Inhibitory Effects of Adlay Extract on Melanin Production and Cellular Oxygen Stress in B16F10 Melanoma Cells


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
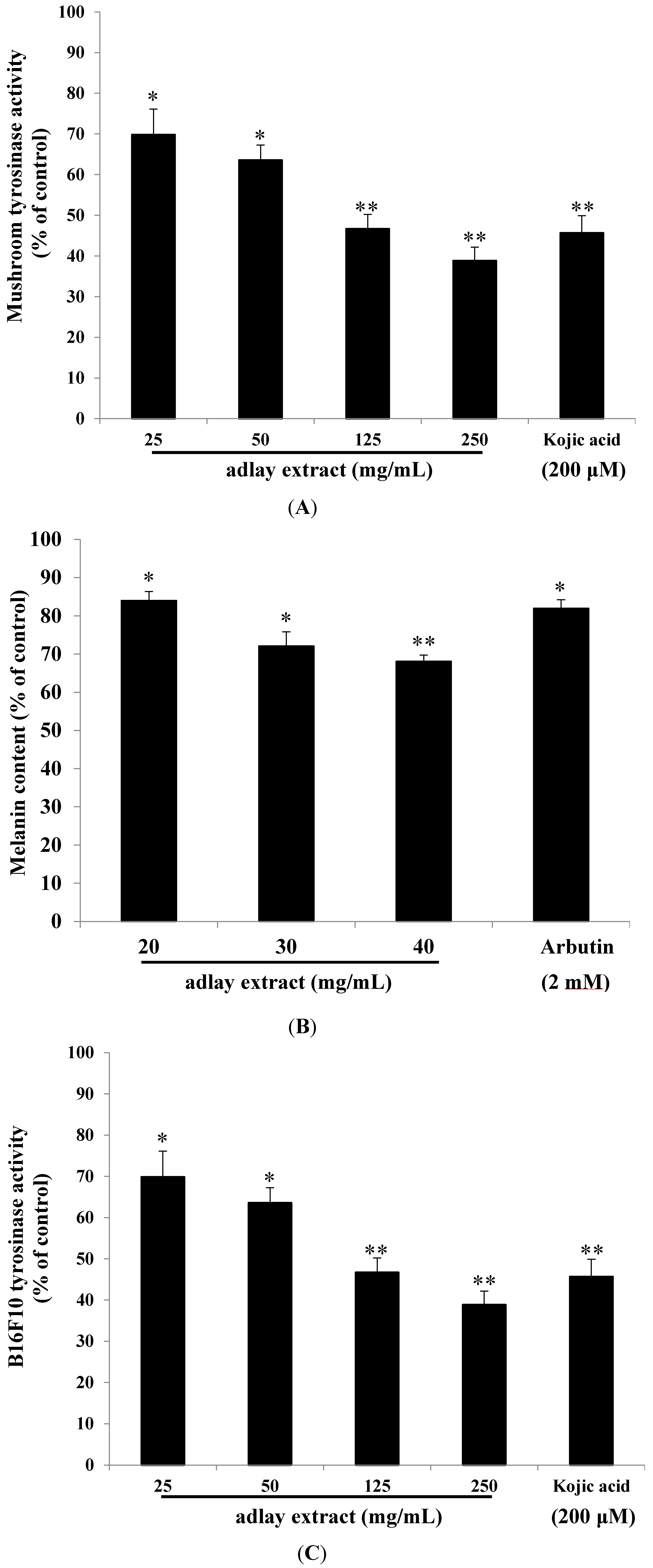
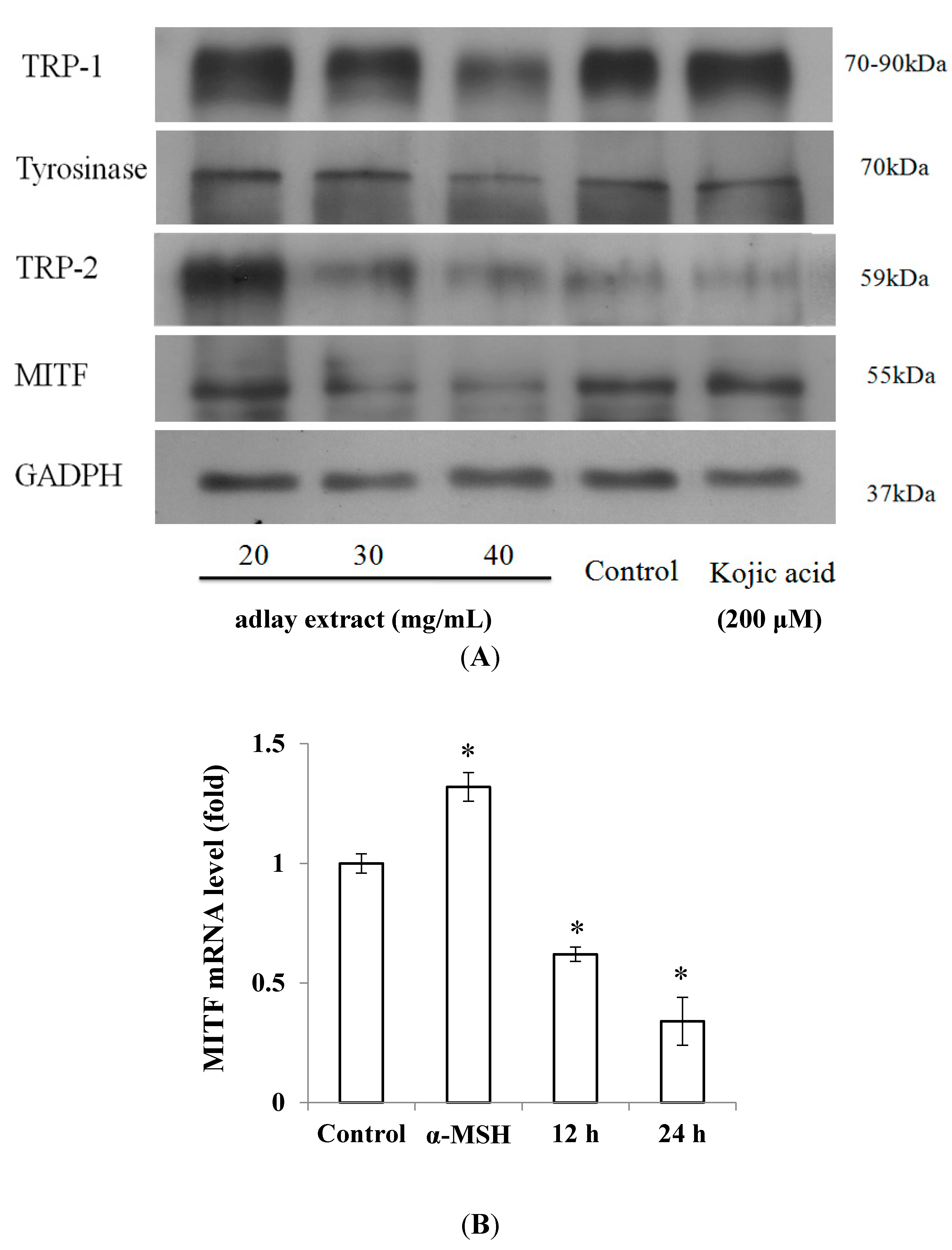
2.1. Effects of Adlay Extract on Mushroom Tyrosinase Activity and Melanogenesis


2.2. Antioxidant Characteristics of the Adlay Extract

3. Experimental Section
3.1. Chemicals and Reagents
3.2. Supercritical Fluid CO2 Extraction (SFE) of Adlay Seeds
3.3. Inhibitory Effects of Adlay Extract on Melanogenesis
3.4. RT-PCR
3.5. Antioxidant Characteristics of Adlay Extract
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Garcia, A.; Fulton, J.E. The combination of glycolic acid and hydroquinone or kojic acid for the treatment of melasma and related conditions. Dermatol. Surg. 1996, 22, 443–447. [Google Scholar] [PubMed]
- Virador, V.M.; Kobayashi, N.; Matsunaga, J.; Hearing, V.J. A standardized protocol for assessing regulators of pigmentation. Anal. Biochem. 1999, 270, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Rendon, M.I.; Gaviria, J.I. Review of Skin-Lightening Agents. Dermatol. Surg. 2005, 31, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J.; Jiménez, M. Mammalian tyrosinase—The critical regulatory control point in melanocyte pigmentation. Int. J. Biochem. 1987, 19, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [PubMed]
- Meyskens, F.L., Jr.; Chau, H.V.; Tohidian, N.; Buckmeier, J. Luminol-enhanced chemiluminescent response of human melanocytes and melanoma cells to hydrogen peroxide stress. Pigment Cell Res. 1997, 10, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Funasaka, Y.; Komoto, M.; Ichihashi, M. Depigmenting effect of alpha-tocopheryl ferulate on normal human melanocytes. Pigment Cell Res. 2000, 13 (Suppl. 8), 170–174. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.K.; Funasaka, Y.; Slominski, A.; Ermak, G.; Hwang, J.; Pawelek, J.M.; Ichihashi, M. Production and release of proopiomelanocortin (POMC) derived peptides by human melanocytes and keratinocytes in culture: Regulation by ultraviolet B. BBA Mol. Cell Res. 1996, 1313, 130–138. [Google Scholar]
- Sasaki, M.; Kizawa, K.; Igarashi, S.; Horikoshi, T.; Uchiwa, H.; Miyachi, Y. Suppression of melanogenesis by induction of endogenous intracellular metallothionein in human melanocytes. Exp. Dermatol. 2004, 13, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Yamakoshi, J.; Otsuka, F.; Sano, A.; Tokutake, S.; Saito, M.; Kikuchi, M.; Kubota, Y. Lightening effect on ultraviolet-induced pigmentation of guinea pig skin by oral administration of a proanthocyanidin-rich extract from grape seeds. Pigment Cell Res. 2003, 16, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Stahl, W. Nutritional protection against skin damage from sunlight. Annu. Rev. Nutr. 2004, 24, 173–200. [Google Scholar] [CrossRef] [PubMed]
- Overwijk, W.W.; Restifo, N.P. B16 as a mouse model for human melanoma. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]
- Otsuka, H.; Hirai, Y.; Nagao, T.; Yamasaki, K. Anti-inflammatory activity of benzoxazinoids from roots of Coix lachryma-jobi var. ma-yuen. J. Nat. Prod. 1988, 51, 74–79. [Google Scholar]
- Chen, H.-J.; Shih, C.-K.; Hsu, H.-Y.; Chiang, W. Mast cell-dependent allergic responses are inhibited by ethanolic extract of adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) testa. J. Agric. Food Chem. 2010, 58, 2596–2601. [Google Scholar]
- Seo, W.-G.; Pae, H.-O.; Chai, K.-Y.; Yun, Y.-G.; Kwon, T.-H.; Chung, H.-T. Inhibitory effects of methanol extract of seeds of job’s tears (Coix Lachryma-Jobi L Var. Ma-Yuen) on nitric oxide and superoxide production in raw 264.7 macrophages. Immunopharm. Immunotoxicol. 2000, 22, 545–554. [Google Scholar]
- Kuo, C.-C.; Shih, M.-C.; Kuo, Y.-H.; Chiang, W. Antagonism of free-radical-induced damage of adlay seed and its antiproliferative effect in human histolytic lymphoma U937 monocytic cells. J. Agric. Food Chem. 2001, 49, 1564–1570. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-L.; Liu, R.H.; Sheu, J.-N.; Chen, S.-T.; Sinchaikul, S.; Tsay, G.J. Toxicogenomics of A375 human malignant melanoma cells treated with arbutin. J. Biomed. Sci. 2007, 14, 87–105. [Google Scholar] [CrossRef] [PubMed]
- García-Gavín, J.; González-Vilas, D.; Fernández-Redondo, V.; Toribio, J. Pigmented contact dermatitis due to kojic acid. A paradoxical side effect of a skin lightener. Contact Dermat. 2010, 62, 63–64. [Google Scholar]
- Emerit, I. Free radicals and aging of the skin. EXS 1992, 62, 328–341. [Google Scholar] [PubMed]
- Sapkota, K.; Park, S.-E.; Kim, J.-E.; Kim, S.; Choi, H.-S.; Chun, H.-S.; Kim, S.-J. Antioxidant and antimelanogenic properties of chestnut flower extract. Biosci. Biotechnol. Biochem. 2010, 74, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.-Y.; Chou, T.-H.; Lin, R.-J.; Chan, L.-P.; Wang, G.-H.; Liang, C.-H. Antioxidant and antimelanogenic behaviors of Paeonia suffruticosa. Plant Food Hum. Nutr. 2011, 66, 275–284. [Google Scholar] [CrossRef]
- Bilodeau, M.L.; Greulich, J.D.; Hullinger, R.L.; Bertolotto, C.; Ballotti, R.; Andrisani, O.M. BMP-2 stimulates tyrosinase gene expression and melanogenesis in differentiated melanocytes. Pigment Cell Res. 2001, 14, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, T.; Kondoh, H.; Hiratsuka, J.; Mishima, Y. Enhanced melanogenesis induced by tyrosinase gene-transfer increases boron-uptake and killing effect of boron neutron capture therapy for amelanotic melanoma. Pigment Cell Res. 1998, 11, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Koo, J.H.; Song, Y.G.; Kwon, K.B.; Lee, J.H.; Sohn, H.S.; Park, B.H.; Jhee, E.C.; Park, J.W. Stimulation of melanogenesis by scoparone in B16 melanoma cells. Acta Pharmacol. Sin. 2006, 27, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Chang, T.-M.; Chang, Y.-J.; Wen, H.-Y. UVB Irradiation Regulates ERK1/2- and p53-Dependent Thrombomodulin Expression in Human Keratinocytes. PLoS One 2013, 8, e67632. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on products of browing reaction: Antioxidative activity of product of browing reaction preapared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Galato, D.; Ckless, K.; Susin, M.F.; Giacomelli, C.; Ribeiro-do-Valle, R.M.; Spinelli, A. Antioxidant capacity of phenolic and related compounds: Correlation among electrochemical, visible spectroscopy methods and structure–antioxidant activity. Redox Rep. 2001, 6, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Murrant, C.L.; Reid, M.B. Detection of reactive oxygen and reactive nitrogen species in skeletal muscle. Microsc. Res. Tech. 2001, 55, 236–248. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, H.-C.; Hsieh, W.-Y.; Niu, Y.-L.; Chang, T.-M. Inhibitory Effects of Adlay Extract on Melanin Production and Cellular Oxygen Stress in B16F10 Melanoma Cells. Int. J. Mol. Sci. 2014, 15, 16665-16679. https://doi.org/10.3390/ijms150916665
Huang H-C, Hsieh W-Y, Niu Y-L, Chang T-M. Inhibitory Effects of Adlay Extract on Melanin Production and Cellular Oxygen Stress in B16F10 Melanoma Cells. International Journal of Molecular Sciences. 2014; 15(9):16665-16679. https://doi.org/10.3390/ijms150916665
Chicago/Turabian StyleHuang, Huey-Chun, Wan-Yu Hsieh, Yu-Lin Niu, and Tsong-Min Chang. 2014. "Inhibitory Effects of Adlay Extract on Melanin Production and Cellular Oxygen Stress in B16F10 Melanoma Cells" International Journal of Molecular Sciences 15, no. 9: 16665-16679. https://doi.org/10.3390/ijms150916665
APA StyleHuang, H. -C., Hsieh, W. -Y., Niu, Y. -L., & Chang, T. -M. (2014). Inhibitory Effects of Adlay Extract on Melanin Production and Cellular Oxygen Stress in B16F10 Melanoma Cells. International Journal of Molecular Sciences, 15(9), 16665-16679. https://doi.org/10.3390/ijms150916665