Mechanistic Studies of Anti-Hyperpigmentary Compounds: Elucidating Their Inhibitory and Regulatory Actions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results
2.1. Effects of the Chemical Compounds on Signaling Protein Expression Levels in Melan-a Cells

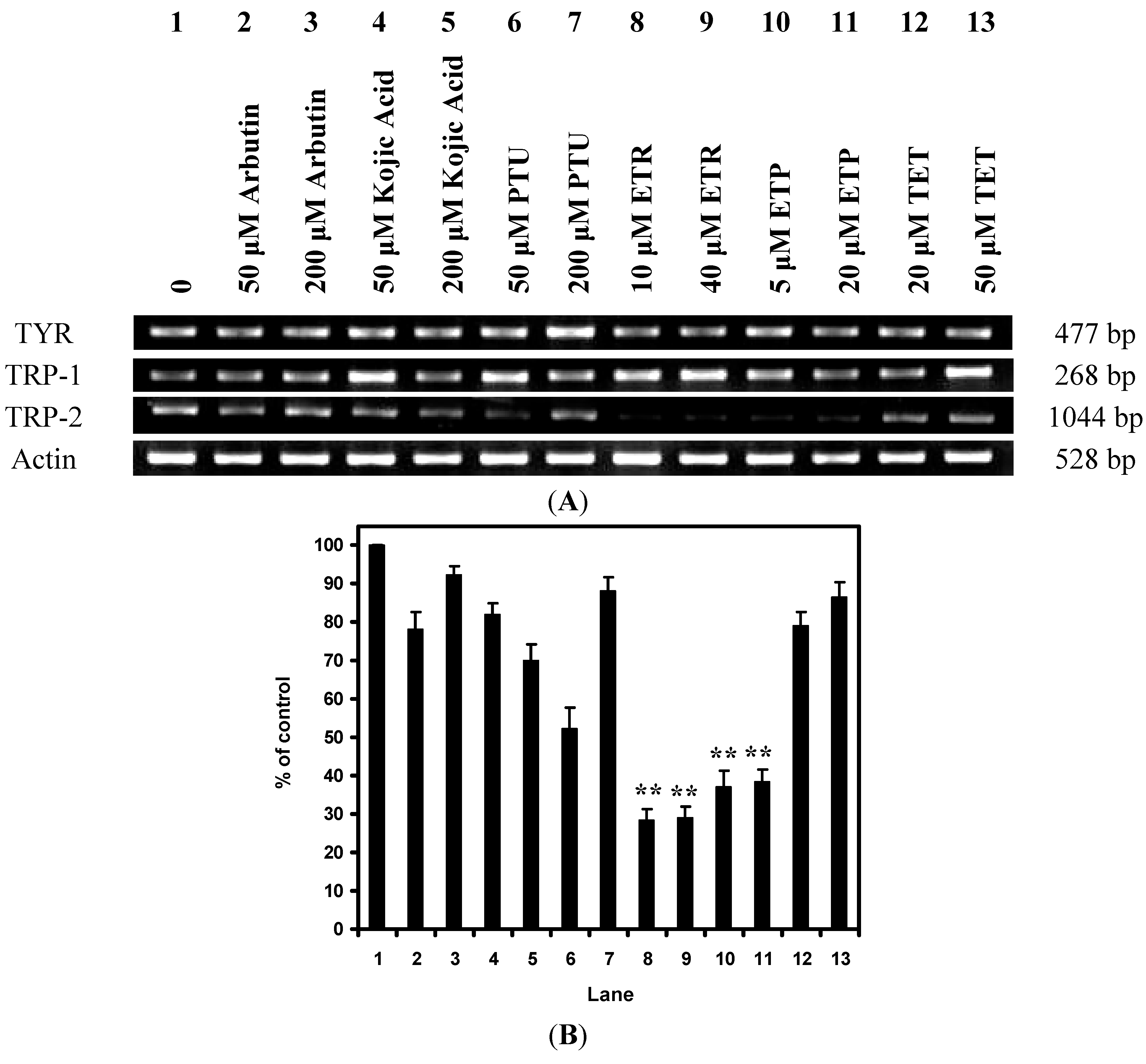
2.2. Effects of the Chemical Compounds on Gene Expression Level in Melan-a Cells

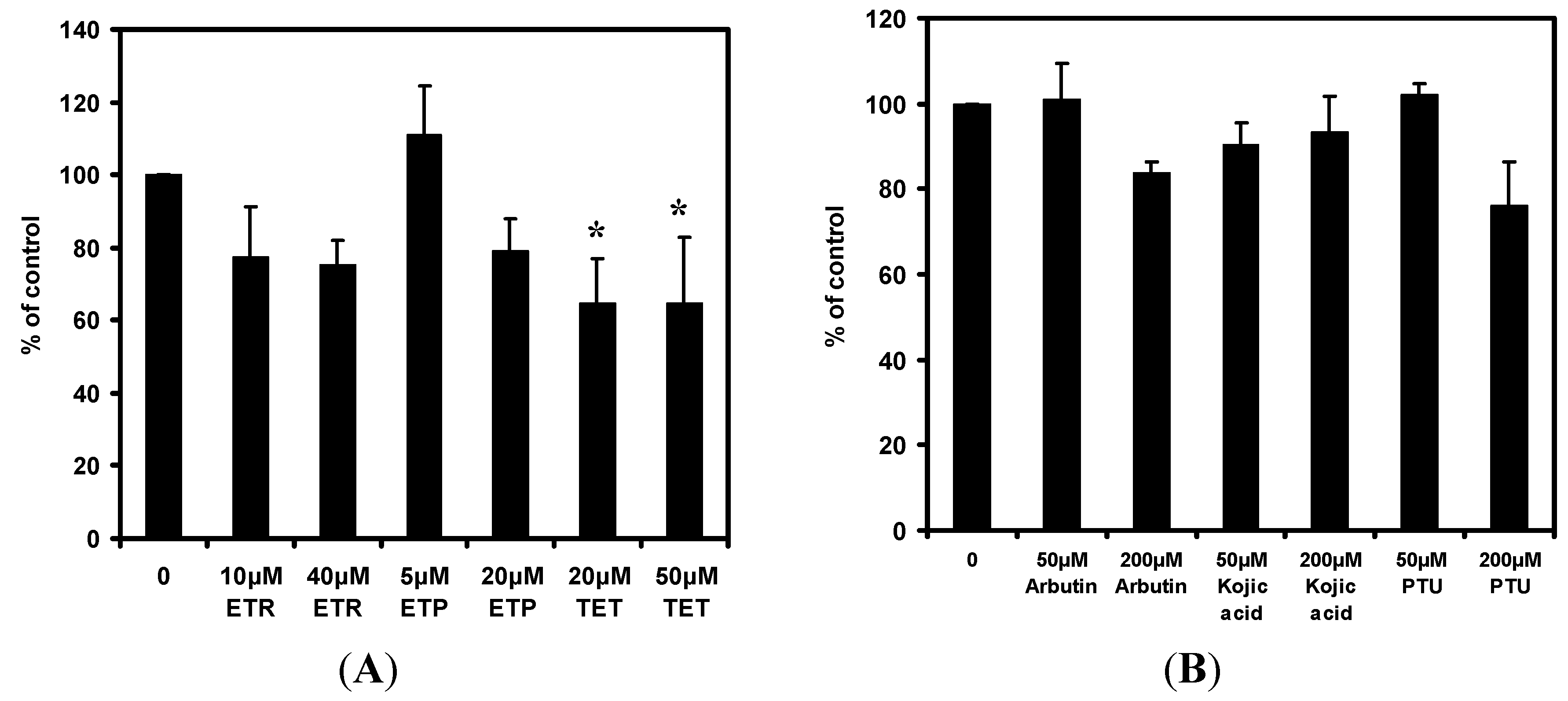
2.3. Effects of the Chemical Compounds on Cyclic Adenosine Monophosphate (cAMP) Level in Melan-a Cells

2.4. Effects of the Chemical Compounds on Protein Kinase A (PKA) Activity in Melan-a Cells

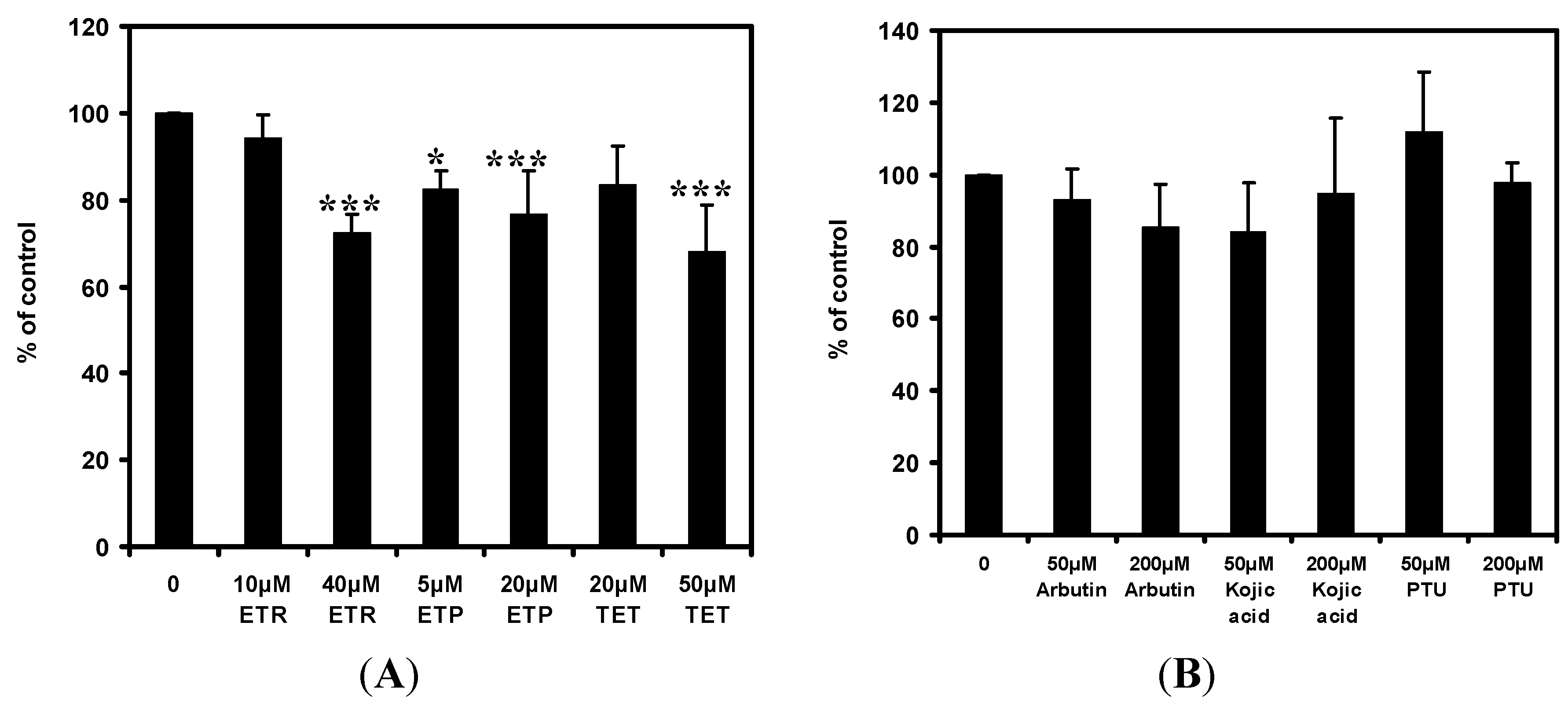
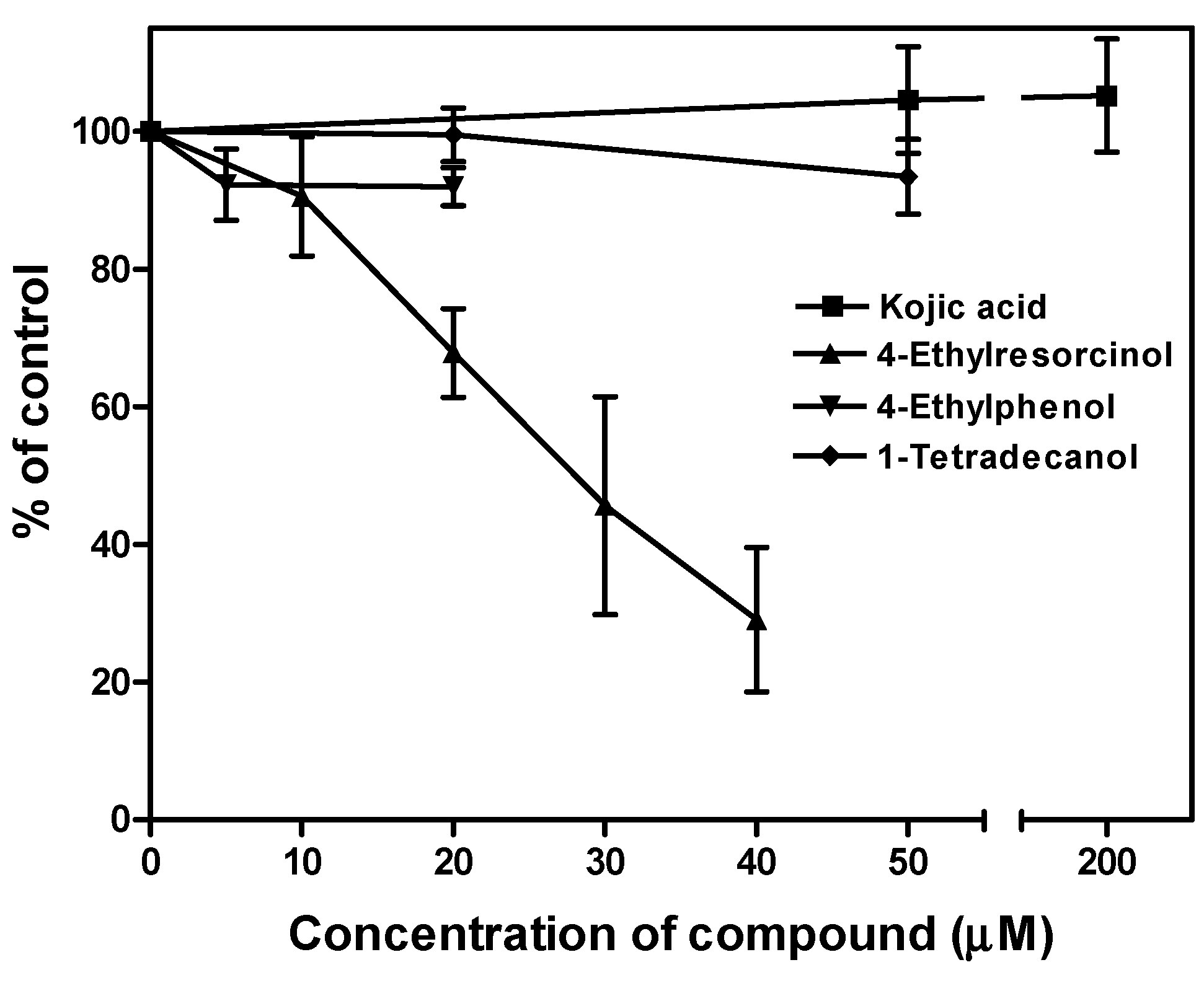
2.5. Effects of the Chemical Compounds on Tyrosinase Enzyme Activity

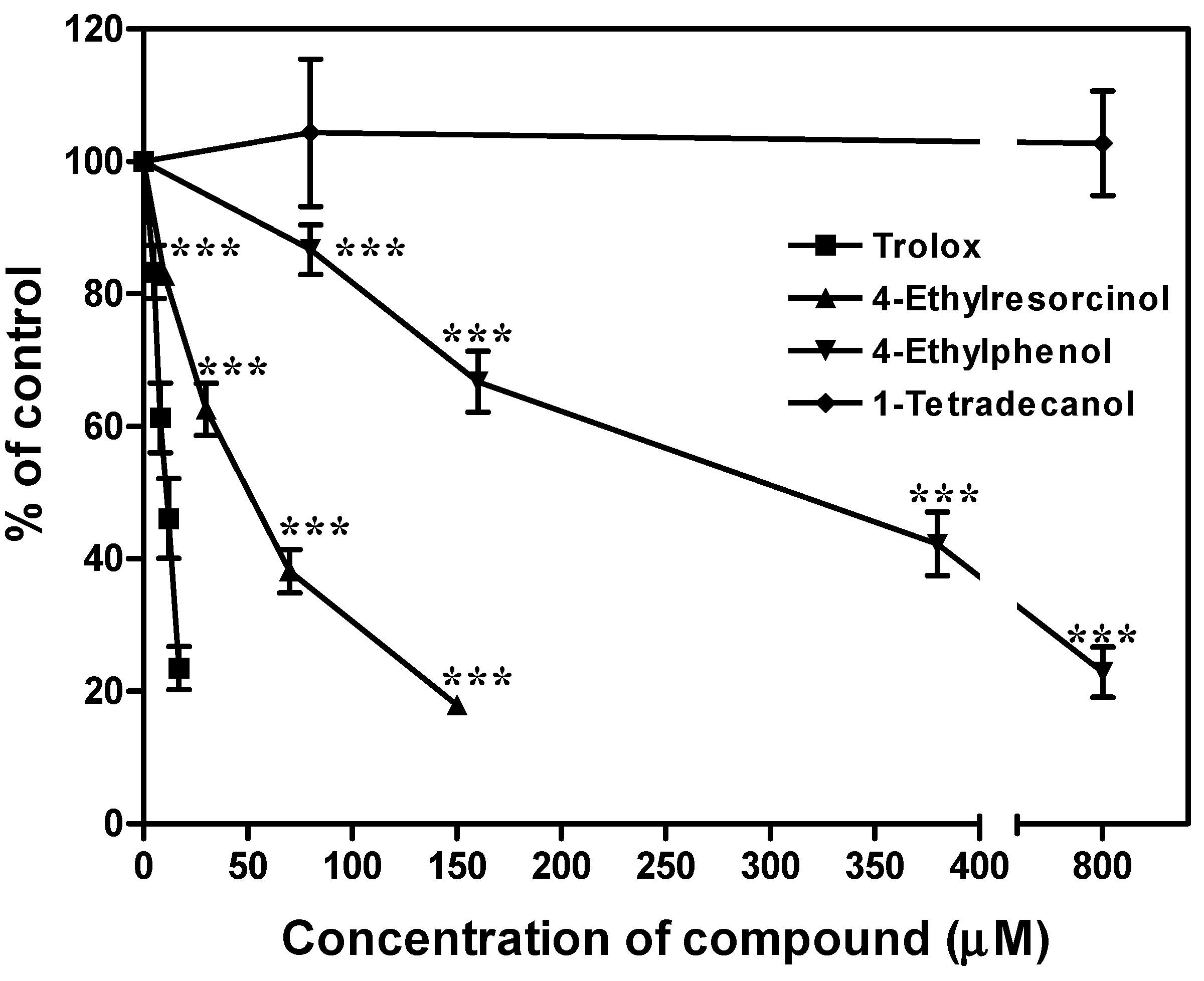
2.6. Effects of the Chemical Compounds on Lipid Peroxidation of Mouse Liver Microsome

3. Discussion
4. Experimental Section
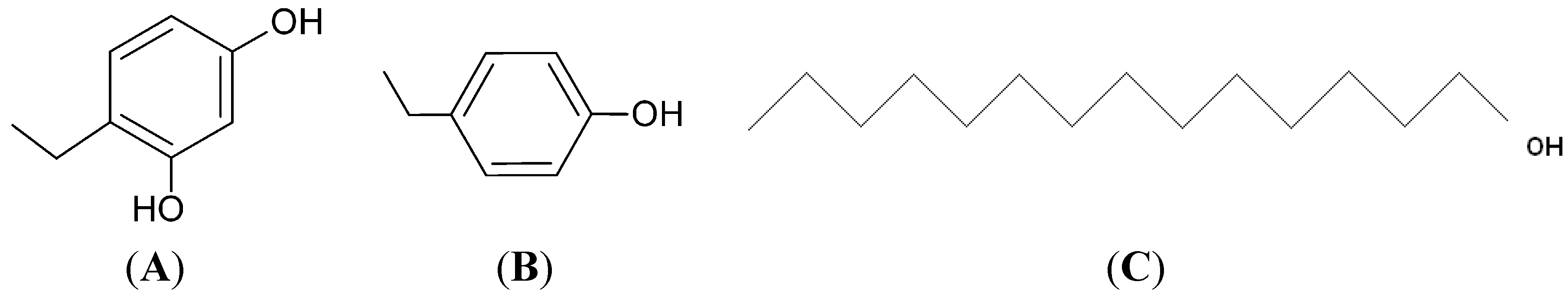
4.1. Materials
4.2. Cell Culture
4.3. Western Blot Analysis
4.3.1. Preparation of Cell Lysates
4.3.2. Protein Assay
4.3.3. Western Blot
4.4. RT-PCR
4.5. cAMP Assay
4.6. The PKA Activity Assay
4.7. Tyrosinase Inhibition Assay
4.8. Antioxidative Activity Assay
4.8.1. Preparation of Mouse Liver Microsomes
4.8.2. Lipid Peroxidation Inhibition Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Im, S.; Moro, O.; Peng, F.; Medrano, E.E.; Cornelius, J.; Babcock, G.; Nordlund, J.J.; Abdel-Malek, Z.A. Activation of the cyclic AMP pathway by α-melanotropin mediates the response of human melanocytes to ultraviolet B radiation. Cancer Res. 1998, 58, 47–54. [Google Scholar] [PubMed]
- Bertolotto, C.; Abbe, P.; Hemesath, T.J.; Bille, K.; Fisher, D.E.; Ortonne, J.P.; Ballotti, R. Microphthalmia gene product as a signal transducer in cAMP-induced differentiation of melanocytes. J. Cell Biol. 1998, 142, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, K.; Yokoyama, K.; Takahashi, K.; Tomita, Y.; Shibahara, S. Functional analysis of microphthalmia-associated transcription factor in pigment cell-specific transcription of the human tyrosinase family genes. J. Biol. Chem. 1997, 272, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar] [PubMed]
- Sanchez-Ferrer, A.; Rodriguez-Lopez, J.N.; Garcia-Canovas, F.; Garcia-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta 1995, 1247, 1–11. [Google Scholar]
- Korner, A.; Pawelek, J. Mammalian tyrosinase catalyzes three reactions in the biosynthesis of melanin. Science 1982, 217, 1163–1165. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Cervantes, C.; Solano, F.; Kobayashi, T.; Urabe, K.; Hearing, V.J.; Lozano, J.A.; Garcia-Borron, J.C. A new enzymatic function in the melanogenic pathway. The 5,6-dihydroxyindole-2-carboxylic acid oxidase activity of tyrosinase-related protein-1 (TRP1). J. Biol. Chem. 1994, 269, 17993–8000. [Google Scholar]
- Kobayashi, T.; Urabe, K.; Winder, A.; Jimenez-Cervantes, C.; Imokawa, G.; Brewington, T.; Solano, F.; Garcia-Borron, J.C.; Hearing, V.J. Tyrosinase related protein 1 (TRP1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J. 1994, 13, 5818–5825. [Google Scholar] [PubMed]
- Kobayashi, T.; Urabe, K.; Winder, A.; Tsukamoto, K.; Brewington, T.; Imokawa, G.; Potterf, B.; Hearing, V.J. DHICA oxidase activity of TRP1 and interactions with other melanogenic enzymes. Pigment. Cell Res. 1994, 7, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [PubMed]
- Yokoyama, K.; Suzuki, H.; Yasumoto, K.; Tomita, Y.; Shibahara, S. Molecular cloning and functional analysis of a cDNA coding for human DOPAchrome tautomerase/tyrosinase-related protein-2. Biochim. Biophys. Acta 1994, 1217, 317–321. [Google Scholar] [CrossRef]
- Saha, B.; Singh, S.K.; Sarkar, C.; Bera, R.; Ratha, J.; Tobin, D.J.; Bhadra, R. Activation of the Mitf promoter by lipid-stimulated activation of p38-stress signalling to CREB. Pigment. Cell Res. 2006, 19, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Sarkar, C.; Mallick, S.; Saha, B.; Bera, R.; Bhadra, R. Human placental lipid induces melanogenesis through p38 MAPK in B16F10 mouse melanoma. Pigment. Cell Res. 2005, 18, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Bertolotto, C.; Ortonne, J.P.; Ballotti, R. Inhibition of the phosphatidylinositol 3-kinase/p70(S6)-kinase pathway induces B16 melanoma cell differentiation. J. Biol. Chem. 1996, 271, 31824–31830. [Google Scholar] [CrossRef] [PubMed]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Khaled, M.; Larribere, L.; Bille, K.; Aberdam, E.; Ortonne, J.P.; Ballotti, R.; Bertolotto, C. Glycogen synthase kinase 3β is activated by cAMP and plays an active role in the regulation of melanogenesis. J. Biol. Chem. 2002, 277, 33690–33697. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Takemoto, C.; Kobayashi, I.; Watanabe, A.; Nobukuni, Y.; Fisher, D.E.; Tachibana, M. Ser298 of MITF, a mutation site in Waardenburg syndrome type 2, is a phosphorylation site with functional significance. Hum. Mol. Genet. 2000, 9, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, G.G.; Cook, T.A. Microtubules and signal transduction. Curr. Opin. Cell Biol. 1999, 11, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Abbe, P.; Mantoux, F.; Aberdam, E.; Peyssonnaux, C.; Eychene, A.; Ortonne, J.P.; Ballotti, R. Ras mediates the cAMP-dependent activation of extracellular signal-regulated kinases (ERKs) in melanocytes. EMBO J. 2000, 19, 2900–2910. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Rezzonico, R.; Durand-Clement, M.; Lallemand, D.; Ortonne, J.P.; Ballotti, R. Mitogen-activated protein kinase pathway and AP-1 are activated during cAMP-induced melanogenesis in B-16 melanoma cells. J. Biol. Chem. 1995, 270, 24315–24320. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Bertolotto, C.; Busca, R.; Brunet, A.; Pages, G.; Ortonne, J.P.; Ballotti, R. Inhibition of the mitogen-activated protein kinase pathway triggers B16 melanoma cell differentiation. J. Biol. Chem. 1998, 273, 9966–9970. [Google Scholar] [CrossRef] [PubMed]
- Hemesath, T.J.; Price, E.R.; Takemoto, C.; Badalian, T.; Fisher, D.E. MAP kinase links the transcription factor Microphthalmia to c-Kit signalling in melanocytes. Nature 1998, 391, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; No, J.K.; Lee, J.H.; Chung, H.Y. 4,4'-Dihydroxybiphenyl as a new potent tyrosinase inhibitor. Biol. Pharm. Bull. 2005, 28, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Hemesath, T.J.; Takemoto, C.M.; Horstmann, M.A.; Wells, A.G.; Price, E.R.; Fisher, D.Z.; Fisher, D.E. c-Kit triggers dual phosphorylations, which couple activation and degradation of the essential melanocyte factor Mi. Genes Dev. 2000, 14, 301–312. [Google Scholar] [PubMed]
- Xu, W.; Gong, L.; Haddad, M.M.; Bischof, O.; Campisi, J.; Yeh, E.T.; Medrano, E.E. Regulation of microphthalmia-associated transcription factor MITF protein levels by association with the ubiquitin-conjugating enzyme hUBC9. Exp. Cell Res. 2000, 255, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.Y.; Lin, Z.X.; Sviderskaya, E.; Cheng, C.H. Application of a combined sulphorhodamine B and melanin assay to the evaluation of Chinese medicines and their constituent compounds for hyperpigmentation treatment. J. Ethnopharmacol. 2010, 132, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, A.; Oka, M.; Okada, T.; Nakamura, S.; Ueyama, T.; Saito, N.; Hearing, V.J.; Ichihashi, M.; Nishigori, C. Down-regulation of melanogenesis by phospholipase D2 through ubiquitin proteasome-mediated degradation of tyrosinase. J. Biol. Chem. 2004, 279, 27774–27780. [Google Scholar] [CrossRef] [PubMed]
- Bertolotto, C.; Busca, R.; Abbe, P.; Bille, K.; Aberdam, E.; Ortonne, J.P.; Ballotti, R. Different cis-acting elements are involved in the regulation of TRP1 and TRP2 promoter activities by cyclic AMP: Pivotal role of M boxes (GTCATGTGCT) and of microphthalmia. Mol. Cell Biol. 1998, 18, 694–702. [Google Scholar] [PubMed]
- Jackson, I.J.; Chambers, D.M.; Budd, P.S.; Johnson, R. The tyrosinase-related protein-1 gene has a structure and promoter sequence very different from tyrosinase. Nucleic Acids Res. 1991, 19, 3799–3804. [Google Scholar] [CrossRef] [PubMed]
- Sturm, R.A.; O’Sullivan, B.J.; Box, N.F.; Smith, A.G.; Smit, S.E.; Puttick, E.R.; Parsons, P.G.; Dunn, I.S. Chromosomal structure of the human TYRP1 and TYRP2 loci and comparison of the tyrosinase-related protein gene family. Genomics 1995, 29, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, J.; Jung, E.; Lee, J.; Huh, S.; Hwang, H.; Kim, Y.; Park, D. 6-Benzylaminopurine stimulates melanogenesis via cAMP-independent activation of protein kinase A. Arch. Dermatol. Res. 2009, 301, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Khatib, S.; Nerya, O.; Musa, R.; Shmuel, M.; Tamir, S.; Vaya, J. Chalcones as potent tyrosinase inhibitors: The importance of a 2,4-substituted resorcinol moiety. Bioorg. Med. Chem. 2005, 13, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, S.Y.; Park, S.H.; Choi, Y.G.; Kwon, S.B.; Kim, M.K.; Na, J.I.; Youn, S.W.; Park, K.C. Inhibitory effects of 4-n-butylresorcinol on tyrosinase activity and melanin synthesis. Biol. Pharm. Bull. 2005, 28, 2216–2219. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J. Antimelanogenic and antioxidant properties of gallic acid. Biol. Pharm. Bull. 2007, 30, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Kondo, R.; Sakai, K. Inhibition of tyrosinase by flavonoids, stilbenes and related 4-substituted resorcinols: Structure-activity investigations. Planta Med. 2000, 66, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Kondo, R.; Sakai, K.; Takeda, N.; Nagahata, T.; Oniki, T. Novel vitamin E derivative with 4-substituted resorcinol moiety has both antioxidant and tyrosinase inhibitory properties. Lipids 2001, 3, 1321–1326. [Google Scholar] [CrossRef]
- Shimizu, K.; Yasutake, S.; Kondo, R. A new stilbene with tyrosinase inhibitory activity from Chlorophora excelsa. Chem. Pharm. Bull. 2003, 51, 318–319. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.E.; Ivanov, A.S.; Kamyshanskaya, N.S.; Kirkel, A.Z.; Moskvitina, T.A.; Gorkin, V.Z.; Li, N.Y.; Marshakov, V.Y. Interaction of indole derivatives with monoamine oxidase A and B. Studies on the structure-inhibitory activity relationship. Biochem. Mol. Biol. Int. 1995, 36, 113–122. [Google Scholar]
- Stupans, I.; Ryan, A.J. In vitro inhibition of 3-methylcholanthrene-induced rat hepatic aryl hydrocarbon hydroxylase by 8-acyl-7-hydroxycoumarins. Structure-activity relationships and metabolite profiles. Biochem. Pharmacol. 1984, 33, 131–139. [Google Scholar]
- Lam, R.Y.; Woo, A.Y.; Leung, P.S.; Cheng, C.H. Antioxidant actions of phenolic compounds found in dietary plants on low-density lipoprotein and erythrocytes in vitro. J. Am. Coll. Nutr. 2007, 26, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.M. The use of standards for melonyldialdehyde. Anal. Biochem. 1975, 69, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.J. Oxidative stress status and cancer: Methodology applicable for human studies. Free Radic. Biol. Med. 1991, 10, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Pan, X.; Li, C.; Zhou, C.; Deng, F.; Li, T. Density functional theory calculations for resveratrol. Bioorg. Med. Chem. Lett. 2003, 13, 1869–1871. [Google Scholar] [CrossRef] [PubMed]
- Nakao, K.; Shimizu, R.; Kubota, H.; Yasuhara, M.; Hashimura, Y.; Suzuki, T.; Fujita, T.; Ohmizu, H. Quantitative structure-activity analyses of novel hydroxyphenylurea derivatives as antioxidants. Bioorg. Med. Chem. 1998, 6, 849–868. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Hsu, F.L.; Chen, C.S.; Chern, J.W.; Lee, M.H. Constituents from the Formosan apple reduce tyrosinase activity in human epidermal melanocytes. Phytochemistry 2007, 68, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, D.S.; Kim, W.G.; Ryoo, I.J.; Lee, D.H.; Huh, C.H.; Youn, S.W.; Yoo, I.D.; Park, K.C. Terrein: A new melanogenesis inhibitor and its mechanism. Cell Mol. Life Sci. 2004, 61, 2878–2885. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Stockley, P.G.; Martin, W.J., II. An improved Western blotting technique effectively reduces background. Electrophoresis 2002, 23, 2373–2376. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.G.; Bae, E.J.; Kim, D.S.; Park, S.H.; Kwon, S.B.; Na, J.I.; Park, K.C. Differential regulation of melanosomal proteins after hinokitiol treatment. J. Dermatol. Sci. 2006, 43, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.D.; Yang, J.Y.; Song, E.S.; Par, J.W. Stimulation of melanogenesis by glycyrrhizin in B16 melanoma cells. Exp. Mol. Med. 2001, 33, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.H.; Kim, H.T.; Yoon, H.Y.; Kwon, K.B.; Choi, I.W.; Jung, S.H.; Kim, H.U.; Park, B.H.; Park, J.W. Effect of xanthohumol on melanogenesis in B16 melanoma cells. Exp. Mol. Med. 2008, 40, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, T.; Fabry, M.E.; Kaul, D.K. Antisickling property of fetal hemoglobin enhances nitric oxide bioavailability and ameliorates organ oxidative stress in transgenic-knockout sickle mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R394–R402. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, S.H.; Bryan, N. Inhibition of LDL oxidation and myeloperoxide dependent tyrosyl radical formation by the selective estrogen receptor modulator raloxifene (LY139481 HCL). Atherosclerosis 1996, 126, 65–75. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lam, R.Y.Y.; Lin, Z.-X.; Sviderskaya, E.V.; Cheng, C.H.K. Mechanistic Studies of Anti-Hyperpigmentary Compounds: Elucidating Their Inhibitory and Regulatory Actions. Int. J. Mol. Sci. 2014, 15, 14649-14668. https://doi.org/10.3390/ijms150814649
Lam RYY, Lin Z-X, Sviderskaya EV, Cheng CHK. Mechanistic Studies of Anti-Hyperpigmentary Compounds: Elucidating Their Inhibitory and Regulatory Actions. International Journal of Molecular Sciences. 2014; 15(8):14649-14668. https://doi.org/10.3390/ijms150814649
Chicago/Turabian StyleLam, Rosanna Y. Y., Zhi-Xiu Lin, Elena V. Sviderskaya, and Christopher H. K. Cheng. 2014. "Mechanistic Studies of Anti-Hyperpigmentary Compounds: Elucidating Their Inhibitory and Regulatory Actions" International Journal of Molecular Sciences 15, no. 8: 14649-14668. https://doi.org/10.3390/ijms150814649
APA StyleLam, R. Y. Y., Lin, Z.-X., Sviderskaya, E. V., & Cheng, C. H. K. (2014). Mechanistic Studies of Anti-Hyperpigmentary Compounds: Elucidating Their Inhibitory and Regulatory Actions. International Journal of Molecular Sciences, 15(8), 14649-14668. https://doi.org/10.3390/ijms150814649