular metabolism of low density lipoprotein complexed with anti-low density lipoprotein antibodies. Arterioscler. Thromb. 1991, 11, 1356–1367. [Google Scholar] [CrossRef]
- Morganelli, P.M.; Kitzmiller, T.J.; Hemmer, R.; Fanger, M.W. Redirected targeting of LDL to human monocyte Fcγ receptors with bispecific antibodies. Arterioscler. Thromb. 1992, 12, 1131–1138. [Google Scholar]
- Lopes-Virella, M.F.; Binzafar, N.; Rackley, S.; Takei, A.; La Via, M.; Virella, G. The uptake of LDL-IC by human macrophages: Predominant involvement of the FcγRI receptor. Atherosclerosis 1997, 135, 161–170. [Google Scholar] [CrossRef]
- Huang, Y.; Jaffa, A.; Koskinen, S.; Takei, A.; Lopes-Virella, M.F. Oxidized LDL-containing immune complexes induce Fcγ receptor I-mediated mitogen-activated protein kinase activation in THP-1 macrophages. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1600–1607. [Google Scholar] [CrossRef]
- Kiener, P.A.; Rankin, B.M.; Davis, P.M.; Yocum, S.A.; Warr, G.A.; Grove, R.I. Immune complexesof LDL induce atherogenic responses in human monocytic cells. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 990–999. [Google Scholar] [CrossRef]
- Nagarajan, S. Anti-oxLDL IgG blocks oxLDL interaction with CD36, but promotes FcγR, CD32A-dependent inflammatory cell adhesion. Immunol. Lett. 2007, 108, 52–61. [Google Scholar] [CrossRef]
- Lappalainen, J.; Lindstedt, K.A.; Oksjoki, R.; Kovanen, P.T. OxLDL-IgG immune complexes induce expression and secretion of proatherogenic cytokines by cultured human mast cells. Atherosclerosis 2011, 214, 357–363. [Google Scholar] [CrossRef]
- Virella, G.; Atchley, D.; Koskinen, S.; Zheng, D.; Lopes-Virella, M.F. Proatherogenic and proinflammatory properties of immune complexes prepared with purified human oxLDL antibodies and human oxLDL. Clin. Immunol. 2002, 105, 81–92. [Google Scholar] [CrossRef]
- Saad, A.F.; Virella, G.; Chassereau, C.; Boackle, R.J.; Lopes-Virella, M.F. OxLDL immune complexes activate complement and induce cytokine production by MonoMac 6 cells and human macrophages. J. Lipid Res. 2006, 47, 1975–1983. [Google Scholar] [CrossRef]
- Truman, J.P.; Al Gadban, M.M.; Smith, K.J.; Jenkins, R.W.; Mayroo, N.; Virella, G.; Lopes-Virella, M.F.; Bielawska, A.; Hannun, Y.A.; Hammad, S.M. Differential regulation of acid sphingomyelinase in macrophages stimulated with oxidized low-density lipoprotein (LDL) and oxidized LDL immune complexes: Role in phagocytosis and cytokine release. Immunology 2012, 136, 30–45. [Google Scholar] [CrossRef]
- Hammad, S.M.; Twal, W.O.; Barth, J.L.; Smith, K.J.; Saad, A.F.; Virella, G.; Argraves, W.S.; Lopes-Virella, M.F. Oxidized LDL immune complexes and oxidized LDL differentially affect the expression of genes involved with inflammation and survival in human U937 monocytic cells. Atherosclerosis 2009, 202, 394–404. [Google Scholar] [CrossRef]
- Virella, G.; Lopes-Virella, M.F. Atherogenesis and the humoral immune response to modified lipoproteins. Atherosclerosis 2008, 200, 239–246. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Kalenich, O.S.; Tertov, V.V.; Novikov, I.D.; Vorob’eva, E.G. Cholesterol levels in circulating immune complexes as a marker of coronary atherosclerosis. In Hypercholesterolemia, Hypocholesterolemia, Hypertriglyceridemia; Marmendier, C.L., Ed.; Plenum Press: New York, NY, USA, 1990; pp. 393–397. [Google Scholar]
- Tertov, V.V.; Sobenin, I.A.; Tonevitsky, A.G.; Orekhov, A.N.; Smirnov, V.N. Isolation of atherogenic modified (desialylated) low density lipoprotein from blood of atherosclerotic patients: Separation from native lipoprotein by affinity chromatography. Biochem. Biophys. Res. Commun. 1990, 167, 1122–1127. [Google Scholar] [CrossRef]
- Lecomte, E.; Herbeth, B.; Clerc, G.; Khalife, K.; Siest, G.; Artur, Y. Cholesterol content of circulating immune complexes in patients with coronary stenosis and subjects without evidence of atherosclerosis. Clin. Chem. 1995, 41, 1526–1531. [Google Scholar]
- Mironova, M.; Virella, G.; Virella-Lowell, I.; Lopes-Virella, M.F. Anti-modified LDL antibodies and LDL-containing immune complexes in IDDM patients and healthy controls. Clin. Immunol. Immunopathol. 1997, 85, 73–82. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; McHenry, M.B.; Lipsitz, S.; Yim, E.; Wilson, P.F.; Lackland, D.T.; Lyons, T.; Jenkins, A.J.; Virella, G. Immune complexes containing modified lipoproteins are related to the progression of internal carotid intima-media thickness in patients with type 1 diabetes. Atherosclerosis 2007, 190, 359–369. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Orekhova, V.A.; Melnichenko, A.A.; Bobryshev, Y.V.; Orekhov, A.N. Low density lipoprotein-containing circulating immune complexes have better prognostic value in carotid intima-media thickness progression than other lipid parameters. Int. J. Cardiol. 2013, 166, 747–748. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Karagodin, V.P.; Melnichenko, A.A.; Bobryshev, Y.V.; Orekhov, A.N. Diagnostic and prognostic value of low density lipoprotein-containing circulating immune complexes in atherosclerosis. J. Clin. Immunol. 2013, 33, 489–495. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Hunt, K.J.; Baker, N.L.; Lachin, J.; Nathan, D.M.; Virella, G. Levels of oxidized LDL and advanced glycation end products-modified LDL in circulating immune complexes are strongly associated with increased levels of carotid intima-media thickness and its progression in type 1 diabetes. Diabetes 2011, 60, 582–589. [Google Scholar] [CrossRef]
- Hunt, K.J.; Baker, N.; Cleary, P.; Backlund, J.Y.; Lyons, T.; Jenkins, A.; Virella, G.; Lopes-Virella, M.F. Oxidized LDL and AGE-LDL in circulating immune complexes strongly predict progression of carotid artery IMT in type 1 diabetes. Atherosclerosis 2013, 231, 315–322. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Baker, N.L.; Hunt, K.J.; Lachin, J.; Nathan, D.; Virella, G. Oxidized LDL immune complexes and coronary artery calcification in type 1 diabetes. Atherosclerosis 2011, 214, 462–467. [Google Scholar] [CrossRef]
- Lopes-Virella, M.F.; Hunt, K.J.; Baker, N.L.; Virella, G.; Moritz, T. The levels of MDA-LDL in circulating immune complexes predict myocardial infarction in the VADT study. Atherosclerosis 2012, 224, 526–531. [Google Scholar] [CrossRef]
- Holvoet, P.; Collen, D.; van de Werf, F. Malondialdehyde-modified LDL as a marker of acute coronary syndromes. JAMA 1999, 281, 1718–1721. [Google Scholar] [CrossRef]
- Yang, T.C.; Chen, Y.J.; Chang, S.F.; Chen, C.H.; Chang, P.Y.; Lu, S.C. Malondialdehyde mediates oxidized LDL-induced coronary toxicity through the Akt-FGF2 pathway via DNA methylation. J. Biomed. Sci. 2014, 21, 11. [Google Scholar] [CrossRef]
- Lyons, T.J.; Jenkins, A.J.; Zheng, D.; Klein, R.L.; Otvos, J.D.; Yu, Y.; Lackland, D.T.; McGee, D.; McHenry, M.B.; Lopes-Virella, M.F.; et al. Nuclear magnetic resonance-determined lipoprotein subclass profile in the DCCT/EDIC cohort: Associations with carotid intima-media thickness. Diabet. Med. 2006, 23, 955–966. [Google Scholar] [CrossRef]
- Klein, R.L.; Carter, R.E.; Jenkins, A.J.; Lyons, T.J.; Baker, N.L.; Gilbert, G.E.; Virella, G.; Lopes-Virella, M.F. LDL-containing immune complexes in the DCCT/EDIC cohort: Associations with lipoprotein subclasses. J. Diabetes Complicat. 2011, 25, 73–82. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Orekhov, A.N.; Bobryshev, Y.V.; Sobenin, I.A.; Melnichenko, A.A.; Chistiakov, D.A. Modified Low Density Lipoprotein and Lipoprotein-Containing Circulating Immune Complexes as Diagnostic and Prognostic Biomarkers of Atherosclerosis and Type 1 Diabetes Macrovascular Disease. Int. J. Mol. Sci. 2014, 15, 12807-12841. https://doi.org/10.3390/ijms150712807
Orekhov AN, Bobryshev YV, Sobenin IA, Melnichenko AA, Chistiakov DA. Modified Low Density Lipoprotein and Lipoprotein-Containing Circulating Immune Complexes as Diagnostic and Prognostic Biomarkers of Atherosclerosis and Type 1 Diabetes Macrovascular Disease. International Journal of Molecular Sciences. 2014; 15(7):12807-12841. https://doi.org/10.3390/ijms150712807
Chicago/Turabian StyleOrekhov, Alexander N., Yuri V. Bobryshev, Igor A. Sobenin, Alexandra A. Melnichenko, and Dimitry A. Chistiakov. 2014. "Modified Low Density Lipoprotein and Lipoprotein-Containing Circulating Immune Complexes as Diagnostic and Prognostic Biomarkers of Atherosclerosis and Type 1 Diabetes Macrovascular Disease" International Journal of Molecular Sciences 15, no. 7: 12807-12841. https://doi.org/10.3390/ijms150712807
APA StyleOrekhov, A. N., Bobryshev, Y. V., Sobenin, I. A., Melnichenko, A. A., & Chistiakov, D. A. (2014). Modified Low Density Lipoprotein and Lipoprotein-Containing Circulating Immune Complexes as Diagnostic and Prognostic Biomarkers of Atherosclerosis and Type 1 Diabetes Macrovascular Disease. International Journal of Molecular Sciences, 15(7), 12807-12841. https://doi.org/10.3390/ijms150712807