High-Resolution Imaging Reveals New Features of Nuclear Export of mRNA through the Nuclear Pore Complexes

Abstract
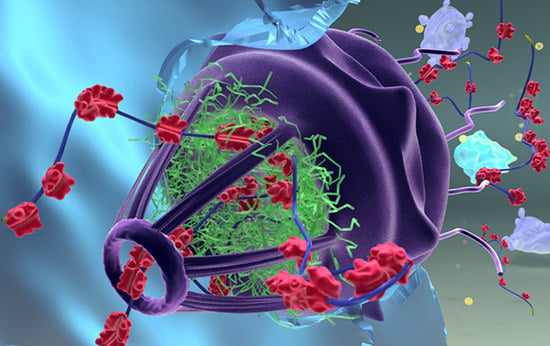
:
1. Introduction
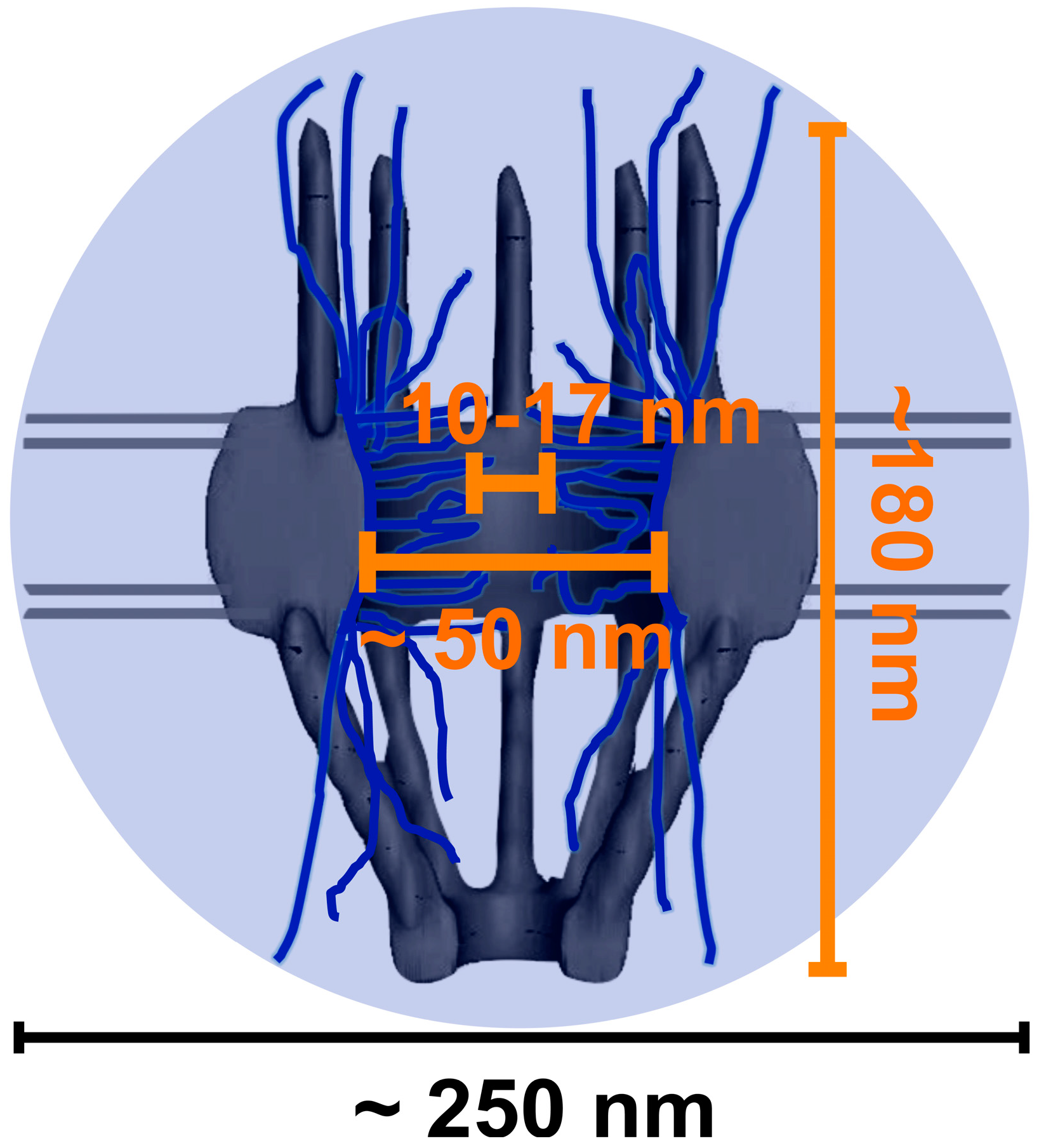
2. A High-Resolution Microscopy Imaging Approach Is Needed to Look Inside the Nuclear Pore Complex (NPC)


{kind=link}
{kind=link}
{kind=link}
| Imaging Method | Cell Type | RNA Type (Size in kb) | Illumination Area | Spatiotemporal Resolution | Major Conclusions | Reference |
|---|---|---|---|---|---|---|
| Wide-field epi-fluorescence microscopy | U2OS | Human (4.8, 8, 14) | NE | N/A nm, 1000 ms | mRNP nuclear export occurs faster than nucleoplasmic diffusion. Transcription, transport, and export occur between 5 and 40 min. | [62] |
| Super-registration microscopy | Mouse embryo fibroblasts | β-Actin (3.3) | NE | 25 nm, 20 ms | mRNA nuclear export includes a three-step process composed of docking, transport, and release with durations at ~80, 5–0, and ~80 ms respectively. mRNA movement is not limited to one direction within the NPC, and not all nuclear pores observed were equally active in mRNA export. | [63] |
| Light sheet microscopy | C. tentans salivary gland | C. tentans (Up to 40) | NE | 10 nm, 20 ms | ~25% of mRNPs successfully export through the NPC. Export time was determined to be between 65 ms to several seconds. With a dwell time of ~55 ms, Dbp5 interacts with the NPC most frequently from the cytoplasmic side. | [64] |
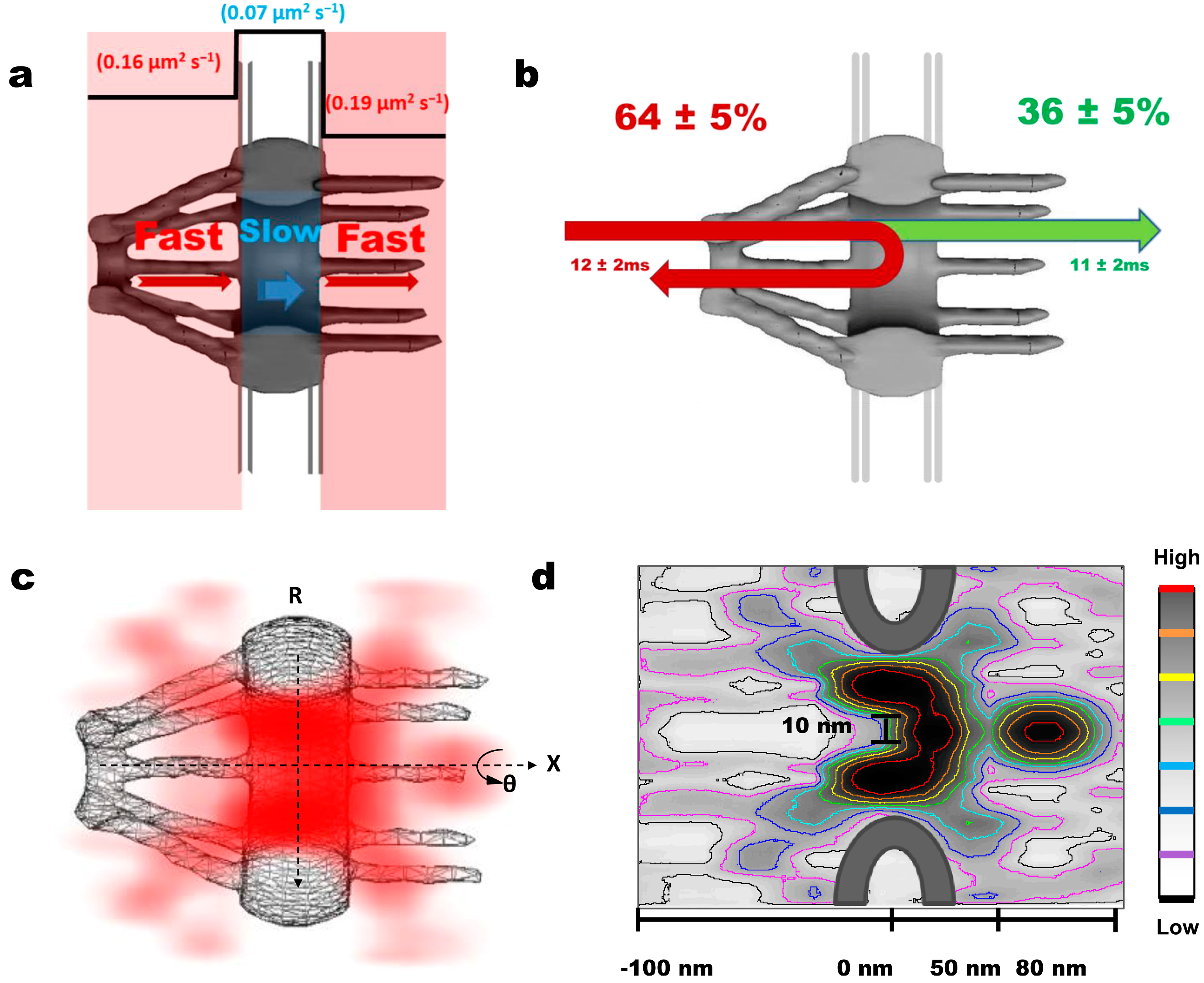
| SPEED microscopy | HeLa | Firefly Luciferase (3.3) | NPC | 8 nm, 2 ms | ~36% of mRNPs interacting with NPC successfully complete the export process. During their ~12 ms transport time, mRNPs adopt a fast-slow-fast diffusion pattern while interacting with the periphery of the NPC and rarely enter the central axial conduit reserved for passive diffusion revealed by a 3D reconstruction of the export route for mRNPs through the NPC in live cells. | [65] |
3. Various Single-Molecule Techniques Applied to Map Messenger Ribonucleoprotein Particles (mRNP) Export through the NPC

4. Conclusions and Perspective
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Köhler, A. Exporting RNA from the nucleus to the cytoplasm. Nat. Rev. Mol. Cell Biol. 2007, 8, 761–773. [Google Scholar] [CrossRef]
- Lamond, A.I.; Earnshaw, W.C. Structure and function in the nucleus. Science 1998, 280, 547–553. [Google Scholar] [CrossRef]
- Moore, M.J. From birth to death: The complex lives of eukaryotic mRNAs. Science 2005, 309, 1514–1518. [Google Scholar] [CrossRef]
- Gallouzi, I.E.; Steitz, J.A. Delineation of mRNA export pathways by the use of cell-permeable peptides. Science 2001, 294, 1895–1901. [Google Scholar] [CrossRef]
- Huang, Y.; Steitz, J.A. Splicing factors SRp20 and 9G8 promote the nucleocytoplasmic export of mRNA. Mol. Cell. 2001, 7, 899–905. [Google Scholar] [CrossRef]
- Huang, Y.; Carmichael, G.G. Role of polyadenylation in nucleocytoplasmic transport of mRNA. Mol. Cell. Biol. 1996, 16, 1534–1542. [Google Scholar]
- Minvielle-Sebastia, L.; Keller, W. mRNA polyadenylation and its coupling to other RNA processing reactions and to transcription. Curr. Opin. Cell Biol. 1999, 11, 352–357. [Google Scholar] [CrossRef]
- Eckner, R.; Ellmeier, W.; Birnstiel, M.L. Mature mRNA 3' end formation stimulates RNA export from the nucleus. EMBO J. 1991, 10, 3513–3522. [Google Scholar]
- Hammell, C.M.; Gross, S.; Zenklusen, D.; Heath, C.V.; Stutz, F.; Moore, C.; Cole, C.N. Coupling of termination, 3' processing, and mRNA export. Mol. Cell. Biol. 2002, 22, 6441–6457. [Google Scholar] [CrossRef]
- Visa, N.; Izaurralde, E.; Ferreira, J.; Daneholt, B.; Mattaj, I.W. A nuclear cap-binding complex binds Balbiani ring pre-mRNA cotranscriptionally and accompanies the ribonucleoprotein particle during nuclear export. J. Cell Biol. 1996, 133, 5–14. [Google Scholar] [CrossRef]
- Cheng, H.; Dufu, K.; Lee, C.S.; Hsu, J.L.; Dias, A.; Reed, R. Human mRNA export machinery recruited to the 5' end of mRNA. Cell 2006, 127, 1389–1400. [Google Scholar] [CrossRef]
- Zenklusen, D.; Vinciguerra, P.; Wyss, J.C.; Stutz, F. Stable mRNP formation and export require cotranscriptional recruitment of the mRNA export factors Yra1p and Sub2p by Hpr1p. Mol. Cell. Biol. 2002, 22, 8241–8253. [Google Scholar] [CrossRef]
- Dreyfuss, G. Structure and function of nuclear and cytoplasmic ribonucleoprotein particles. Annu. Rev. Cell. Biol. 1986, 2, 459–498. [Google Scholar] [CrossRef]
- Aguilera, A. Cotranscriptional mRNP assembly: From the DNA to the nuclear pore. Curr. Opin. Cell Biol. 2005, 17, 242–250. [Google Scholar] [CrossRef]
- Terry, L.J.; Shows, E.B.; Wente, S.R. Crossing the nuclear envelope: Hierarchical regulation of nucleocytoplasmic transport. Science 2007, 318, 1412–1416. [Google Scholar] [CrossRef]
- Kelly, S.M.; Corbett, A.H. Messenger RNA export from the nucleus: A series of molecular wardrobe changes. Traffic 2009, 10, 1199–1208. [Google Scholar] [CrossRef]
- Montpetit, B.; Weis, K. An alternative route for nuclear mRNP export by membrane budding. Science 2012, 336, 809–810. [Google Scholar] [CrossRef]
- Naim, B.; Brumfeld, V.; Kapon, R.; Kiss, V.; Nevo, R.; Reich, Z. Passive and facilitated transport in nuclear pore complexes is largely uncoupled. J. Biol. Chem. 2007, 282, 3881–3888. [Google Scholar]
- Weis, K. The nuclear pore complex: Oily spaghetti or gummy bear? Cell 2007, 130, 405–407. [Google Scholar] [CrossRef]
- Goryaynov, A.; Yang, W. Role of molecular charge in nucleocytoplasmic transport. PLoS One 2014, 9, e88792. [Google Scholar] [CrossRef]
- Grünwald, D.; Singer, R.H.; Rout, M. Nuclear export dynamics of RNA–protein complexes. Nature 2011, 475, 333–341. [Google Scholar] [CrossRef]
- Rout, M.P.; Aitchison, J.D.; Magnasco, M.O.; Chait, B.T. Virtual gating and nuclear transport: The hole picture. Trends Cell Biol. 2003, 13, 622–628. [Google Scholar] [CrossRef]
- Ma, J.; Goryaynov, A.; Sarma, A.; Yang, W. Self-regulated viscous channel in the nuclear pore complex. Proc. Natl. Acad. Sci. USA 2012, 109, 7326–7331. [Google Scholar] [CrossRef]
- Kahms, M.; Hüve, J.; Wesselmann, R.; Farr, J.C.; Baumgärtel, V.; Peters, R. Lighting up the nuclear pore complex. Eur. J. Cell Biol. 2011, 90, 751–758. [Google Scholar] [CrossRef]
- Fried, H.; Kutay, U. Nucleocytoplasmic transport: Taking an inventory. Cell. Mol. Life Sci. 2003, 60, 1659–1688. [Google Scholar] [CrossRef]
- Labokha, A.A.; Gradmann, S.; Frey, S.; Hülsmann, B.B.; Urlaub, H.; Baldus, M.; Görlich, D. Systematic analysis of barrier forming FG hydrogels from Xenopus nuclear pore complexes. EMBO J. 2013, 32, 204–218. [Google Scholar]
- Hülsmann, B.B.; Labokha, A.A.; Görlich, D. The permeability of reconstituted nuclear pores provides direct evidence for the selective phase model. Cell 2012, 150, 738–751. [Google Scholar] [CrossRef]
- Petri, M.; Frey, S.; Menzel, A.; Görlich, D. Techert, S Structural characterization of nanoscale meshworks within a nucleoporin FG hydrogel. Biomacromolecules 2012, 13, 1882–1889. [Google Scholar] [CrossRef]
- Milles, S.; Lemke, E.A. Single molecule study of the intrinsically disordered FG-repeat nucleoporin 153. Biophys. J. 2011, 101, 1710–1719. [Google Scholar] [CrossRef]
- Wälde, S.; Kehlenbach, R.H. The part and the whole: Functions of nucleoporins in nucleocytoplasmic transport. Trends Cell. Biol. 2010, 20, 461–469. [Google Scholar] [CrossRef]
- Stelter, P.; Kunze, R.; Fischer, J.; Hurt, E. Probing the nucleoporin FG repeat network defines structural and functional features of the nuclear pore complex. J. Cell Sci. 2011, 195, 183–192. [Google Scholar]
- Yang, W. “Natively unfolded” nucleoporins in nucleocytoplasmic transport: Clustered or evenly distributed. Nucleus 2010, 2, 10–16. [Google Scholar] [CrossRef]
- Patel, S.; Belmont, B.; Sante, J.; Rexach, M. Natively unfolded nucleoporins gate protein diffusion across the nuclear pore complex. Cell 2007, 129, 83–96. [Google Scholar] [CrossRef]
- Reed, R.; Hurt, E. A conserved mRNA export machinery coupled to pre-mRNA splicing. Cell 2002, 108, 523–531. [Google Scholar] [CrossRef]
- Wickramasinghe, V.O.; Andrews, R.; Ellis, P.; Langford, C.; Gurdon, J.B.; Stewart, M.; Venkitaraman, A.R.; Laskey, R.A. Selective nuclear export of specific classes of mRNA from mammalian nuclei is promoted by GANP. Nucleic Acids Res. 2014, 42, 5059–5071. [Google Scholar] [CrossRef]
- Sträßer, K.; Masuda, S.; Mason, P.; Pfannstiel, J.; Oppizzi, M.; Rodriguez-Navarro, S.; Rondón, A.G.; Aguilera, A.; Struhl, K.; Reed, R. TREX is a conserved complex coupling transcription with messenger RNA export. Nature 2002, 417, 304–308. [Google Scholar] [CrossRef]
- Lei, H.; Dias, A.P.; Reed, R. Export and stability of naturally intronless mRNAs require specific coding region sequences and the TREX mRNA export complex. Proc. Natl. Acad. Sci. USA 2011, 108, 17985–17990. [Google Scholar] [CrossRef]
- Jani, D.; Lutz, S.; Hurt, E.; Laskey, R.A.; Stewart, M.; Wickramasinghe, V.O. Functional and structural characterization of the mammalian TREX-2 complex that links transcription with nuclear messenger RNA export. Nucleic Acids Res. 2012, 40, 4562–4573. [Google Scholar] [CrossRef]
- Wang, L.; Miao, Y.L.; Zheng, X.; Lackford, B.; Zhou, B.; Han, L.; Chengguo, Y.; Ward, J.M.; Burkholder, A.; Lipchina, I.; et al. The THO complex regulates pluripotency gene mRNA export and controls embryonic stem cell self-renewal and somatic cell reprogramming. Cell Stem Cell 2013, 13, 676–690. [Google Scholar] [CrossRef]
- Guria, A.; Tran, D.D.H.; Ramachandran, S.; Koch, A.; el Bounkari, O.; Dutta, P.; Hauser, H.; Tamura, T. Identification of mRNAs that are spliced but not exported to the cytoplasm in the absence of THOC5 in mouse embryo fibroblasts. RNA 2011, 17, 1048–1056. [Google Scholar] [CrossRef]
- Wickramasinghe, V.O.; Savill, J.M.; Chavali, S.; Jonsdottir, A.B.; Rajendra, E.; Grüner, T.; Laskey, R.A.; Babu, M.M.; Venkitaraman, A.R. Human inositol polyphosphate multikinase regulates transcript-selective nuclear mRNA export to preserve genome integrity. Mol. Cell. 2013, 51, 737–750. [Google Scholar] [CrossRef]
- Grossman, E.; Medalia, O.; Zwerger, M. Functional architecture of the nuclear pore complex. Annu. Rev. Biophys. 2012, 41, 557–584. [Google Scholar] [CrossRef]
- Rout, M.P.; Blobel, G. Isolation of the yeast nuclear pore complex. J. Cell Biol. 1993, 123, 771–783. [Google Scholar] [CrossRef]
- Hoelz, A.; Debler, E.W.; Blobel, G. The structure of the nuclear pore complex. Annu. Rev. Biochem. 2011, 80, 613–643. [Google Scholar] [CrossRef]
- Yang, W. Distinct, but not completely separate spatial transport routes in the nuclear pore complex. Nucleus 2013, 4, 166–175. [Google Scholar] [CrossRef]
- Tamura, K.; Hara-Nishimura, I. The molecular architecture of the plant nuclear pore complex. J. Exp. Bot. 2013, 64, 823–832. [Google Scholar] [CrossRef]
- Diebolder, C.; Koster, A.J.; Koning, R.I. Pushing the resolution limits in cryo electron tomography of biological structures. J. Microsc. 2012, 248, 1–5. [Google Scholar] [CrossRef]
- Penczek, P. Chapter three—Resolution measures in electron microscopy. Methods Enzymol. 2010, 482, 73–100. [Google Scholar] [CrossRef]
- Beermann, W.; Bahr, G.F. The submicroscopic structure of the Balbiani-ring. Exp. Cell Res. 1954, 6, 195–201. [Google Scholar] [CrossRef]
- Mehlin, H.; Daneholt, B.; Skoglund, U. Translocation of a specific premessenger ribonucleoprotein particle through the nuclear pore studied with electron microscope tomography. Cell 1992, 69, 605–613. [Google Scholar] [CrossRef]
- Lamb, M.M.; Daneholt, B. Characterization of active transcription units in Balbiani rings of Chironomus tentans. Cell 1979, 17, 835–848. [Google Scholar] [CrossRef]
- Stevens, B.J.; Swift, H. RNA transport from nucleus to cytoplasm in Chironomus salivary glands. J. Cell. Biol. 1966, 31, 55–77. [Google Scholar] [CrossRef]
- Mehlin, H.; Skoglund, U.; Daneholt, B. Transport of Balbiani ring granules through nuclear pores in Chironomus tentans. Exp. Cell Res. 1991, 193, 72–77. [Google Scholar] [CrossRef]
- Cole, C.N.; Scarcelli, J.J. Unravelling mRNA export. Nat. Cell Biol. 2006, 8, 645–647. [Google Scholar] [CrossRef]
- Patterson, G.H.; Lippincott-Schwartz, J. A photoactivatable GFP for selective photolabeling of proteins and cells. Science 2002, 297, 1873–1877. [Google Scholar] [CrossRef]
- Huang, B.; Bates, M.; Zhuang, X. Super resolution fluorescence microscopy. Annu. Rev. Biochem. 2009, 78, 993–1016. [Google Scholar] [CrossRef]
- Huang, B.; Wang, W.; Bates, M.; Zhuang, X. Three-dimensional super-resolution imaging by stochastic optical reconstruction microscopy. Science 2008, 319, 810–813. [Google Scholar] [CrossRef]
- Juette, M.F.; Gould, T.J.; Lessard, M.D.; Mlodzianoski, M.J.; Nagpure, B.S.; Bennett, B.T.; Hess, S.T.; Bewersdorf, J. Three-dimensional sub-100 nm resolution fluorescence microscopy of thick samples. Nat. Methods 2008, 5, 527–529. [Google Scholar] [CrossRef]
- Bates, M.; Huang, B.; Zhuang, X. Super-resolution microscopy by nanoscale localization of photo-switchable fluorescent probes. Curr. Opin. Chem. Biol. 2008, 12, 505–514. [Google Scholar] [CrossRef]
- Pavani, S.R.P.; Thompson, M.A.; Biteen, J.S.; Lord, S.J.; Liu, N.; Twieg, R.J.; Piestun, R.; Moerner, W.E. Three-dimensional, single-molecule fluorescence imaging beyond the diffraction limit by using a double-helix point spread function. Proc. Natl. Acad. Sci. USA 2009, 106, 2995–2999. [Google Scholar] [CrossRef]
- Löschberger, A.; van de Linde, S.; Dabauvalle, M.C.; Rieger, B.; Heilemann, M.; Krohne, G.; Sauer, M. Super-resolution imaging visualizes the eightfold symmetry of gp210 proteins around the nuclear pore complex and resolves the central channel with nanometer resolution. J. Cell Sci. 2012, 125, 570–575. [Google Scholar] [CrossRef]
- Mor, A.; Suliman, S.; Ben-Yishay, R.; Yunger, S.; Brody, Y.; Shav-Tal, Y. Dynamics of single mRNP nucleocytoplasmic transport and export through the nuclear pore in living cells. Nat. Cell Biol. 2010, 12, 543–552. [Google Scholar] [CrossRef]
- Grünwald, D.; Singer, R.H. In vivo imaging of labelled endogenous β-actin mRNA during nucleocytoplasmic transport. Nature 2010, 467, 604–607. [Google Scholar] [CrossRef]
- Siebrasse, J.P.; Kaminski, T.; Kubitscheck, U. Nuclear export of single native mRNA molecules observed by light sheet fluorescence microscopy. Proc. Natl. Acad. Sci. USA 2012, 109, 9426–9431. [Google Scholar] [CrossRef]
- Ma, J.; Liu, Z.; Michelotti, N.; Pitchiaya, S.; Veerapaneni, R.; Androsavich, J.R.; Walter, N.G.; Yang, W. High-resolution three-dimensional mapping of mRNA export through the nuclear pore. Nat. Commun. 2013, 4, 2414. [Google Scholar]
- Stewart, M. Ratcheting mRNA out of the nucleus. Mol. Cell 2007, 25, 327–330. [Google Scholar] [CrossRef]
- Huisken, J.; Stainier, D.Y. Selective plane illumination microscopy techniques in developmental biology. Development 2009, 136, 1963–1975. [Google Scholar] [CrossRef]
- Keller, P.J.; Dodt, H.U. Light sheet microscopy of living or cleared specimens. Curr. Opin. Neurobiol. 2012, 22, 138–143. [Google Scholar] [CrossRef]
- Ritter, J.G.; Veith, R.; Veenendaal, A.; Siebrasse, J.P.; Kubitscheck, U. Light sheet microscopy for single molecule tracking in living tissue. PLoS One 2010, 5, e11639. [Google Scholar]
- Santi, P.A. Light sheet fluorescence microscopy: A review. J. Histochem. Cytochem. 2011, 59, 129–138. [Google Scholar] [CrossRef]
- Ribbeck, K.; Lipowsky, G.; Kent, H.M.; Stewart, M.; Görlich, D. NTF2 mediates nuclear import of Ran. EMBO J. 1998, 17, 6587–6598. [Google Scholar] [CrossRef]
- Paschal, B.M.; Gerace, L. Identification of NTF2, a cytosolic factor for nuclear import that interacts with nuclear pore complex protein p62. J. Cell. Biol. 1995, 129, 925–937. [Google Scholar] [CrossRef]
- Smith, A.; Brownawell, A.; Macara, I.G. Nuclear import of Ran is mediated by the transport factor NTF2. Curr. Biol. 1998, 8, 1403–1406. [Google Scholar] [CrossRef]
- Tran, E.J.; Zhou, Y.; Corbett, A.H.; Wente, S.R. The DEAD-box protein Dbp5 controls mRNA export by triggering specific RNA: Protein remodeling events. Mol. Cell 2007, 28, 850–859. [Google Scholar] [CrossRef]
- Zhao, J.; Jin, S.B.; Björkroth, B.; Wieslander, L.; Daneholt, B. The mRNA export factor Dbp5 is associated with Balbiani ring mRNP from gene to cytoplasm. EMBO J. 2002, 21, 1177–1187. [Google Scholar] [CrossRef]
- Goryaynov, A.; Ma, J.; Yang, W. Single-molecule studies of nucleocytoplasmic transport: From one dimension to three dimensions. Integr. Biol. 2012, 4, 10–21. [Google Scholar] [CrossRef]
- Ma, J.; Yang, W. Three-dimensional distribution of transient interactions in the nuclear pore obtained by single-molecule snapshots. Proc. Natl. Acad. Sci. USA 2010, 107, 7305–7310. [Google Scholar] [CrossRef]
- Paulillo, S.M.; Phillips, E.M.; Köser, J.; Sauder, U.; Ullman, K.S.; Powers, M.A.; Fahrenkrog, B. Nucleoporin domain topology is linked to the transport status of the nuclear pore complex. J. Mol. Biol. 2005, 351, 784–798. [Google Scholar] [CrossRef]
- Walther, T.C.; Pickersgill, H.S.; Cordes, V.C.; Goldberg, M.W.; Allen, T.D.; Mattaj, I.W.; Fornerod, M. The cytoplasmic filaments of the nuclear pore complex are dispensable for selective nuclear protein import. J. Cell Boil. 2002, 158, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Mahadevan, K.; Zhang, H.; Akef, A.; Cui, X.A.; Gueroussov, S.; Cenik, C.; Roth, F.P.; Palazzo, A.F. RanBP2/Nup358 potentiates the translation of a subset of mRNAs encoding secretory proteins. PLoS Biol. 2013, 11, e1001545. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kelich, J.M.; Yang, W. High-Resolution Imaging Reveals New Features of Nuclear Export of mRNA through the Nuclear Pore Complexes. Int. J. Mol. Sci. 2014, 15, 14492-14504. https://doi.org/10.3390/ijms150814492
Kelich JM, Yang W. High-Resolution Imaging Reveals New Features of Nuclear Export of mRNA through the Nuclear Pore Complexes. International Journal of Molecular Sciences. 2014; 15(8):14492-14504. https://doi.org/10.3390/ijms150814492
Chicago/Turabian StyleKelich, Joseph M., and Weidong Yang. 2014. "High-Resolution Imaging Reveals New Features of Nuclear Export of mRNA through the Nuclear Pore Complexes" International Journal of Molecular Sciences 15, no. 8: 14492-14504. https://doi.org/10.3390/ijms150814492
APA StyleKelich, J. M., & Yang, W. (2014). High-Resolution Imaging Reveals New Features of Nuclear Export of mRNA through the Nuclear Pore Complexes. International Journal of Molecular Sciences, 15(8), 14492-14504. https://doi.org/10.3390/ijms150814492