Identification of Cold-Responsive miRNAs and Their Target Genes in Nitrogen-Fixing Nodules of Soybean

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
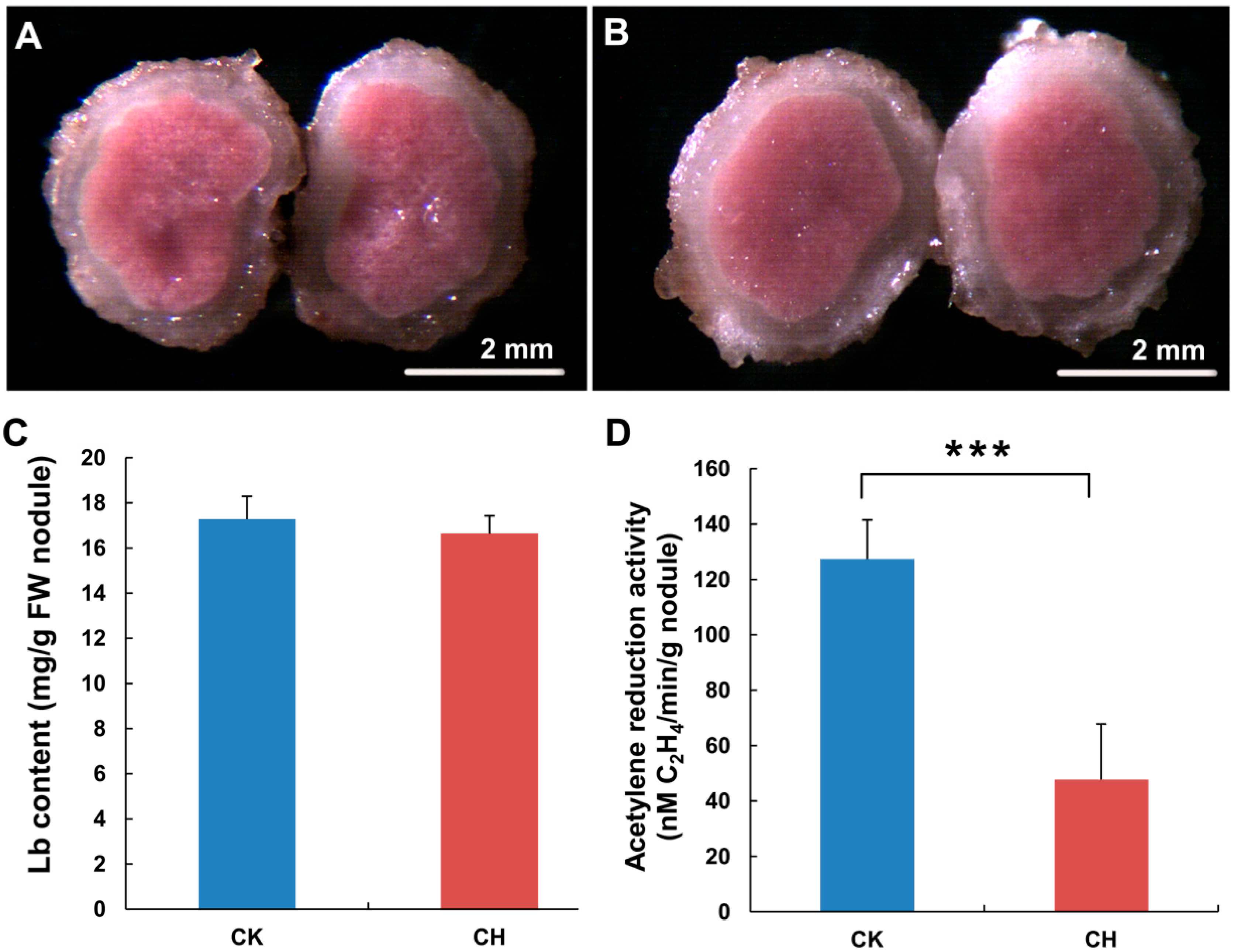
2.1.1. The Nitrogen Use Efficiency of Nitrogen-Fixing Nodules Was Decreased in Response to Low Temperature Treatment
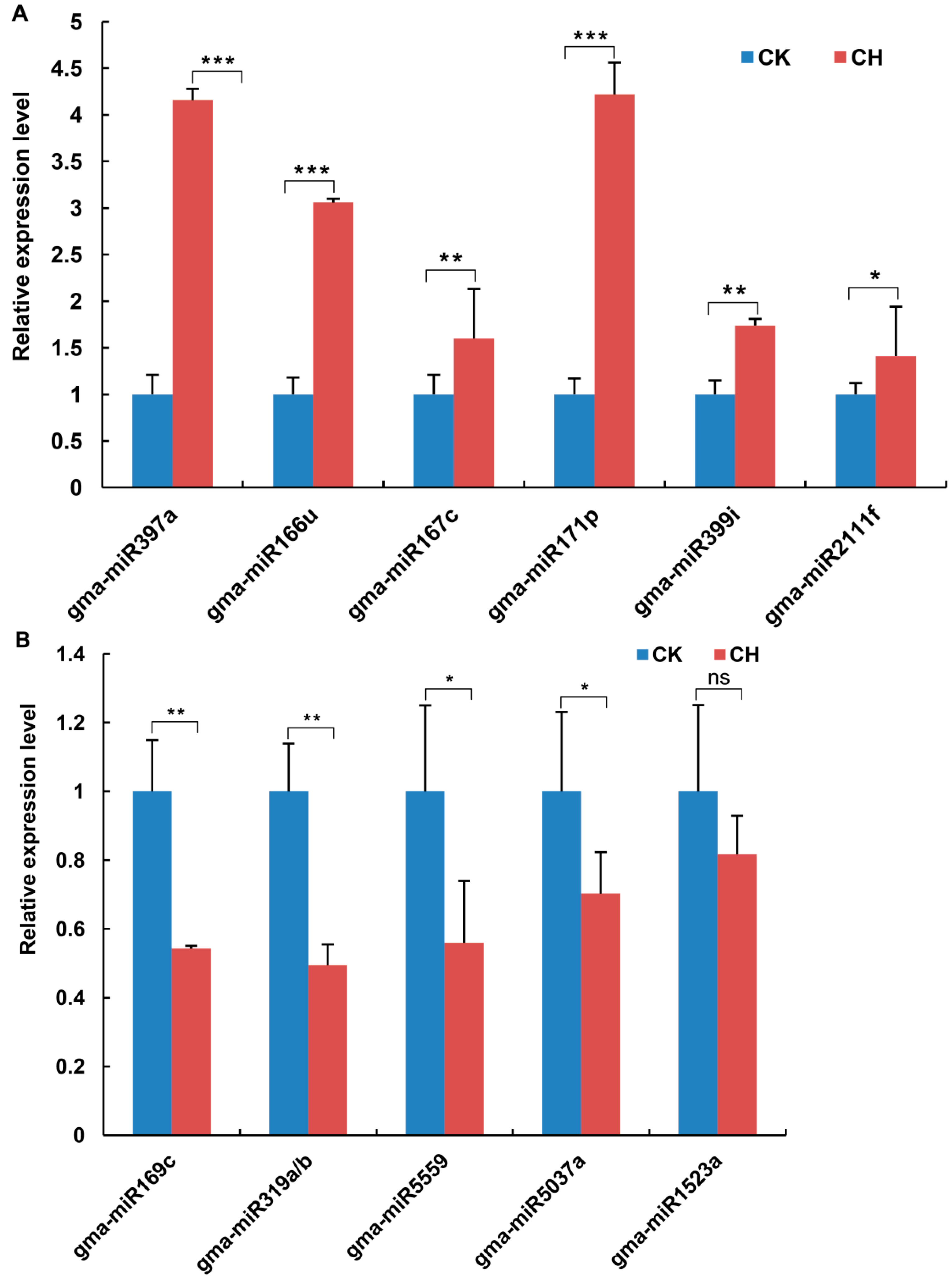
2.1.2. Identification and Validation of Low Temperature-Responsive miRNAs in Functional Nodules


2.1.3. Target Prediction and Validation for the Identified Low Temperature-Responsive miRNAs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Target Gene | Target Annotation |
|---|---|---|
| gma-miR397a | Glyma01g26750 a,b | Multicopper oxidase, oxidation/reduction |
| Glyma18g07240 b | Multicopper oxidase, oxidation/reduction | |
| Glyma03g15800 a,b | Multicopper oxidase, oxidation/reduction | |
| Glyma03g14450 b | Multicopper oxidase, oxidation/reduction | |
| Glyma06g43700 b | Multicopper oxidase, oxidation/reduction | |
| Glyma12g14230 b | Multicopper oxidase, oxidation/reduction | |
| gma-miR166u | Glyma08g21620 a,c | bZIP transcription factor |
| Glyma07g01940 a,c | Homeobox-leucine zipper protein | |
| gma-miR171p | Glyma01g18040 a,b,c | GRAS family transcription factor |
| Glyma11g17490 a,c | GRAS family transcription factor | |
| gma-miR2111f | Glyma19g01430 a,b | EamA-like transporter family |
| Glyma15g34831 a,b | EamA-like transporter family | |
| Glyma16g06160 b,c | Kelch motif; Protin Binding; Kelch-related protein | |
| Glyma19g25770 b,c | Kelch motif; Protin Binding; Kelch-related protein | |
| gma-miR169c | Glyma02g35190 a,b,c | HAP2 like transcription factor |
| Glyma19g38800 a,b | HAP2 like transcription factor | |
| Glyma13g16770 a,b | HAP2 like transcription factor | |
| Glyma07g04050 a,b | HAP2 like transcription factor | |
| Glyma13g27230 a,b | HAP2 like transcription factor | |
| gma-miR5037a | Glyma17g13680 a,b | GRAS family transcription factor |
| Glyma05g03020 a,b | GRAS family transcription factor | |
| gma-miR167c | Glyma11g31940 a,b | Auxin response factor |
| Glyma18g05330 a,b | Auxin response factor | |
| Glyma02g40650 a,b | Auxin response factor | |
| Glyma14g03650 a,b | Auxin response factor | |
| Glyma14g38940 a,b | Auxin response factor | |
| Glyma08g10550 a,b | Auxin response factor | |
| gma-miR319a/b | Glyma13g04030 a,b | MYB family transcription factor |
| Glyma13g25720 a,b | MYB family transcription factor | |
| Glyma15g35860 a,b | MYB family transcription factor | |
| Glyma20g11040 a,b | MYB family transcription factor | |
| Glyma06g43720 a,b | TCP family transcription factor | |
| Glyma12g14200 a,b | TCP family transcription factor | |
| gma-miR399i | Glyma10g04230 a,b | Inorganic phosphate and sugar transporter |
| Glyma14g36650 b | Inorganic phosphate and sugar transporter | |
| Glyma19g34710 a | Inorganic phosphate and sugar transporter | |
| Glyma20g34620 a | Inorganic phosphate and sugar transporter | |
| gma-miR1523a | Glyma13g25040 b | Putative lysophospholipase |
| Glyma13g25050 b | Putative lysophospholipase | |
| gma-miR5559 | Glyma20g34670 b | Mediator complex subunit 28 |
| Glyma02g19340 b | No functional annotation | |
| Glyma14g02030 b | Protein binding | |
| Glyma02g46640 b | Protein binding |

2.1.4. Correlation between the Expression of miRNAs and Their Targets in Response to Low Temperature Stress in Mature Nodules

2.2. Discussion
3. Experimental Section
3.1. Plant Materials and Low Temperature Treatment
3.2. Analysis of the Leghemoglobin (Lb) Concentration
3.3. Analysis of the Acetylene Reduction Activity (ARA) Level

3.4. Small RNA Library Construction, Solexa Sequencing and Data Analysis
3.5. Quantitative Reverse Transcription PCR (RT-qPCR)
3.6. miRNA Target Gene Prediction and Experimental Validation
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Achard, P.; Gong, F.; Cheminant, S.; Alioua, M.; Hedden, P.; Genschik, P. The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. Plant Cell 2008, 20, 2117–2129. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.-K.; Sunkar, R. Gene regulation during cold stress acclimation in plants. Methods Mol. Biol. 2010, 639, 39–55. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta 2012, 1819, 137–148. [Google Scholar] [CrossRef]
- Sunkar, R.; Li, Y.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Cell 2012, 17, 196–203. [Google Scholar]
- Miura, K.; Furumoto, T. Cold signaling and cold response in plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, G.; Sutoh, K.; Zhu, J.; Zhang, W. Identification of cold-inducible microRNAs in plants by transcriptome analysis. Biochim. Biophys. Acta 2008, 1779, 780–788. [Google Scholar]
- Sunkar, R. Novel and stress-regulated microRNAs and other small rnas from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Huan, Q.; Chong, K. Deep sequencing of Brachypodium small RNAs at the global genome level identifies microRNAs involved in cold stress response. BMC Genomics 2009, 10, 449. [Google Scholar] [CrossRef]
- Tang, Z.; Zhang, L.; Xu, C.; Yuan, S.; Zhang, F.; Zheng, Y.; Zhao, C. Uncovering small RNA-mediated responses to cold stress in a wheat thermosensitive genic male-sterile line by deep sequencing. Plant Physiol. 2012, 159, 721–738. [Google Scholar] [CrossRef]
- Thiebaut, F.; Rojas, C.A.; Almeida, K.L.; Grativol, C.; Domiciano, G.C.; Lamb, C.R.C.; de Almeida Engler, J.; Hemerly, A.S.; Ferreira, P.C. Regulation of miR319 during cold stress in sugarcane. Plant Cell Environ. 2012, 35, 502–512. [Google Scholar] [CrossRef]
- Lu, S.; Sun, Y.-H.; Chiang, V.L. Stress-responsive microRNAs in Populus. Plant J. 2008, 55, 131–151. [Google Scholar] [CrossRef]
- Keyser, H.; Li, F. Potential for increasing biological nitrogen fixation in soybean. In Biological Nitrogen Fixation for Sustainable Agriculture; Ladha, J.K., George, T., Bohlool, B.B., Eds.; Springer: Berlin, Germany, 1992; pp. 119–135. [Google Scholar]
- Duke, S.; Scharader, L.; Miller, M. Low temperature effects on soybean (Glycine max) mitochondrial respiration and several dehydrogenases during imbibition and germination. Plant Physiol. 1977, 60, 716–722. [Google Scholar] [CrossRef]
- Walsh, K.; Layzell, D. Carbon and nitrogen assimilation and partitioning in soybeans exposed to low root temperatures. Plant Physiol. 1986, 80, 249–255. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Baird, L.M.; Escuredo, P.R.; Dalton, D.A.; Minchin, F.R.; Iturbe-Ormaetxe, I.; Rubio, M.C.; Moran, J.F.; Gordon, A.J.; Becana, M. Stress-induced legume root nodule senescence. Physiological, biochemical, and structural alterations. Plant Physiol. 1999, 121, 97–112. [Google Scholar] [CrossRef]
- Van Heerden, P.D.R.; Kiddle, G.; Pellny, T.K.; Mokwala, P.W.; Jordaan, A.; Strauss, A.J.; de Beer, M.; Schluter, U.; Kunert, K.J.; Foyer, C.H. Regulation of respiration and the oxygen diffusion barrier in soybean protect symbiotic nitrogen fixation from chilling-induced inhibition and shoots from premature senescence. Plant Physiol. 2008, 148, 316–327. [Google Scholar] [CrossRef]
- Allen, D.J.; Ort, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 2001, 6, 36–42. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Harbinson, J.; Williams, J.; Foyer, C.H. Effect of chilling on carbon assimilation, enzyme activation, and photosynthetic electron transport in the absence of photoinhibition in maize leaves. Plant Physiol. 1997, 114, 1039–1046. [Google Scholar]
- Joshi, T.; Yan, Z.; Libault, M.; Jeong, D.-H.; Park, S.; Green, P.J.; Sherrier, D.J.; Farmer, A.; May, G.; Meyers, B.C.; et al. Prediction of novel miRNAs and associated target genes in Glycine max. BMC Bioinform. 2010, 11, S14. [Google Scholar]
- Wang, Y.; Li, P.; Cao, X.; Wang, X.; Zhang, A.; Li, X. Identification and expression analysis of miRNAs from nitrogen-fixing soybean nodules. Biochem. Biophys. Res. Commun. 2009, 378, 799–803. [Google Scholar] [CrossRef]
- Song, Q.-X.; Liu, Y.-F.; Hu, X.-Y.; Zhang, W.-K.; Ma, B.; Chen, S.-Y.; Zhang, J.-S. Identification of miRNAs and their target genes in developing soybean seeds by deep sequencing. BMC Plant Biol. 2011, 11, 5. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Stellwag, E.J. Identification of soybean microRNAs and their targets. Planta 2008, 229, 161–182. [Google Scholar] [CrossRef]
- Wong, C.E.; Zhao, Y.T.; Wang, X.J.; Croft, L.; Wang, Z.H.; Haerizadeh, F.; Mattick, J.S.; Singh, M.B.; Carroll, B.J.; Bhalla, P.L. MicroRNAs in the shoot apical meristem of soybean. J. Exp. Bot. 2011, 62, 2495–2506. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; de Oliveira, L.F.V.; Molina, L.G.; Almerão, M.P.; Rodrigues, F.A.; Marcolino, J.; Barbosa, J.F.; Stolf-Moreira, R.; Nepomuceno, A.L.; Marcelino-Guimarães, F.C.; et al. Identification of novel soybean microRNAs involved in abiotic and biotic stresses. BMC Genomics 2011, 12, 307. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, N.E.; Torres, C.M.; Wagner, F.W. Proteolytic activity in soybean root nodules activity in host cell cytosol and bacteroids throughout physiological development and senescence. Plant Physiol. 1983, 71, 797–802. [Google Scholar] [CrossRef]
- Chen, P.-C.; Phillips, D.A. Induction of root nodule senescence by combined nitrogen in Pisum sativum L. Plant Physiol. 1977, 59, 440–442. [Google Scholar] [CrossRef]
- Delgado, M.J.; Garrido, J.M.; Ligero, F.; Lluch, C. Nitrogen fixation and carbon metabolism by nodules and bacteroids of pea plants under sodium chloride stress. Physiol. Plant. 1993, 89, 824–829. [Google Scholar] [CrossRef]
- Delgado, M.J.; Ligero, F.; Lluch, C. Effects of salt stress on growth and nitrogen fixation by pea, faba-bean, common bean and soybean plants. Soil Biol. Biochem. 1994, 26, 371–376. [Google Scholar] [CrossRef]
- Balestrasse, K.B.; Gardey, L.; Gallego, S.M.; Tomaro, M. Response of antioxidant defence system in soybean nodules and roots subjected to cadmium stress. Funct. Plant Biol. 2001, 28, 497–504. [Google Scholar] [CrossRef]
- López, M.; Herrera-Cervera, J.A.; Iribarne, C.; Tejera, N.A.; Lluch, C. Growth and nitrogen fixation in Lotus japonicus and Medicago. truncatula under NaCl stress: Nodule carbon metabolism. J. Plant Physiol. 2008, 165, 641–650. [Google Scholar] [CrossRef]
- Sánchez-Pardo, B.; Carpena, R.O.; Zornoza, P. Cadmium in white lupin nodules: Impact on nitrogen and carbon metabolism. J. Plant Physiol. 2013, 170, 265–271. [Google Scholar] [CrossRef]
- Navascués, J.; Pérez-Rontomé, C.; Gay, M.; Marcos, M.; Yang, F.; Walker, F.A.; Desbois, A.; Abián, J.; Becana, M. Leghemoglobin green derivatives with nitrated hemes evidence production of highly reactive nitrogen species during aging of legume nodules. Proc. Natl. Acad. Sci. USA 2012, 109, 2660–2665. [Google Scholar] [CrossRef] [Green Version]
- Vorster, B.J.; Schlüter, U.; du Plessis, M.; van Wyk, S.; Makgopa, M.E.; Ncube, I.; Quain, M.D.; Kunert, K.; Foyer, C.H. The cysteine protease–cysteine protease inhibitor system explored in soybean nodule development. Agronomy 2013, 3, 550–570. [Google Scholar] [CrossRef]
- Boyd, E.S.; Lange, R.K.; Mitchell, A.C.; Havig, J.R.; Hamilton, T.L.; Lafrenière, M.J.; Shock, E.L.; Peters, J.W.; Skidmore, M. Diversity, abundance, and potential activity of nitrifying and nitrate-reducing microbial assemblages in a subglacial ecosystem. Appl. Environ. Microb. 2011, 77, 4778–4787. [Google Scholar] [CrossRef]
- Fenta, B.A.; Driscoll, S.P.; Kunert, K.J.; Foyer, C.H. Characterization of drought-tolerance traits in nodulated soya beans: The importance of maintaining photosynthesis and shoot biomass under drought-induced limitations on nitrogen metabolism. J. Agron. Crop. Sci. 2012, 198, 92–103. [Google Scholar] [CrossRef]
- Furlan, A.; Bianucci, E.; del Carmen, T.M.; Castro, S.; Dietz, K. Antioxidant enzyme activities and gene expression patterns in peanut nodules during a drought and rehydration cycle. Funct. Plant Biol. 2014, 41, 81–86. [Google Scholar]
- Hungria, M.; Kaschuk, G. Regulation of N2 fixation and NO3−/NH4+ assimilation in nodulated and N-fertilized Phaseolus. vulgaris L. exposed to high temperature stress. Environ. Exp. Bot. 2014, 98, 32–39. [Google Scholar] [CrossRef]
- De Costa, W.; Becher, M.; Schubert, S. Effects of water stress on nitrogen fixation of common bean (Phaseolus. vulgaris L.). J. Nat. Sci. Found. Sri Lanka 2012, 25, 83–94. [Google Scholar]
- Palma, F.; Tejera, N.A.; Lluch, C. Nodule carbohydrate metabolism and polyols involvement in the response of Medicago sativa to salt stress. Environ. Exp. Bot. 2013, 85, 43–49. [Google Scholar] [CrossRef]
- Plantpan. Available online: http://plantpan.mbc.nctu.edu.tw/ (accessed on 17 July 2012).
- Dai, X.; Zhao, P. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, 155–159. [Google Scholar] [CrossRef]
- Ossowski, S.; Schwab, R.; Weigel, D. Gene silencing in plants using artificial microRNAs and other small RNAs. Plant J. 2008, 53, 674–690. [Google Scholar] [CrossRef]
- Simon, S.A.; Meyers, B.C.; Sherrier, D.J. MicroRNAs in the rhizobia legume symbiosis. Plant Physiol. 2009, 151, 1002–1008. [Google Scholar] [CrossRef]
- Devers, E.; Branscheid, A.; May, P.; Krajinski, F. Stars and symbiosis: MicroRNA- and microRNA*-mediated transcript cleavage involved in arbuscular mycorrhizal symbiosis. Plant Physiol. 2011, 156, 1990–2010. [Google Scholar] [CrossRef]
- Dong, Z.; Shi, L.; Wang, Y.; Chen, L.; Cai, Z.; Wang, Y.; Jin, J.; Li, X. Identification and dynamic regulation of microRNAs involved in salt stress responses in functional soybean nodules by high-throughput sequencing. Int. J. Mol. Sci. 2013, 14, 2717–2738. [Google Scholar]
- Lv, D.-K.; Bai, X.; Li, Y.; Ding, X.-D.; Ge, Y.; Cai, H.; Ji, W.; Wu, N.; Zhu, Y.-M. Profiling of cold-stress-responsive miRNAs in rice by microarrays. Gene 2010, 459, 39–47. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Y.; Ren, Y.; Xu, J.; Zhang, Z.; Wang, Y. Genome-wide identification of cold-responsive and new microRNAs in Populus tomentosa by high-throughput sequencing. Biochem. Biophys. Res. Commun. 2012, 417, 892–896. [Google Scholar] [CrossRef]
- Li, Y.; Fu, Y.; Ji, L.; Wu, C.; Zheng, C. Characterization and expression analysis of the Arabidopsis mir169 family. Plant Sci. 2010, 178, 271–280. [Google Scholar] [CrossRef]
- Zhang, X.-N.; Li, X.; Liu, J.-H. Identification of conserved and novel cold-responsive microRNAs in trifoliate orange (Poncirus. trifoliata (L.) Raf.) using high-throughput sequencing. Plant Mol. Biol. Rep. 2014, 32, 328–341. [Google Scholar] [CrossRef]
- Boualem, A.; Laporte, P.; Jovanovic, M.; Laffont, C.; Plet, J.; Combier, J.P.; Niebel, A.; Crespi, M.; Frugier, F. microRNA166 controls root and nodule development in Medicago truncatula. Plant J. 2008, 54, 876–887. [Google Scholar] [CrossRef]
- Høgslund, N.; Radutoiu, S.; Krusell, L.; Voroshilova, V.; Hannah, M.A.; Goffard, N.; Sanchez, D.H.; Lippold, F.; Ott, T.; Sato, S. Dissection of symbiosis and organ development by integrated transcriptome analysis of Lotus japonicus mutant and wild-type plants. PLoS One 2009, 4, e6556. [Google Scholar] [CrossRef]
- Lelandais-Briere, C.; Naya, L.; Sallet, E.; Calenge, F.; Frugier, F.; Hartmann, C.; Gouzy, J.; Crespi, M. Genome-wide Medicago truncatula small RNA analysis revealed novel microRNAs and isoforms differentially regulated in roots and nodules. Plant Cell 2009, 21, 2780–2796. [Google Scholar] [CrossRef]
- Takahara, M.; Magori, S.; Soyano, T.; Okamoto, S.; Yoshida, C.; Yano, K.; Sato, S.; Tabata, S.; Yamaguchi, K.; Shigenobu, S. TOO MUCH LOVE, a Novel Kelch repeat-containing F-box protein, functions in the long-distance regulation of the legume–rhizobium symbiosis. Plant Cell Physiol. 2013, 54, 433–447. [Google Scholar]
- De Luis, A.; Markmann, K.; Cognat, V.; Holt, D.B.; Charpentier, M.; Parniske, M.; Stougaard, J.; Voinnet, O. Two microRNAs linked to nodule infection and nitrogen-fixing ability in the legume Lotus japonicus. Plant Physiol. 2012, 160, 2137–2154. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, Z.; Zhu, C. Microarray-based analysis of cadmium-responsive microRNAs in rice (Oryza sativa). J. Exp. Bot. 2011, 62, 3563–3573. [Google Scholar] [CrossRef]
- Ding, D.; Li, W.; Han, M.; Wang, Y.; Fu, Z.; Wang, B.; Tang, J. Identification and characterisation of maize microRNAs involved in developing ears. Plant Biol. 2014, 16, 9–15. [Google Scholar] [CrossRef]
- Uno, Y. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef]
- Shin, D.; Koo, Y.D.; Lee, J.; Lee, H.-J.; Baek, D.; Lee, S.; Cheon, C.-I.; Kwak, S.-S.; Lee, S.Y.; Yun, D.-J. Athb-12, a homeobox-leucine zipper domain protein from Arabidopsis thaliana, increases salt tolerance in yeast by regulating sodium exclusion. Biochem. Biophys. Res. Commun. 2004, 323, 534–540. [Google Scholar] [CrossRef]
- Rodriguez-Uribe, L. A root-specific bZIP transcription factor is responsive to water deficit stress in tepary bean (Phaseolus. acutifolius) and common bean (P. vulgaris). J. Exp. Bot. 2006, 57, 1391–1398. [Google Scholar] [CrossRef]
- Kuo, H.-F.; Chiou, T.-J. The role of microRNAs in phosphorus deficiency signaling. Plant Physiol. 2011, 156, 1016–1024. [Google Scholar] [CrossRef]
- Liang, G.; He, H.; Yu, D. Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PLoS One 2012, 7, e48951. [Google Scholar] [CrossRef]
- Hsieh, L.C.; Lin, S.I.; Shih, A.C.C.; Chen, J.W.; Lin, W.Y.; Tseng, C.Y.; Li, W.H.; Chiou, T.J. Uncovering small rna-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiol. 2009, 151, 2120–2132. [Google Scholar] [CrossRef]
- Xu, F.; Liu, Q.; Chen, L.; Kuang, J.; Walk, T.; Wang, J.; Liao, H. Genome-wide identification of soybean microRNAs and their targets reveals their organ-specificity and responses to phosphate starvation. BMC Genomics 2013, 14, 66. [Google Scholar]
- Smit, P.; Raedts, J.; Portyanko, V.; Debellé, F.; Gough, C.; Bisseling, T.; Geurts, R. NSP1 of the GRAS protein family is essential for rhizobial Nod factor-induced transcription. Science 2005, 308, 1789–1791. [Google Scholar] [CrossRef]
- Murakami, Y.; Miwa, H.; Imaizumi-Anraku, H.; Kouchi, H.; Downie, J.A.; Kawaguchi, M.; Kawasaki, S. Positional cloning identifies Lotus japonicus NSP2, a putative transcription factor of the GRAS family, required for NIN and ENOD40 gene expression in nodule initiation. DNA Res. 2007, 13, 255–265. [Google Scholar]
- Gobbato, E.; Marsh, J.F.; Vernié, T.; Wang, E.; Maillet, F.; Kim, J.; Miller, J.B.; Sun, J.; Bano, S.A.; Ratet, P. A GRAS-type transcription factor with a specific function in mycorrhizal signaling. Curr. Biol. 2012, 22, 2236–2241. [Google Scholar] [CrossRef]
- Maity, S.N.; de Crombrugghe, B. Role of the CCAAT-binding protein CBF/NF-Y in transcription. Trends Biochem. Sci. 1998, 23, 174–178. [Google Scholar] [CrossRef]
- Combier, J.P.; Frugier, F.; de Billy, F.; Boualem, A.; El-Yahyaoui, F.; Moreau, S.; Vernie, T.; Ott, T.; Gamas, P.; Crespi, M.; et al. MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula. Gene. Dev. 2006, 20, 3084–3088. [Google Scholar] [CrossRef]
- Appleby, C.; Bergersen, F. Preparation and experimental use of leghaemoglobin. In Methods for Evaluating Biological Nitrogen Fixation; Bergersen, F.J., Ed.; John Wiley & Sons: Chichester, UK, 1980; pp. 315–335. [Google Scholar]
- Shearer, G.; Kohl, D.H. N2-fixation in field settings: Estimations based on natural 15N abundance. Funct. Plant Biol. 1986, 13, 699–756. [Google Scholar]
- NCBI Genbank Database. Available online: http://www.ncbi.nlm.nih.gov/blast/Blast.cgi (accessed on 15 July 2012).
- Sanger. Available online: http://www.sanger.ac.uk/Software/Rfam (accessed on 15 July 2012).
- Phytozome. Available online: http://www.phytozome.net/ (accessed on 20 July 2012).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, S.; Wang, Y.; Li, K.; Zou, Y.; Chen, L.; Li, X. Identification of Cold-Responsive miRNAs and Their Target Genes in Nitrogen-Fixing Nodules of Soybean. Int. J. Mol. Sci. 2014, 15, 13596-13614. https://doi.org/10.3390/ijms150813596
Zhang S, Wang Y, Li K, Zou Y, Chen L, Li X. Identification of Cold-Responsive miRNAs and Their Target Genes in Nitrogen-Fixing Nodules of Soybean. International Journal of Molecular Sciences. 2014; 15(8):13596-13614. https://doi.org/10.3390/ijms150813596
Chicago/Turabian StyleZhang, Senlei, Youning Wang, Kexue Li, Yanmin Zou, Liang Chen, and Xia Li. 2014. "Identification of Cold-Responsive miRNAs and Their Target Genes in Nitrogen-Fixing Nodules of Soybean" International Journal of Molecular Sciences 15, no. 8: 13596-13614. https://doi.org/10.3390/ijms150813596
APA StyleZhang, S., Wang, Y., Li, K., Zou, Y., Chen, L., & Li, X. (2014). Identification of Cold-Responsive miRNAs and Their Target Genes in Nitrogen-Fixing Nodules of Soybean. International Journal of Molecular Sciences, 15(8), 13596-13614. https://doi.org/10.3390/ijms150813596