Genetic Breeding and Diversity of the Genus Passiflora: Progress and Perspectives in Molecular and Genetic Studies


 and
and
Abstract
:
1. Introduction
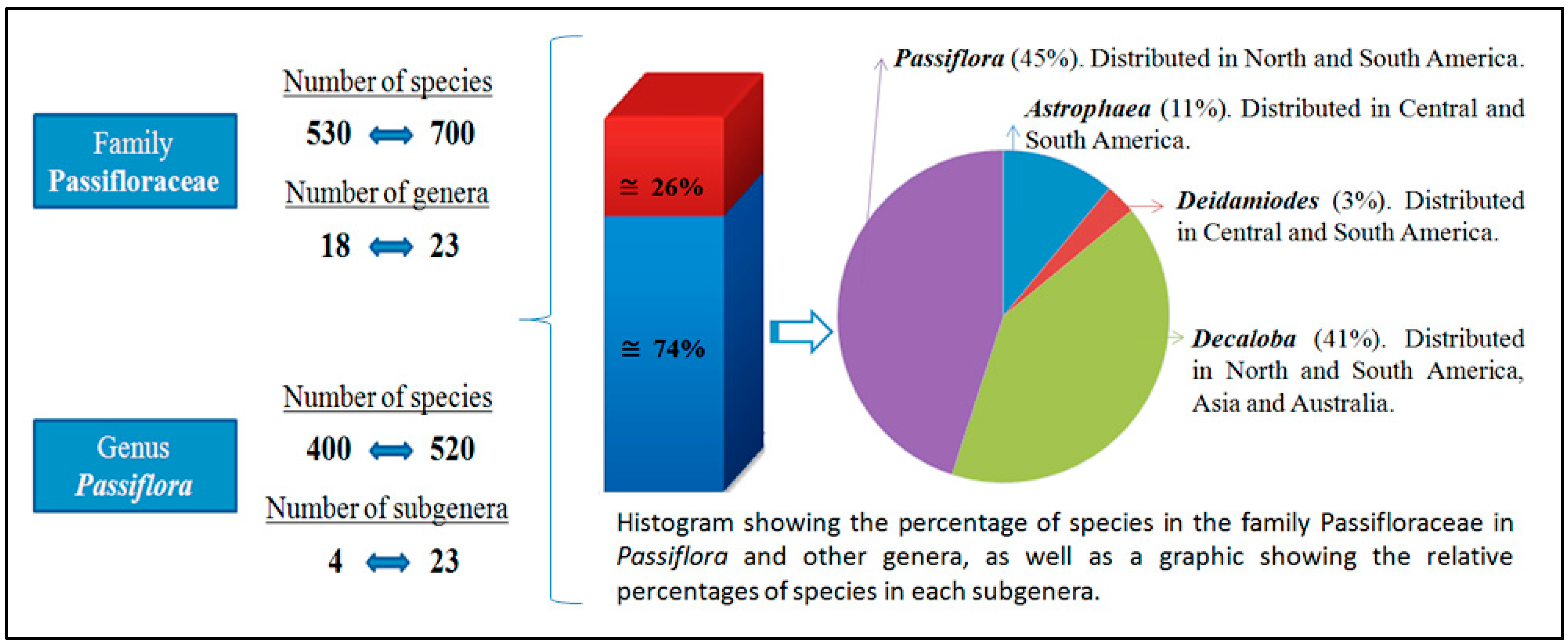
1.1. General Characteristics of the Genus Passiflora, Family Passifloraceae

1.2. Economic Importance of Passion Fruit Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period (Years) | Planted Area (ha × 1000) | Production (t × 1000) | Productivity (t/ha) |
|---|---|---|---|
| 2003 | 34.9 | 485.3 | 13.9 |
| 2004 | 36.6 | 491.6 | 13.4 |
| 2005 | 35.8 | 479.8 | 13.4 |
| 2006 | 44.4 | 615.2 | 13.9 |
| 2007 | 46.8 | 664.3 | 14.2 |
| 2008 | 48.7 | 684.4 | 14.0 |
| 2009 | 50.8 | 718.8 | 14.2 |
| 2010 | 62.0 | 920.2 | 14.8 |
| 2011 | 61.6 | 923.0 | 14.9 |
| 2012 | 57.8 | 776.1 | 13.4 |
| Average (CV) | 48 (21%) | 676 (24%) | 14 (4%) |
2. Contributions of Molecular and Genetic Studies for the Characterization and Use of Passiflora Biodiversity
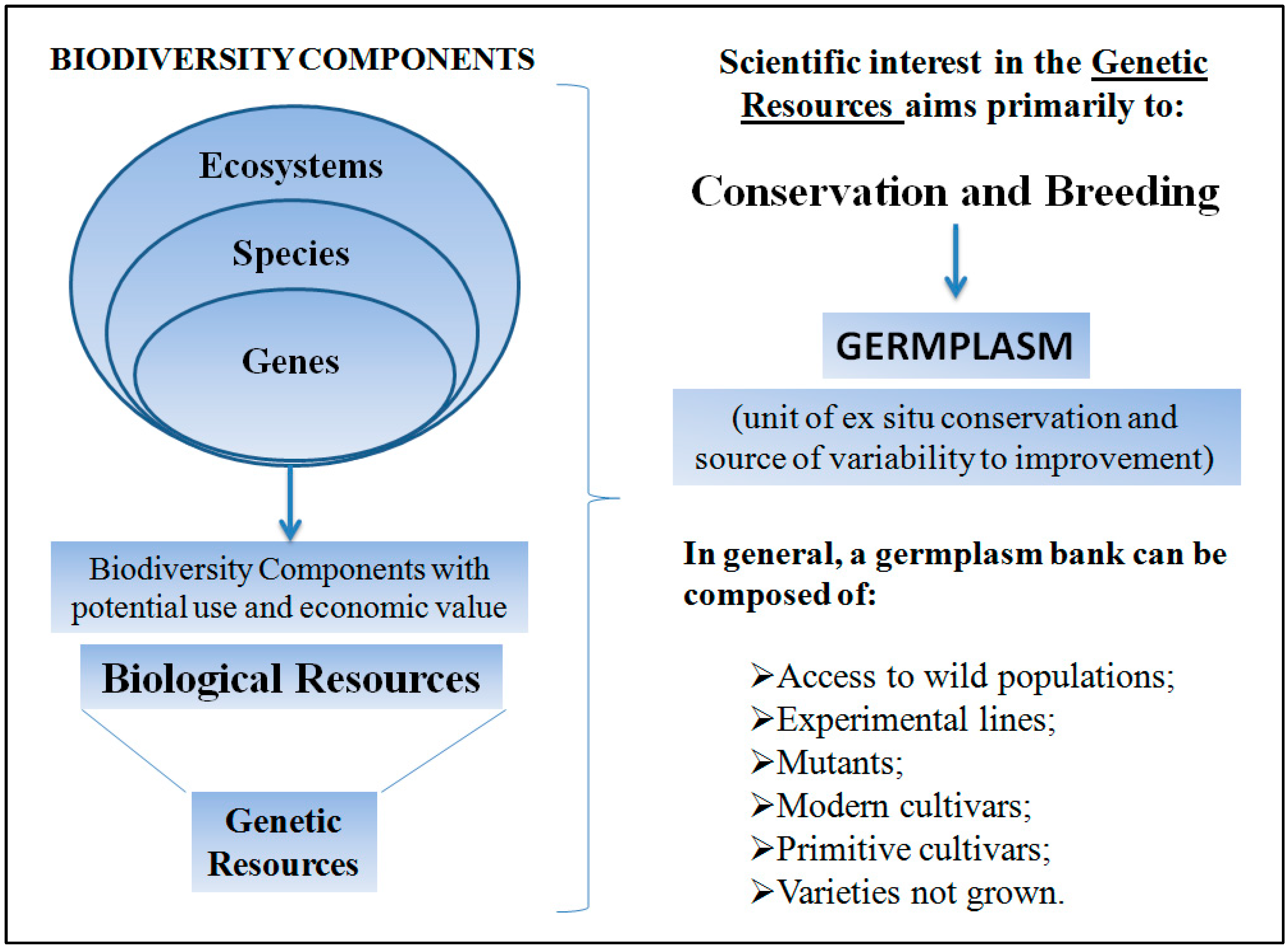
2.1. Biodiversity, Conservation, and Breeding of Passion Fruit


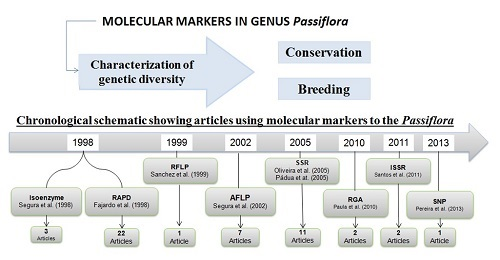
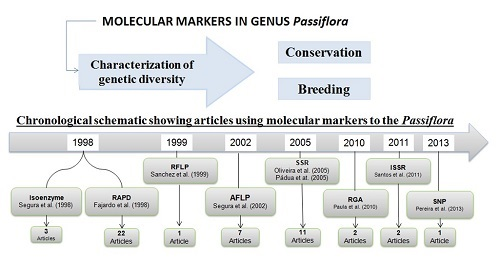
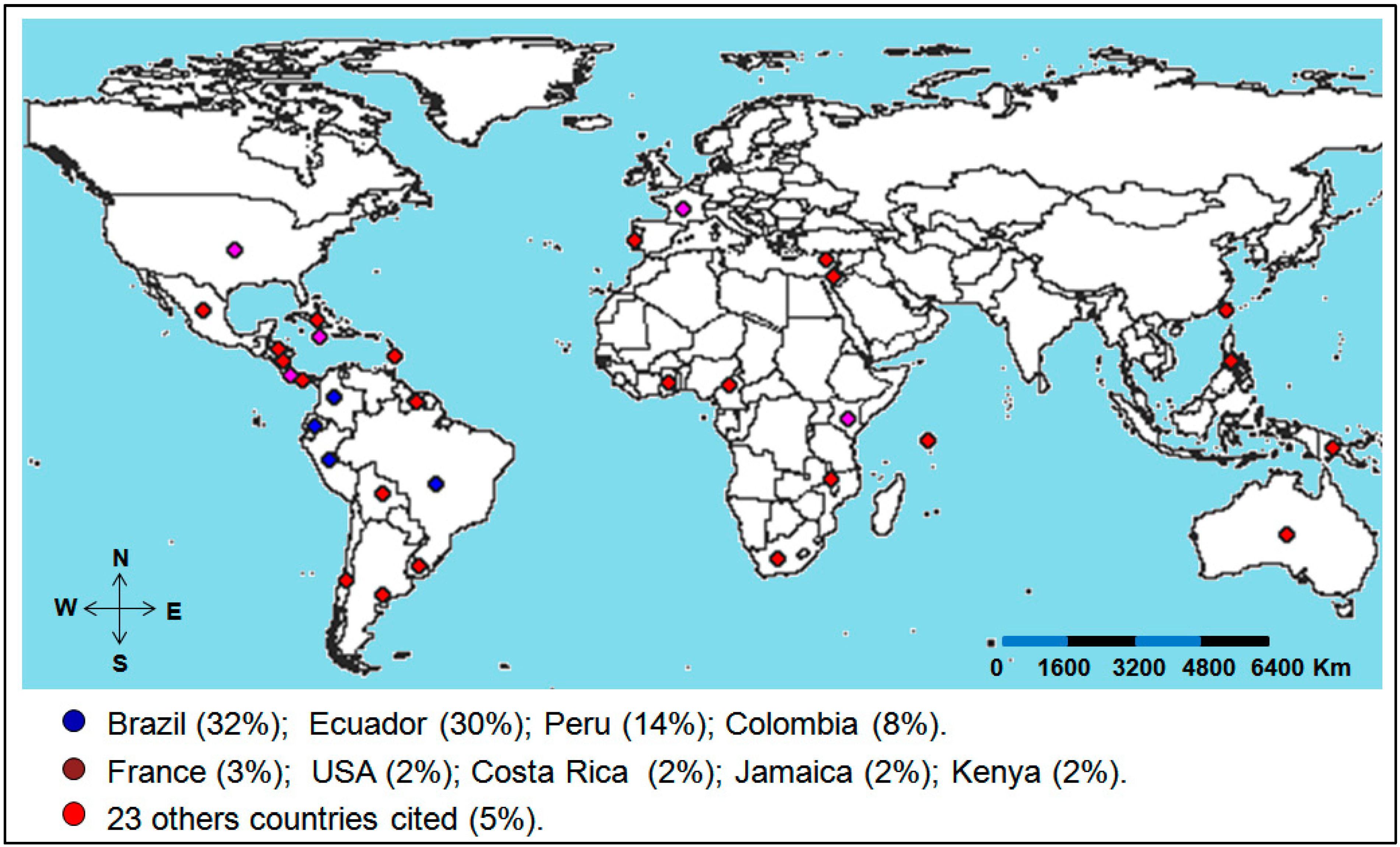
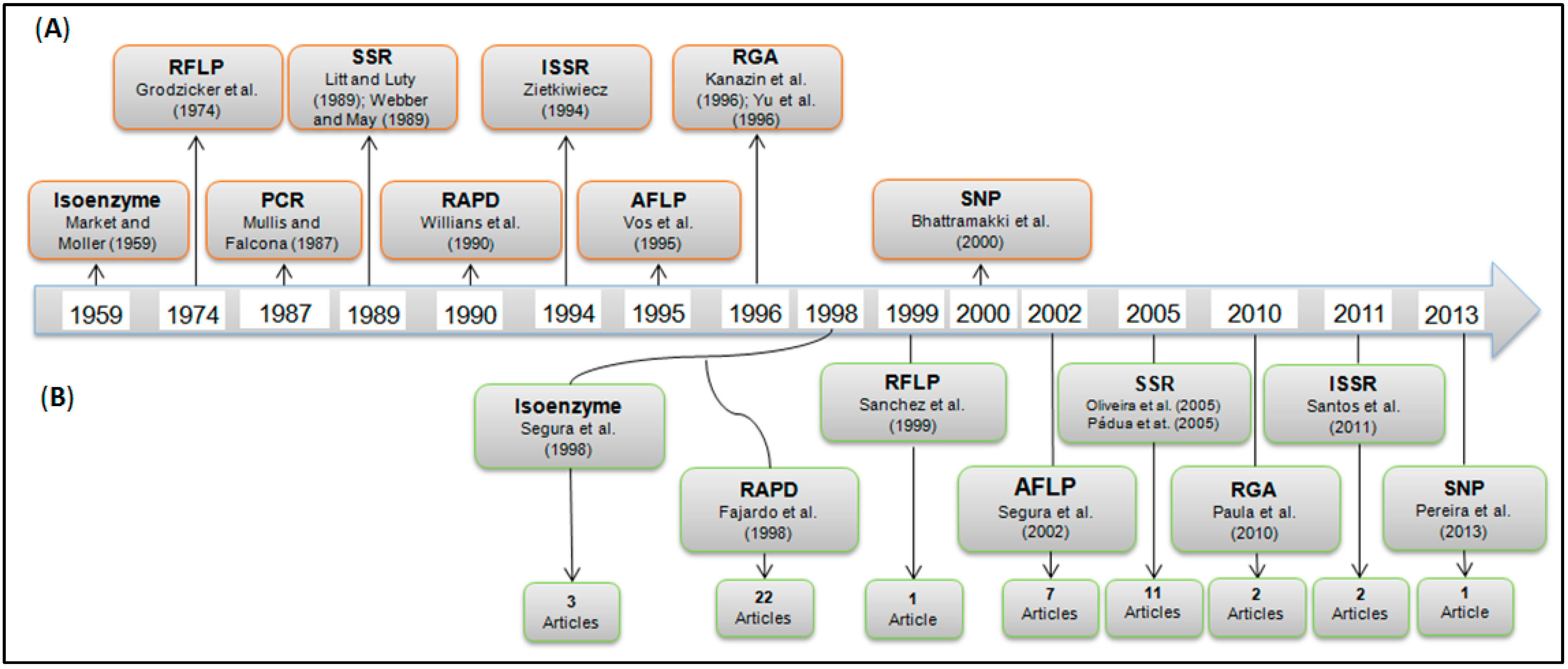
2.2. Application of Molecular Markers for the Characterization of Passiflora Diversity

| Molecular Markers | General Aim of the Article | Evaluated Species | No. of Species | No. of Articles |
|---|---|---|---|---|
| Isozymes | Estimation of diversity (germplasm) | P. ampullacea, P. antioquensis, P. bracteosa, P. cumbalensis, P. manicata, P. mixta, P. pinnatistipula, P. tarminiana, P. tripartita | 10 | 3 |
| RFLP | Estimation of diversity (germplasm) | P. edulis, P. ligulares, P. maliformis, P. caerulea, P. mollissima, P. sp. india, P. cumbalensis, P. antioquiensis, P. pinnatistipula, P. x rosea, P. adenopoda, P. coriacea | 12 | 1 |
| RAPD | Estimation of diversity (germplasm) | P. adenopoda, P. alata, P. amethystina, P. antioquiensis, P. caerulea, P. capsularis, P. cincinnata, P. coccinea, P. coriacea, P. cumbalensis, P. edulis, P. foetida, P. gibertii, P. laurifolia, P. ligularis, P. macrocarpa, P. malacophylla, P. maliformis, P. micropetala, P. mollissima, P. morifolia, P. mucronata, P. nitida, P. palmeri, P. pinnatistipula, P. serrato, P. setacea, P. spinosa, P. suberosa, P. subpeltata, P. trintae, P. vitifolia, P. xrosea | 33 | 15 |
| Characterization and confirmation of hybrids | P. alata, P. gardneri, P. gibertii, P. foetida, P. sublanceolata, P. watsoniana | 6 | 3 | |
| Genetic mapping | P. edulis | 1 | 1 | |
| Other | P. actinia, P. alata, P. amethystina, P. caerulea, P. coccinea, P. eichleriana, P. edulis, P. galbana, P. glandulosa, P. gibertii, P. laurifolia, P. mucronata, P. nitida, P. sidaefolia, P. setacea | 15 | 2 | |
| AFLP | Estimation of diversity (germplasm) | P. alnifolia, P. ampullacea, P. antioquensis, P. bracteosa, P. cumbalensis, P. edulis, P. fimbratistipula, P. gracilens, P. ligularis, P. mixta, P. manicata, P. parritae, P. pinnatistipula, P. popenovii, P. tarminiana, P. tenerifensis, P. tiliaefolia, P. tripartita, P. trifoliata, P. trinervia | 20 | 3 |
| Genetic mapping | P. alata, P. edulis | 2 | 2 | |
| SSR | Estimation of diversity (germplasm) | P. capsularis, P. edulis, P. rubra | 2 | 4 |
| Characterization and confirmation of hybrids | P. foetida, P. sublanceolata | 2 | 1 | |
| Genetic mapping | P. edulis | 1 | 1 | |
| Development, characterization and selection of markers | P. alata, P. cincinnata, Passiflora contracta, P. edulis, P. setacea | 5 | 7 | |
| Cross-amplification | P. caerulea, P. cincinnata, P. edulis, P. foetida, P. gibertii, P. ligularis, P. maliformis, P. mucronata, P. rubra, P. setacea, P. suberosa | 11 | 2 | |
| Other | P. alata | 1 | 1 | |
| RGA | Estimation of diversity (germplasm) | P. caerulea, P. coccinea, P. edulis, P. gibertii, P. nitida, P. odontophyla, P. serratodigitata, P. setacea | 8 | 1 |
| Genetic mapping | P. alata | 1 | 1 | |
| ISSR | Estimation of diversity (germplasm) | P. alata, P. edulis | 2 | 2 |
| SNP | Genetic mapping | P. alata | 1 | 1 |
| Other markers | Estimation of diversity (germplasm) | P. edulis | 1 | 1 |
| Genetic mapping | P. alata | 1 | 1 |
| Species | Characteristics of Microsatellite Loci | References | ||
|---|---|---|---|---|
| Na | H0 | HE | ||
| P. alata | 3.1 | 0.26 | 0.53 | Pádua et al. [82] |
| P. cincinnata | 5 | 0.52 | 0.52 | Cerqueira-Silva et al. [84] |
| P. cincinnata | 3.3 | 0.26 | 0.36 | Cerqueira-Silva et al. [81] |
| P. contracta | 4.9 | 0.53 | 0.61 | Cazé et al. [85] |
| P. edulis | 7.6 | 0.58 | 0.62 | Oliveira et al. [79] |
| P. edulis | 3.4 | 0.31 | 0.36 | Cerqueira-Silva et al. [81] |
| P. setacea | 2.8 | 0.34 | 0.41 | Cerqueira-Silva et al. [81] |
2.3. Application and Perspectives of Genetic Engineering for Passion Fruit Culture
2.4. Contributions and Perspectives of Molecular Biology in Pre- and Post-Breeding Programs
3. Conclusions, Perspectives, and Challenges
Acknowledgments
Supplementary Information
| Molecular Markers | General Aim of the Article | No. of Article | References |
|---|---|---|---|
| Isoenzyme | Estimation of diversity (germplasm) | 3 | [118,119,120] |
| RFLP | Estimation of diversity (germplasm) | 1 | [121] |
| RAPD | Estimation of diversity (germplasm) | 15 | [7,8,58,62,63,80,81,122,123,124,125,126,127,128,129] |
| Characterization and confirmation of hybrids | 3 | [47,72,113] | |
| Genetic mapping | 1 | [115] | |
| Others | 2 | [114,130] | |
| AFLP | Estimation of diversity (germplasm) | 3 | [79,95,131] |
| Genetic mapping | 2 | [84,86] | |
| SSR | Estimation of diversity (germplasm) | 4 | [95,96,97,98] |
| Characterization and confirmation of hybrids | 1 | [47] | |
| Genetic mapping | 1 | [86] | |
| Development, characterization and
selection of markers | 7 | [85,86,87,88,89,90,91] | |
| Cross-amplification | 2 | [87,92] | |
| Others | 1 | [130] | |
| RGA | Estimation of diversity (germplasm) | 1 | [82] |
| Genetic mapping | 1 | [84] | |
| ISSR | Estimation of diversity (germplasm) | 2 | [132,133] |
| SNP | Genetic mapping | 1 | [84] |
| RGA | Estimation of diversity (germplasm) | 1 | [134] |
| Genetic mapping | 1 | [84] |
Conflicts of Interest
References
- Jorgensen, P.M.; MacDougal, J.M. Taxonomic History of Passiflora. Mo. Bot. Gard. Available online: http://www.mobot.org/mobot/research/passiflora/taxonomic_history.shtml (accessed on 14 March 2014).
- Feuillet, C. Passifloraceae (passion flower family). In Flowering Plants of the Neotropics; Smith, N., Mori, S.A., Henderson, A., Stevenson, D.W., Held, S.V., Eds.; Princeton University Press: Oxford, MS, USA, 2004; pp. 286–287. [Google Scholar]
- Watson, L.; Dallwitz, M.J. The Families of Flowering Plants: Descriptions, Illustrations, Identification Retrieval. Available online: http://www.citeulike.org/user/Flit/article/728499 (accessed on 19 December 2012).
- Bernacci, L.C. Passifloraceae. In Flora Fanerogâmica do Estado de São Paulo; Wanderley, M.G.L., Shepherd, G.J., Giulietti, A.M., Melhem, T.S., Eds.; RIMA/FAPESP: São Paulo, Brazil, 2003; pp. 247–248. [Google Scholar]
- MacDougal, J.M.; Feuillet, C. Systematics. In Passiflora: Passionflowers of the World; Ulmer, T., Mac Dougal, J.M., Eds.; Timber Press: Portland, OR, USA, 2004; pp. 27–31. [Google Scholar]
- Barroso, G.M. Passifloraceae. In Rio de Jeneiro: Livros Técnicos e Científicos; Editora da Universidade de São Paulo: São Paulo, Brazil, 1978; Volume 1, pp. 194–197. [Google Scholar]
- Fajardo, D.; Angel, F.; Grum, M.; Tohme, J.; Lobo, M.; Roca, W.M.; Sanchez, I. Genetic variation analysis of the genus Passiflora L. using RAPD markers. Euphytica 1998, 101, 341–347. [Google Scholar] [CrossRef]
- Viana, A.P.; Pereira, T.N.S.; Pereira, M.G.; Souza, M.M.; Maldonado, J.V.M.; Amaral Junior, A.T. Genetic diversity among yellow passion fruit commercial genotypes and among Passiflora species using RAPD. Rev. Bras. Frutic. 2003, 25, 489–493. [Google Scholar] [CrossRef]
- Bernacci, L.C.; Cervi, A.C.; Milward-de-Azevedo, M.A.; Nunes, T.S.; Imig, D.C.; Mezzonato, A.C. Lista de Espécies da Flora do BRASIL. Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB182 (accessed on 20 January 2014). (In Portuguese)
- Ocampo, J.; d’Eeckenbrugge, J.C.; Jarvis, A. Distribution of the genus Passiflora L. diversity in Colombia and its potential as an indicator for biodiversity management in the coffee growing zone. Diversity 2010, 2, 1158–1180. [Google Scholar] [CrossRef] [Green Version]
- Muschner, V.C.; Lorenz, A.P.; Cervi, A.C.; Bonatto, S.L.; Souza-Chies, T.T.; Salzano, F.M.; Freitas, L.B. A first molecular phylogenetic analysis of Passiflora (Passifloracae). Am. J. Bot. 2003, 90, 1229–1238. [Google Scholar] [CrossRef]
- Feuillet, C.; MacDougal, J.M. A new infrageneric classification of Passiflora L. (Passifloraceae). Passiflora 2004, 13, 34–38. [Google Scholar]
- Killip, E.P. Publications field museum of natural history—Botanical series. In American Species of Passifloraceae; Field Museum of Natural History: London, UK, 1938; Volume 19, pp. 1–613. [Google Scholar]
- Escobar, L.K. A new subgenus and five new species in Passiflora (Passifloraceae) from South America. Annu. Mo. Bot. Gard. 1989, 76, 877–885. [Google Scholar] [CrossRef]
- Pádua, J.G. Análise Genetic de Espécies do Gênero Passiflora L. Com Base Em Abordagens Filogenéticas, Morfometricas e em Marcadores Microssatélites. Ph.D. Thesis, Universidade Estadual de São Paulo, Piracicaba, São Paulo, Brazil, July 2004. [Google Scholar]
- Muschner, V.C.; Zamberlan, P.M.; Bonatto, S.L.; Freitas, L.B. Phylogeny, biogeography and divergence times in Passiflora (Passifloraceae). Genet. Mol. Biol. 2012, 35, 1036–1043. [Google Scholar] [CrossRef]
- Freitas, L.B. História evolutiva das espécies de Passiflora L. de ocorrência no Rio Grande do Sul: Aspectos genéticos, estrutura populacional e filogenia. R. Bras. Bioci. 2011, 9, 41–47. (In Portuguese) [Google Scholar]
- Muschner, V.C. Filogenia Molecular, Taxas Evolutivas, Tempo de Divergência e Herança Organelar em Passiflora L. (Passifloraceae). Ph.D. Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil, August 2005. [Google Scholar]
- Torsten, U.; John, M.M. Passiflora:Passionflowers of the World; Ulmer, T., Mac Dougal, J.M., Eds.; Timber Press: Portland Oregon, OR, USA, 2004; pp. 27–31. [Google Scholar]
- Hansen, A.K.; Escobar, L.K.; Gilbert, L.E.; Jansen, R.K. Paternal, maternal, and biparental inheritance of the chloroplast genome in Passiflora (Passifloraceae): Implications for phylogenetic studies. Am. J. Bot. 2007, 94, 42–46. [Google Scholar] [CrossRef]
- Mäder, G.; Zamberlan, P.M.; Fagundes, N.J.R.; Magnus, T.; Salzano, F.M.; Bonatto, S.L.; Freitas, L.B. The use and limits of ITS data in the analysis of intraspecific variation in Passiflora L. (Passifloraceae). Genet. Mol. Biol. 2010, 33, 99–108. [Google Scholar] [CrossRef]
- Cazé, A.L.R.; Mäder, G.; Bonatto, S.L.; Freitas, L.B. A molecular systematic analysis of Passiflora ovalis and Passiflora contracta (Passifloraceae). Phytotaxa 2013, 132, 39–46. [Google Scholar]
- Laiou, A.; Mandolini, L.A.; Piredda, R.; Bellarosa, R.; Simeone, M.C. DNA barcoding as a complementary tool for conservation and valorisation of forest resources. ZooKeys 2013, 365, 197–213. [Google Scholar] [CrossRef]
- Shneyer, V.S. DNA barcoding is a new approach in comparative genomics of plants. Russ. J. Genet. 2009, 45, 1267–1278. [Google Scholar] [CrossRef]
- Wetzel, M.M.V.S.; Gimenes, M.A.; Pádua, J.G.; José, S.C.B.R.; Neto, L.G.P. Conservação de espécies silvestres com potencial de utilização em programas de pré-melhoramento na coleção base da Embrapa. In Pré-melhoramento de Plantas. Estado da Arte e Experiências de Sucesso; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J.F., Faleiro, F.G., Folle, S.M., Guimarães, E.P., Eds.; Embrapa Informações Tecnologicas: Brasilia, Brazil, 2011; pp. 99–122. (In Portuguese) [Google Scholar]
- Vanderplank, J. Passionflowers, 2nd ed.; Cassel: London, UK, 1996. [Google Scholar]
- Ital. In Maracujá: Cultura, Matéria Prima, Processamento e Aspectos Econômicos, 2nd ed.; Teixeira, C.G. (Ed.) ITAL: Campinas, Brazil, 1994; pp. 1–142. (In Portuguese)
- Oliveira, J.C. Melhoramento Genético de P. edulis f. Flavicarpa Deg. Visando Aumento de Produtividade. Ph.D. Thesis, Universidade Estadual de São Paulo, Jaboticabal, São Paulo, Brazil, 1980. [Google Scholar]
- Meletti, L.M.M.; Soares-Scott, M.D.; Bernacci, L.C. Caracterização fenotípica de três seleções de maracujazeiro-roxo (Passiflora edulis Sims). Rev. Bras. Frutic. 2005, 27, 268–272. (In Portuguese) [Google Scholar] [CrossRef]
- Meletti, L.M.M.; Bernacci, L.C.; Soares-Scott, M.D.; Azevedo Filho, J.A.; Martins, A.M. Variabilidade genética em caracteres morfológicos, agronômicos e citogenéticos de populações de maracujazeiro-doce (Passiflora alata Curtis). Rev. Bras. Frutic. 2003, 25, 275–278. (In Portuguese) [Google Scholar] [CrossRef]
- Cerqueira-Silva, C.B.M.; Cardoso-Silva, C.B.; Nonato, J.V.A.; Corrêa, R.X.; Oliveira, A.C. Genetic dissimilarity of “yellow” and “sleep” passion fruit accessions based on the fruits physical-chemical characteristics. Crop. Breed. Appl. Biotechnol. 2009, 9, 210–218. [Google Scholar] [CrossRef]
- Junqueira, N.T.V.; Anjos, J.R.N.; Silva, A.N.P.; Chaves, R.C.; Gomes, A.C. Reaction to diseases and yield of eleven cultivars of sour-passion fruit cultivated with no pesticides. Pesq. Agropec. Bras. 2003, 38, 1005–1010. [Google Scholar] [CrossRef]
- Junqueira, N.T.V.; Braga, M.F.; Faleiro, F.G.; Peixoto, J.R.; Bernacci, L.C. Potencial de espécies silvestres de maracujazeiro como fonte de resistência a de doenças. In Maracujá: Germoplasma e melhoramento genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 79–108. (In Portuguese) [Google Scholar]
- Oliveira, E.J.; Soares, T.L.; Barbosa, C.J.; Santos Filho, H.P.; Jesus, O.N. Severidade de doenças em maracujazeiro para identificação de fontes de resistência em condições de campo. Rev. Bras. Frutic. 2013, 35, 485–492. (In Portuguese) [Google Scholar] [CrossRef]
- Hansen, A.K.; Gibert, L.E.; Simpson, B.B.; Downie, S.R.; Cervi, A.C.; Jansen, R.K. Phylogenetic relationships and chromosome number evolution in Passiflora. Syst. Bot. 2006, 31, 138–150. [Google Scholar] [CrossRef]
- Soares-Scott, M.D.; Meletti, L.M.M.; Bernacci, L.C.; Passos, I.R.S. Citogenética clássica e molecular em passifloras. In Maracujá: Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 210–240. (In Portuguese) [Google Scholar]
- Souza, M.M.; Pereira, T.N.S.; Vieira, M.L.C. Cytogenetic studies in some species of Passiflora L. (Passifloraceae): A review emphasizing Brazilian species. Braz. Arch. Biol. Technol. 2008, 51, 247–258. [Google Scholar] [CrossRef]
- Melo, N.F.; Cervi, A.C.; Guerra, M. Kariology and citotaxonomy of the genus Passiflora L. Plant. Syst. Evol. Berl. 2001, 226, 68–84. [Google Scholar]
- Melo, N.F.; Guerra, M. Variability of the 5S and rDNA sites in Passiflora L. with species with distinct base chromosome numbers. Ann. Bot. 2003, 92, 309–316. [Google Scholar] [CrossRef]
- Cuco, S.M.; Vieira, M.L.C.; Modin, M.; Aguiar-Perecin, M.L.R. Comparative karyotipe analysis of three Passiflora L. species and cytogenetic characterization of somatic hybrids. Caryologia 2005, 58, 220–228. [Google Scholar] [CrossRef]
- Praça, M.M.; Carvalho, R.C.; Marcelino, F.C.; Medonça, M.A.C. Morphological aspects of Passiflora edulis f. flavicarpa chromossomes using acridine orange bading and rDNA-FISH tools. Caryologia 2008, 61, 154–159. [Google Scholar] [CrossRef]
- Penha, H.A. Construção de uma Biblioteca Genômica de Passiflora Edullis f. Flavicarpa Inserida em BACs (Bacterial Artificial Chromosome) e Mapeamento Cromossômico Usando Hibridação in situ Fluorescente. Ph.D. Thesis, Universidade Estadual de São Paulo, Piracicaba, São Paulo, Brazil, July 2012. [Google Scholar]
- Santos, A.A. Exploração de uma Biblioteca Genomica de Passiflora edulis f. flavicarpa por Sequenciamento de BAC-ends. Master Thesis, Universidade Estadual de São Paulo, Piracicaba, São Paulo, Brazil, July 2013. [Google Scholar]
- Yotoko, K.S.C.; Dornelas, M.C.; Togni, P.D.; Fonseca, T.C.; Salzano, F.M.; Bonatto, S.L.; Freitas, L.B. Does variation in genome sizes reflect adaptive or neutral processes? New clues from Passiflora. PLoS One 2011, 6, e18212. [Google Scholar] [CrossRef]
- Peixoto, M. Problemas e perspectivas do maracujá ornamental. In Maracujá: Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 456–464. (In Portuguese) [Google Scholar]
- Embrapa Cerrados—Memória do Lançamento dos Híbridos de Maracujazeiro Ornamental. Available online: http://www.cpac.embrapa.br/lancamentoornamental/ (accessed on 20 May 2014). (In Portuguese)
- Santos, E.A.; Souza, M.M.; Abreu, P.P.; Conceição, L.D.H.C.S.; Araujo, I.S.; Viana, A.P.; Almeida, A.F.; Freitas, J.C.O. Confirmation and characterization of interspecific hybrids of Passiflora L. (Passifloraceae) for ornamental use. Euphytica 2012, 184, 389–399. [Google Scholar] [CrossRef]
- Grupo Passiflora—UESC. Available online: http://www.passifloras.org (accessed on 20 May 2014). (In Portuguese)
- Abreu, P.P.; Souza, M.M.; Santos, E.A.; Pires, M.V.; Pires, M.M.; Almeida, A.F. Passion flower hybrids and their use in the ornamental plant market: Perspectives for sustainable development with emphasis on Brazil. Euphytica 2009, 166, 307–315. [Google Scholar] [CrossRef]
- Costa, A.M.; Tupinambá, D.D.O. Maracujá e suas propriedade medicinais—Estado da arte. In Maracujá: Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 475–508. (In Portuguese) [Google Scholar]
- Amata, R.L.; Otipa, M.J.; Waiganjo, M.; Wabule, M.; Thuranira, E.G.; Erbaugh, M.; Miller, S. Incidence, prevalence and severity of passion fruit fungal diseases in major production regions of Kenya. J. Appl. Biosci. 2009, 20, 1146–1152. [Google Scholar]
- Dhawan, K.; Dhawan, S.; Sharma, A. Passiflora: A review update. J. Ethnopharmacol. 2004, 94, 1–23. [Google Scholar] [CrossRef]
- Zeraik, M.L.; Pereira, C.A.M.; Zuin, V.G.; Yariware, J.H. Maracujá: Um alimento funcional? Rev. Bras. Farmacogn. 2010, 20, 459–471. (In Portuguese) [Google Scholar] [CrossRef]
- Faleiro, F.G.; Junqueira, N.T.V.; Braga, M.F.; Peixoto, J.R. Pré-melhoramento do maracujá. In Pré-Melhoramento de Plantas. Estado da Arte e Experiências de Sucesso; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J.F., Faleiro, F.G., Folle, S.M., Guimarães, E.P., Eds.; Embrapa Informações Tecnologicas: Brasilia, Brazil, 2011; pp. 549–570. (In Portuguese) [Google Scholar]
- Ngan, A.; Conduit, R. A double-blind, placebo-controlled investigation of the effects of Passiflora incarnata (passionflower) herbal tea on subjective sleep quality. Phytother. Res. 2011, 25, 1153–1159. [Google Scholar] [CrossRef]
- Konta, E.M.; Almeida, M.R.; Amaral, C.L.; Darin, J.D.C.; Rosso, V.V.; Mercadante, A.Z.; Antunes, L.M.G.; Bianchi, M.L.P. Evaluation of the antihypertensive properties of yellow passion fruit pulp (Passiflora edulis Sims f. flavicarpa deg.) in spontaneously hypertensive rats. Phytother. Res. 2014, 28, 28–32. [Google Scholar] [CrossRef]
- Souza, J.S.I.; Meletti, L.M.M. Maracujá: Espécies, Variedades, Cultivo; Fealq: Piracicaba, Brazil, 1997; p. 179. (In Portuguese) [Google Scholar]
- Bellon, G.; Faleiro, F.G.; Junqueira, K.P.; Junqueira, N.T.V.; Santos, E.C.; Braga, M.F.; Guimarães, C.T. Variabilidade genética de acessos silvestres e comerciais de Passiflora edulis Sims. com base em marcadores RAPD. Rev. Bras. Frutic. 2007, 29, 124–127. (In Portuguese) [Google Scholar] [CrossRef]
- Ferreira, F.R. Recursos genéticos de Passiflora. In Maracujá: Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 41–50. (In Portuguese) [Google Scholar]
- IBGE—Diretoria de Pesquisas, Coordenação de Agropecuária, Produção Agrícola Municipal, 2012. Available online: http://www.ibge.gov.br/home/estatistica/economia/pam/2012/default_perm_xls.shtm (accessed on 14 March 2014). (In Portuguese)
- Meletti, L.M.M.; Soares-Scott, M.D.; Bernacci, L.C.; Passos, I.R.S. Melhoramento genético do maracujá: passado e futuro. In Maracujá: Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 55–78. (In Portuguese) [Google Scholar]
- Cerqueira-Silva, C.B.M.; Conceição, L.D.H.C.S.; Santos, E.S.L.; Cardoso-Silva, C.B.; Pereira, A.S.; Oliveira, A.C.; Corrêia, R.X. Genetic variability in wild genotypes of Passiflora cincinnata based on RAPD markers. Genet. Mol. Res. 2010, 9, 2421–2428. [Google Scholar] [CrossRef]
- Cerqueira-Silva, C.B.M.; Santos, E.S.L.; Conceição, L.D.H.C.S.; Cardoso-Silva, C.B.; Pereira, A.S.; Oliveira, A.C.; Corrêa, R.X. Short communication genetic variation in a wild population of the sleep passion fruit (Passiflora setacea) based on molecular markers. Genet. Mol. Res. 2012, 11, 731–738. [Google Scholar] [CrossRef]
- Pereira, M.G.; Pereira, T.N.S.; Viana, A.P. Marcadores moleculares aplicados ao melhoramento genético do maracujazeiro. In Maracujá: Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 275–292. (In Portuguese) [Google Scholar]
- Meletti, L.M.M.; Santos, R.R.; Minami, K. Melhoramento do maracujazeiro-amarelo: Obtenção do cultivar “composto IAC-27” 1,2. Sci. Agric. 2000, 57, 491–498. (In Portuguese) [Google Scholar] [CrossRef]
- Rao, V.R.; Hodgkin, T. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Organ Cult. 2002, 68, 1–19. [Google Scholar]
- Nass, L.L. Pré-melhoramento vegetal. In Pré-melhoramento de Plantas. Estado da Arte e Experiências de Sucesso; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J.F., Faleiro, F.G., Folle, S.M., Guimarães, E.P., Eds.; Embrapa Informações Tecnologicas: Brasilia, Brazil, 2011; pp. 23–38. (In Portuguese) [Google Scholar]
- Da Silva, A.R.; Fávero, A.P.; Alves, A.A.C.; Sartorato, A.; da Silva Noronha, A.C.; Albuquerque, A.C.S.; de Souza Peñaloza, A.D.P.; da Silva Souza, A.; Brondani, C.; Pacheco, C.A.P.; et al. Pré-Melhoramento de Plantas. Estado da Arte e Experiências de Sucesso; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J.F., Faleiro, F.G., Folle, S.M., Guimarães, E.P., Eds.; Embrapa Informações Tecnologicas: Brasilia, Brazil, 2011; pp. 1–614. (In Portuguese) [Google Scholar]
- SCOPUS Database. Available online: http://www.scopus.com (accessed on 20 May 2014).
- Thomson Reuters Web of Science (formerly ISI Web of Knowledge). Available online: http://apps.webofknowledge.com (accessed on 20 May 2014).
- Maluf, W.R.; Silva, J.R.; Grattapaglia, D.; Toma-Braghini, M.; Corte, R.D.; Machado, M.A.; Caldas, L.S. Genetic gains via clonal selection in passion fruit Passiflora edulis Sims. Rev. Bras. Genet. 1989, 12, 833–841. [Google Scholar]
- Junqueira, K.P.; Faleiro, F.G.; Junqueira, N.T.V.; Bellon, G.; Ramos, J.D.; Braga, M.F.; Souza, L.S. Confirmação de híbridos interespecíficos artificiais no gênero Passiflora por meio de marcadores RAPD. Rev. Bras. Frutic. 2008, 30, 191–196. (In Portuguese) [Google Scholar] [CrossRef]
- Junqueira, N.T.V.; Santos, E.C.; Junqueira, K.P.; Faleiro, F.G.; Bellon, G.; Braga, M.F. Características físico-químicas e produtividade de acessos de Passiflora nitida Kunth procedentes do Centro-Norte do Brasil. Rev. Bras. Frutic. 2010, 32, 791–797. (In Portuguese) [Google Scholar] [CrossRef]
- Cerqueira-Silva, C.B.M.; Moreira, C.N.; Figueira, A.R.; Corrêa, R.X.; Oliveira, A.C. Detection of resistance gradient to the passion fruit woodiness virus and selection of yellow passion fruit plants in field conditions. Genet. Mol. Res. 2008, 7, 1209–1216. [Google Scholar] [CrossRef]
- Silva, A.S.; Oliveira, E.J.; Haddad, F.; Laranjeira, F.F.; Jesus, O.N.; Oliveira, S.A.S.; Costa, M.A.P.C.; Freitas, J.P.X. Identification of passion fruit genotypes resistant to Fusarium oxysporum f. sp. passiflorae. Trop. Plant Pathol. 2013, 38, 236–242. [Google Scholar]
- Faleiro, F.G.; Junqueira, N.T.V.; Braga, M.F.; Junqueira, K.P.; Bellon, G.; Fonseca, K.G.; Peixoto, J.R. Cruzamentos inter-específicos e retrocruzamentos visando à resistência do maracujazeiro a doenças. In IV Congresso Brasileiro de Melhoramento de Plantas; 2007; Volume 4. (In Portuguese) [Google Scholar]
- Fonseca, K.G.; Faleiro, F.G.; Bellon, G.; Junqueira, K.P.; Junqueira, N.T.V.; Braga, M.F.; Peixoto, J.R. Caracterização de plantas RC4 e recuperação do genoma recorrente com base em marcadores RAPD. In IV Congresso Brasileiro de Melhoramento de Plantas; Repositório Institucional: Brasília, Brazil, 2007. (In Portuguese) [Google Scholar]
- NCBI—National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 20 May 2014).
- Ganga, R.M.D.; Ruggiero, C.; Lemos, E.G.M.; Grili, G.V.G.; Gonçalves, M.M.; Chagas, E.A.; Wickert, E. Diversidade genética em maracujazeiro-amarelo utilizando marcadores moleculares fAFLP. Rev. Bras. Frutic. 2004, 26, 494–498. (In Portuguese) [Google Scholar] [CrossRef]
- Cerqueira-Silva, C.B.M.; Conceição, L.D.H.C.S.; Cardoso-Silva, C.B.; Pereira, A.S.; Santos, E.S.L.; Oliveira, A.C.; Correa, R.X. Genetic diversity of yellow passion fruit (Passiflora edulis Sims) based on RAPD markers. Crop. Breed. Appl. Biotechnol. 2010, 10, 154–159. [Google Scholar] [CrossRef]
- Junqueira, K.P.; Faleiro, F.G.; Ramos, J.D.; Bellon, G.; Junqueira, N.T.V.; Braga, M.F. Variabilidade genética de acessos de maracujá-suspiro com base em marcadores moleculares. Rev. Bras. Frutic. 2007, 29, 571–575. (In Portuguese) [Google Scholar] [CrossRef]
- Paula, M.S.; Fonseca, M.E.N.; Boiteux, L.S.; Peixoto, J.R. Genetic characterization of Passiflora species via resistance genes analog markers. Rev. Bras. Frutic. 2010, 32, 222–229. [Google Scholar] [CrossRef]
- Gonçalves, Z.S. Genetic Diversity in Passion Fruit (Passiflora spp.) with Use of the Resistance Gene Analogs Markers (RGA) Aiming the Genetic Improvement. Graduation Thesis, Universidade Estadual do Sudoeste da Bahia, Itapetinga, Bahia, Brazil, September 2013. [Google Scholar]
- Pereira, G.S.; Nunes, E.S.; Laperuta, L.D.C.; Braga, M.F.; Penha, H.A.; Diniz, A.L.; Munhoz, C.F.; Gazaffi, R.; Garcia, A.A.F.; Vieira, M.L.C. Molecular polymorphism and linkage analysis in sweet passion fruit, an outcrossing species. Ann. Appl. Biol. 2013, 162, 347–361. [Google Scholar] [CrossRef]
- Oliveira, E.J.; Pádua, J.G.; Zucchi, M.I.; Camargo, L.E.A.; Fungaro, M.H.P.; Vieira, M.L.C. Development and characterization of microsatellite markers from the yellow passion fruit (Passiflora edulis f. flavicarpa). Mol. Ecol. Notes 2005, 5, 331–333. [Google Scholar] [CrossRef]
- Oliveira, E.J.; Vieira, M.L.C.; Garcia, A.A.F.; Munhoz, C.E.F.; Margarido, G.R.A.; Consoli, L.; Matta, F.P.; Moraes, M.C. An integrated molecular map of yellow passion fruit based on simultaneous maximum-likelihood estimation of linkage and linkage phases. J. Am. Soc. Hortic. Sci. 2008, 133, 35–41. [Google Scholar]
- Cerqueira-Silva, C.B.M.; Santos, E.S.L.; Vieira, J.G.P.; Mori, G.M.; Jesus, O.N.; Correa, R.X.; Souza, A.P. New microsatellite markers for wild and commercial species of Passiflora (Passifloraceae) and cross-amplification. Appl. Plant Sci. 2014, 2, 1–5. [Google Scholar]
- Pádua, J.G.; Oliveira, E.J.; Zucchi, M.I.; Oliveira, G.C.X.; Camargo, L.E.A.; Vieira, M.L.C. Isolation and characterization of microsatellite markers from the sweet passion fruit (Passiflora alata Curtis: Passifloraceae). Mol. Ecol. Notes 2005, 5, 863–865. [Google Scholar] [CrossRef]
- Penha, H.A.; Pereira, G.S.; Zucchi, M.I.; Diniz, A.L.; Vieira, M.L.C.; Flachowsky, H. Development of microsatellite markers in sweet passion fruit, and identification of length and conformation polymorphisms within repeat sequences. Plant Breed. 2013, 132, 731–735. [Google Scholar] [CrossRef]
- Cerqueira-Silva, C.B.M.; Santos, E.S.L.; Souza, A.M.; Mori, G.M.; Oliveira, E.J.; Corrêa, R.X.; Souza, A.P. Development and characterization of microsatellite markers for the wild South American Passiflora cincinnata (Passifloraceae). Am. J. Bot. 2012, 99, e170–e172. [Google Scholar] [CrossRef]
- Cazé, A.L.R.; Kriedt, R.A.; Beheregaray, L.B.; Bonatto, S.L.; Freitas, L.B. Isolation and characterization of microsatellite markers for Passiflora contracta. Int. J. Mol. Sci. 2012, 13, 11343–11348. [Google Scholar] [CrossRef]
- Oliveira, G.A.F.; Pádua, J.G.; Costa, J.L.; Jesus, O.N.; Carvalho, F.M.; Oliveira, E.J. Cross-species amplification of microsatellite loci developed for Passifloraedulis Sims. in related Passiflora Species. Braz. Arch. Biol. Technol. 2013, 56, 785–792. [Google Scholar] [CrossRef]
- Silva, M.A.A.; Souza, M.M.; Silva, G.S.; Melo, C.A.F.; Correa, R.X.; Araujo, I.S.; Conceição, L.D.H.C.S. Analysis of transferability of microsatellite primers (SSR) in five wild Passiflora species and intraspecific genetic diversity in P. alata. Genet. Mol. Res. 2013. accepted article. [Google Scholar]
- Paiva, C.L. Descritores Morfológicos e Marcadores Microssatélites na Caracterização de Germoplasma de Passiflora spp. Master Thesis, Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos dos Goytacazes, Rio de Janeiro, Brazil, April 2013. [Google Scholar]
- Ortiz, D.C.; Bohórquez, A.; Duque, M.C.; Tohme, J.; Cuéllar, D.; Vásquez, T.M. Evaluating purple passion fruit (Passiflora edulis Sims f. edulis) genetic variability in individuals from commercial plantations in Colombia. Genet. Resour. Crop. Evol. 2012, 59, 1089–1099. [Google Scholar]
- Amorim, J.S.; Souza, M.M.; Viana, A.J.C.; Correa, R.X.; Araujo, I.S.; Ahnert, D. Cytogenetic, molecular and morphological characterization of Passiflora capsularis L. and Passiflora rubra L. Plant. Syst. Evol. 2014, 300, 1147–1162. [Google Scholar]
- Reis, R.V.; Oliveira, E.J.; Viana, A.P.; Pereira, T.N.S.; Pereira, M.G.; Silva, M.G.M. Diversidade genética em seleção recorrente de maracujazeiro-amarelo detectada por marcadores microssatélites. Pesq. Agropec. Bras. 2011, 46, 51–57. (In Portuguese) [Google Scholar] [CrossRef]
- Reis, R.V.; Viana, A.P.; Oliveira, E.J.; Silva, M.G.M. Phenotypic and molecular selection of passion fruit progenies in the second cycle of recurrent selection. Crop. Breed. Appl. Biotechnol. 2012, 12, 17–24. [Google Scholar]
- Alves, E.O.S.; Cerqueira-Silva, C.B.M.; Souza, A.M.; Santos, C.A.F.; Lima Neto, F.P.; Corrêa, R.X. Short communication comparison of efficiency of distance measurement methodologies in mango (Mangifera indica) progenies based on physicochemical descriptors. Genet. Mol. Res. 2012, 11, 591–596. [Google Scholar] [CrossRef]
- Sesli, M.; Yegenoglu, E.D. Comparison of similarity coefficients used for cluster analysis based on RAPD markers in wild olives. Genet. Mol. Res. 2010, 9, 2248–2253. [Google Scholar] [CrossRef]
- Balestre, M.; Von Pinho, R.G.; Souza, J.C.; Lima, J.L. Comparison of maize similarity and dissimilarity genetic coefficients based on microsatellite markers. Genet. Mol. Res. 2008, 7, 695–705. [Google Scholar]
- Gonçalves, L.S.; Rodrigues, R.; Amaral, A.T.; Karasawa, M.; Sudré, C.P. Comparison of multivariate statistical algorithms to cluster tomato heirloom accessions. Genet. Mol. Res. 2008, 7, 1289–1297. [Google Scholar]
- Cerqueira-Silva, C.B.M.; Cardoso-Silva, C.B.; Conceicao, L.D.H.C.S.; Nonato, J.V.A.; Oliveira, A.C.; Correa, R.X. Comparison of coefficients and distance measurements in passion fruit plants based on molecular markers and physicochemical descriptors. Genet. Mol. Res. 2009, 8, 870–879. [Google Scholar]
- Vieira, M.L.C.; Oliveira, E.J.; Matta, F.P.; Pádua, J.G.; Monteiro, M. Métodos biotecnológicos aplicados ao melhoramento genético do maracujá. In Maracujá:Germoplasma e Melhoramento Genético; Faleiro, F.G., Junqueira, N.T.V., Braga, M.F., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2005; pp. 410–454. (In Portuguese) [Google Scholar]
- Alfenas, P.F.; Braz, A.S.K.; Torres, L.B.; Santana, E.N.; Nascimento, A.V.S.; Carvalho, M.G.; Otoni, W.C.; Zerbini, F.M. Transgenic passion fruit expressing RNA derived from Cowpea aphid-borne mosaic virus is resistant to passion fruit woodiness disease. Fitopatol. Bras. 2005, 30, 33–38. [Google Scholar] [CrossRef]
- Trevisan, F.; Maciel, S.C.; Mendes, B.M.J.; Vieira, M.L.C.; Rezende, J.A.M.; Meletti, L.M.M. Resistance to passion fruit woodiness virus in transgenic passion flower expressing the virus coat protein gene. Plant Dis. 2006, 90, 1026–1030. [Google Scholar] [CrossRef]
- Monteiro-Hara, A.C.B.A.; Jadão, A.S.; Mendes, B.M.J.; Rezende, J.A.M.; Trevisan, F.; Mello, A.P.O.A.; Vieira, M.L.C.; Meletti, L.M.M.; Piedade, S.M.S. Genetic transformation of passionflower and evaluation of r1 and r2 generations for resistance to Cowpea aphid borne mosaic virus. Plant Dis. 2011, 95, 1021–1025. [Google Scholar] [CrossRef]
- Freitas, D.S.; Coelho, M.C.F.; Souza, M.T.; Marques, A.; Ribeiro, B.M. Introduction of the anti-apoptotic baculovirus p35 gene in passion fruit induces herbicide tolerance, reduced bacterial lesions, but does not inhibits passion fruit woodiness disease progress induced by cowpea aphid-borne mosaic virus (CABMV). Biotechnol. Lett. 2007, 29, 79–87. [Google Scholar] [CrossRef]
- Nascimento, A.V.S. Estudo de Isolados Brasileiros de Potuvirus Causadores de Endurecimento dos Frutos do Maracujazeiro e Análise da Resistência à Doença em Linhagens Transgênicas R1 de Maracujá-Amarelo. Ph.D. Thesis, Universidade Federal de Viçosa, Minas Gerais, Brazil, August 2006. [Google Scholar]
- Monteiro, M. Transformação Genetica de Maracujá Amarelo Visando Resistência à Xanthomonasaxonopodis PV. Passiflorae. Ph.D. Thesis, Universidade Estadual de São Paulo, Piracicaba, São Paulo, Brazil, April 2005. [Google Scholar]
- Pino, A.P.C. Transformação Genética de Maracujazeiro (Passiflora Alata Curtis) Para Resistência ao Cowpea Aphid-Borne Mosaic Virus (CABMV); Universidade Estadual de São Paulo: Piracicaba, São Paulo, Brazil, August 2010. (In Portuguese) [Google Scholar]
- Kirk, H.; Freeland, J.R. Applications and implications of neutral versus non-neutral markers in molecular ecology. Int. J. Mol. Sci. 2011, 12, 3966–3988. [Google Scholar] [CrossRef]
- Conceição, L.D.H.C.S.; Belo, G.O.; Souza, M.M.; Santos, S.F.; Cerqueira-Silva, C.B.M.; Correa, R.X. Confirmation of cross-fertilization using molecular markers in ornamental passion flower hybrids. Genet. Mol. Res. 2011, 10, 47–52. [Google Scholar] [CrossRef]
- Fonseca, K.G.; Faleiro, F.G.; Peixoto, J.R.; Junqueira, N.T.V.; Silva, M.S.; Bellon, G.; Junqueira, K.P.; Vaz, C.F. Análise da recuperação do genitor recorrente em maracujazeiro-azedo por meio de marcadores RAPD. Rev. Bras. Frutic. 2009, 31, 145–153. (In Portuguese) [Google Scholar] [CrossRef]
- Carneiro, M.S.; Camargo, L.E.A.; Coeloho, A.S.G.; Vencovsky, R.; Leite, R.P.; Stenzel, N.M.; Vieira, M.L.C. RAPD-based genetic linkage maps of yellow passion fruit (Passiflora edulis Sims. f. flavicarpa Deg.). Genome 2002, 45, 670–678. [Google Scholar] [CrossRef]
- Lopes, R.; Lopes, M.T.G.; Carneiro, M.S.; Matta, F.P.; Camargo, L.E.A.; Vieira, M.L.C. Linkage and mapping of resistance genes to Xanthomonas axonopodis pv. passiflorae in yellow passion fruit. Genome 2006, 49, 17–29. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS One 2011, 6, e19379. [Google Scholar]
- Segura, S.D.; Coppens d’Eeckenbrugge, G.; Ollitrault, P. Isozyme variation in five species of Passflora subgenus Tacsonia and Passiflora manicata. Proc. Interam.Soc. Trop. Hort. 1998, 42, 260–266. [Google Scholar]
- Segura, S.D.; Coppens d’Eeckenbrugge, G.; Ocampo, C.H.; Ollitrault, P. Isozyme variation in Passiflora subgenera Tacsonia and Manicata. Relationships between cultivated and wild species. Genet. Resour. Crop. Evol. 2003, 50, 417–427. [Google Scholar] [CrossRef]
- Segura, S.D.; Coppens d’Eeckenbrugge, G.; Ocampo, C.H.; Ollitrault, P. Isozyme variation in Passiflora subgenus Tacsonia: Geographic and interspecific differentiation among the three most common species. Genet. Resour. Crop. Evol. 2005, 52, 455–463. [Google Scholar] [CrossRef]
- Sánchez, I.; Angel, F.; Grum, M.; Duque, M.C.; Lobo, M.; Thome, J.; Roca, W. Variability of chloroplast DNA in the genus Passiflora L. Euphytica 1999, 106, 15–26. [Google Scholar] [CrossRef]
- Aukar, A.P.A.; Lemos, E.G.M.; Oliveira, J.C.O. Genetic variations among passion fruit species using RAPD markers. Rev. Bras. Frutic. 2002, 24, 738–740. [Google Scholar] [CrossRef]
- Bellon, G.; Faleiro, F.G.; Peixoto, J.R.; Junqueira, K.P.; Junqueira, N.T.V.; Fonsceca, K.G.; Braga, M.F. Variabilidade genética de acessos obtidos de populações cultivadas e silvestres de maracujazeiro-doce com base em marcadores rapd. Rev. Bras. Frutic 2009, 31, 197–202. (In Portuguese) [Google Scholar] [CrossRef]
- Castro, A.P.G.; Faleiro, F.G.; Carvalho, D.D.C.; Fonseca, K.G.; Vilela, M.F.; Junqueira, N.T.V.; Cares, J.E. Genetic variability of Passiflora spp. from commercial fields in the Federal District, Brazil. Ciência Rural 2011, 41, 996–1002. [Google Scholar]
- Cerqueira-Silva, C.B.M.; Cardoso-Silva, C.B; Santos, E.S.L.; Conceição, L.D.H.C.S.; Pereira, A.S.; Oliveira, A.C.; Corrêa, R.X. Genetic diversity in wild species of passion fruit (Passiflora trintae) based on molecular markers. Genet. Mol. Res. 2010, 9, 2123–2130. [Google Scholar]
- Crochemore, M.L.; Molinari, H.B.; Stenzel, M.N. Agro-morphological characterization of passion fruit (Passiflora spp.) germplasm. Rev. Bras. Frutic. 2003, 25, 5–10. (In Portuguese) [Google Scholar] [CrossRef]
- Pérez-Almeida, I.; Vásquez García, S.; Pérez, D.; Rosa, O.; Salazar, E. Huella genetic de genotipos silvestres y comerciales de Passiflora spp. utilizando patrones RAPD. Bioagro 2009, 21, 203–208. (In Spanish) [Google Scholar]
- Pérez-Almeida, I.; Vásquez García, S.; Pérez, D.; Salazar, E. Genetic diversity in six species of Passiflora spp. using RAPD Introducción. Rev. Fac. Agron. 2010, 27, 347–359. [Google Scholar]
- Viana, A.J.C.; Souza, M.M.; Araújo, I.S.; Corrêa, R.X.; Ahnert, D. Genetic diversity in Passifloraspecies determined by morphological and molecular characteristics. Biologia Plantarum 2010, 54, 535–538. [Google Scholar] [CrossRef]
- Ferreira, T.G.T.; Penha, H.A.; Zucchi, M.I.; Santos, A.A.; Hanai, L.R.; Junqueira, N.; Braga, N.F.; Vencovsky, R.; Vieira, M.L.C. Outcrossing rate in sweet passion fruit based on molecular markers. Plant Breed. 2010, 129, 727–730. [Google Scholar] [CrossRef]
- Segura, S.D.; Coppens d’Eeckenbrugge, G.; Bohorquez, A.; Ollitrault, P.; Tohmé, J. An AFLP diversity study of the genus Passiflora focusing on subgenus Tacsonia. Genet. Resour. Crop. Evol. 2002, 49, 111–123. [Google Scholar] [CrossRef]
- Costa, J.L.; Jesus, O.N.; Oliveira, G.A.F.; Oliveira, E.J. Effect of selection on genetic variability in yellow passion fruit. Crop. Breed. Appl. Biotechnol. 2012, 12, 253–260. [Google Scholar]
- Santos, L.F.; Oliveira, E.J.; Silva, A.S.; Carvalho, S.M.; Costa, J.L.; Pádua, J.C. ISSR markers as a tool for the assessment of genetic diversity in passiflora. Biochem. Genet. 2011, 49, 540–554. [Google Scholar] [CrossRef]
- Fonseca-Trujillo, N.; Márquez-Cardona, M.P.; Moreno-Osorio, J.H.; Terán-Pérez, W.; Schuler-García, I. Caracterización molecular de materiales cultivados de gulupa (Passiflora edulis f. edulis). Univ. Sci. 2009, 14, 135–140. (In Spanish) [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cerqueira-Silva, C.B.M.; Jesus, O.N.; Santos, E.S.L.; Corrêa, R.X.; Souza, A.P. Genetic Breeding and Diversity of the Genus Passiflora: Progress and Perspectives in Molecular and Genetic Studies. Int. J. Mol. Sci. 2014, 15, 14122-14152. https://doi.org/10.3390/ijms150814122
Cerqueira-Silva CBM, Jesus ON, Santos ESL, Corrêa RX, Souza AP. Genetic Breeding and Diversity of the Genus Passiflora: Progress and Perspectives in Molecular and Genetic Studies. International Journal of Molecular Sciences. 2014; 15(8):14122-14152. https://doi.org/10.3390/ijms150814122
Chicago/Turabian StyleCerqueira-Silva, Carlos Bernard M., Onildo N. Jesus, Elisa S. L. Santos, Ronan X. Corrêa, and Anete P. Souza. 2014. "Genetic Breeding and Diversity of the Genus Passiflora: Progress and Perspectives in Molecular and Genetic Studies" International Journal of Molecular Sciences 15, no. 8: 14122-14152. https://doi.org/10.3390/ijms150814122
APA StyleCerqueira-Silva, C. B. M., Jesus, O. N., Santos, E. S. L., Corrêa, R. X., & Souza, A. P. (2014). Genetic Breeding and Diversity of the Genus Passiflora: Progress and Perspectives in Molecular and Genetic Studies. International Journal of Molecular Sciences, 15(8), 14122-14152. https://doi.org/10.3390/ijms150814122