
The Oxidative Fermentation of Ethanol in Gluconacetobacter diazotrophicus Is a Two-Step Pathway Catalyzed by a Single Enzyme: Alcohol-Aldehyde Dehydrogenase (ADHa)

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
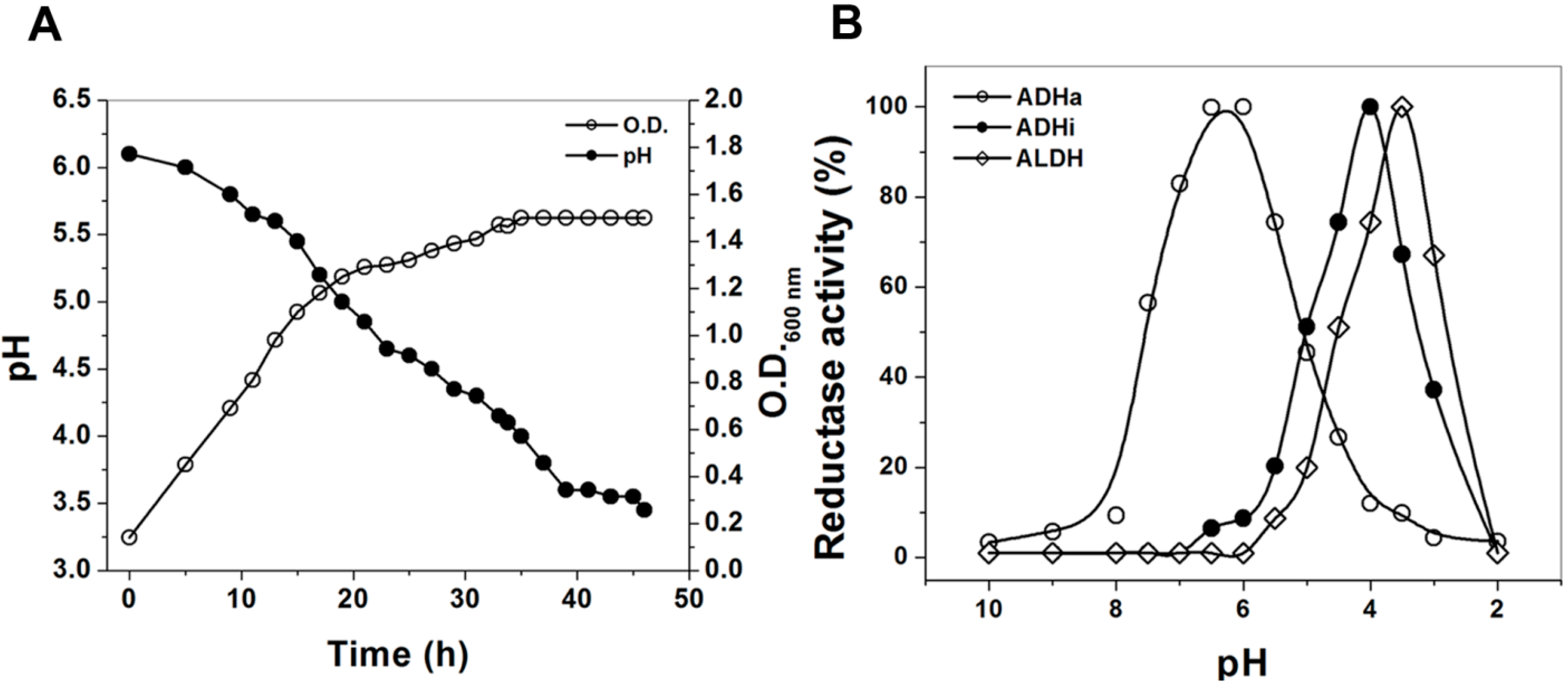
2. Results
2.1. Purification of the ADHa from Ga. diazotrophicus
2.2. N-Terminal Sequence Analysis
2.3. Kinetic Characterization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetic Constants | Ethanol | Acetaldehyde | ||
|---|---|---|---|---|
| 1 Dehydrogenase | Oxidase Activity | 1 Dehydrogenase | Oxidase Activity | |
| Km (M) | 4.6 × 10−4 | 7.6 × 10−5 | 3.7 × 10−4 | 1.1 × 10−3 |
| kcat (s−1) | 1.6 × 105 | 1.4 × 105 | 9.3 × 104 | 1.1 × 105 |
| kcat/Km (M−1·s−1) | 3.6 × 108 | 1.8 × 109 | 2.5 × 107 | 1.0 × 108 |
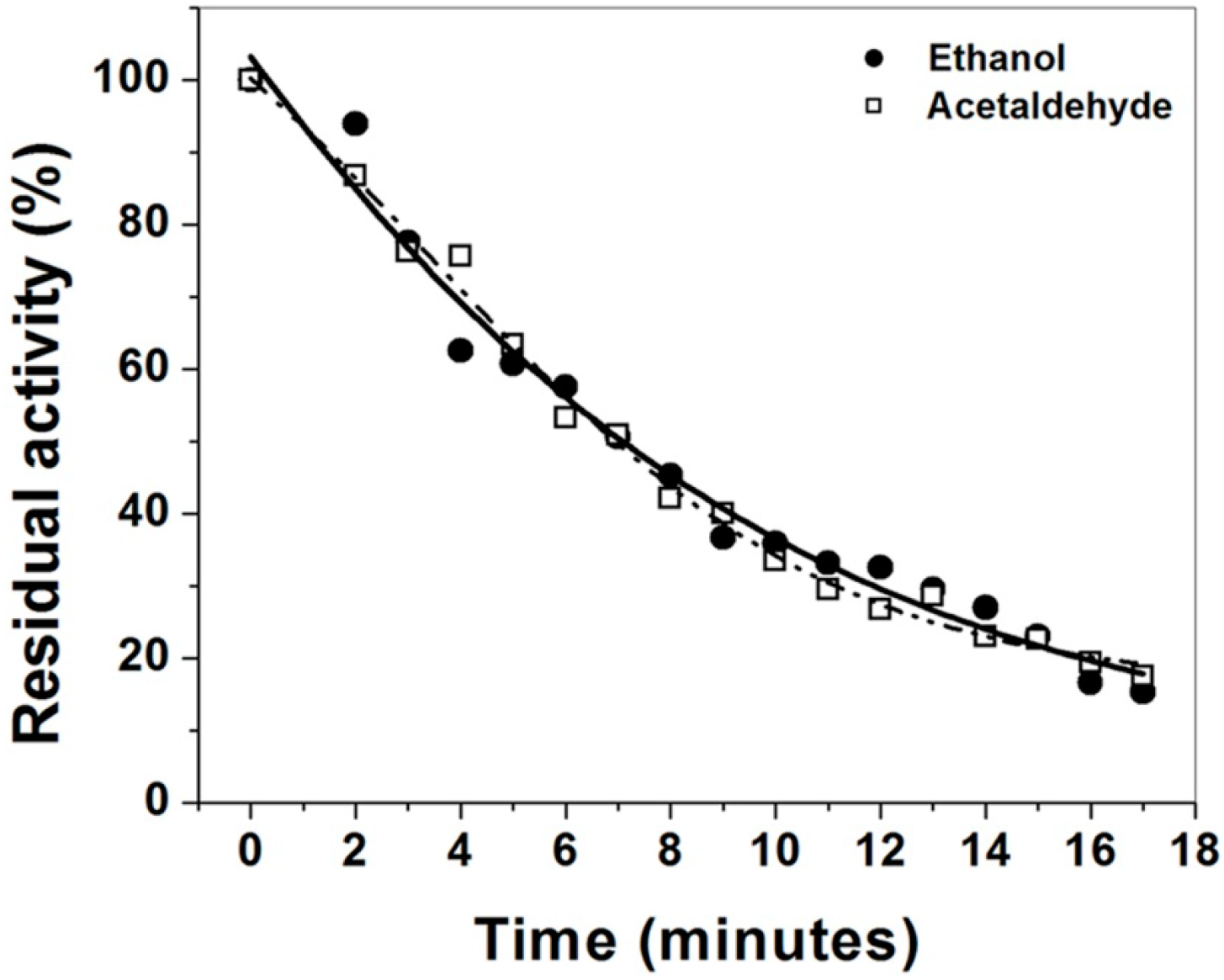
2.4. Thermal Denaturation Curves of the Purified ADHa Enzyme

2.5. Determination of Ethanol and Acetate as Final Products of the Reaction
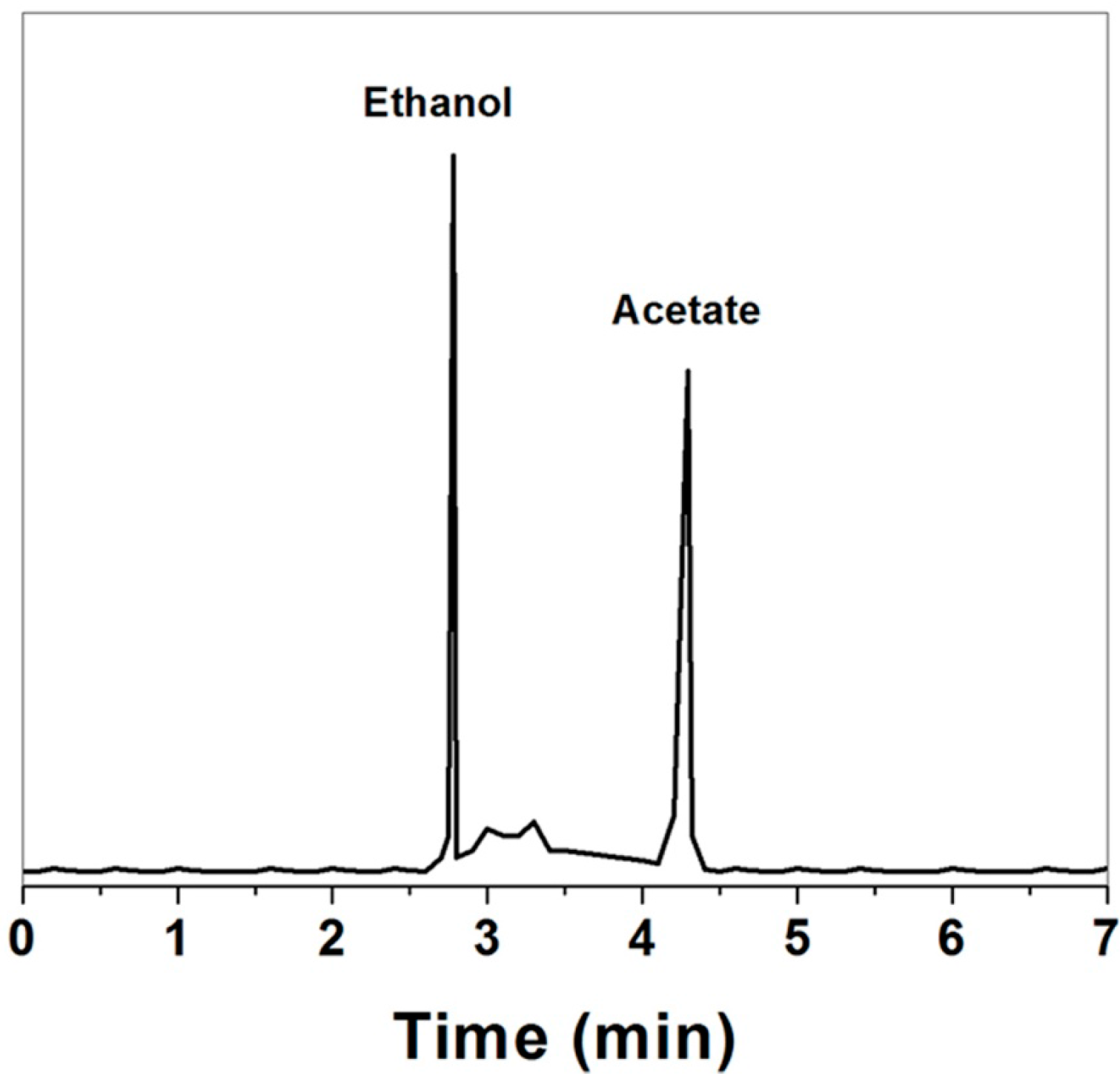
2.5.1. Gas Chromatography-Mass Spectrometry (GC-MS)

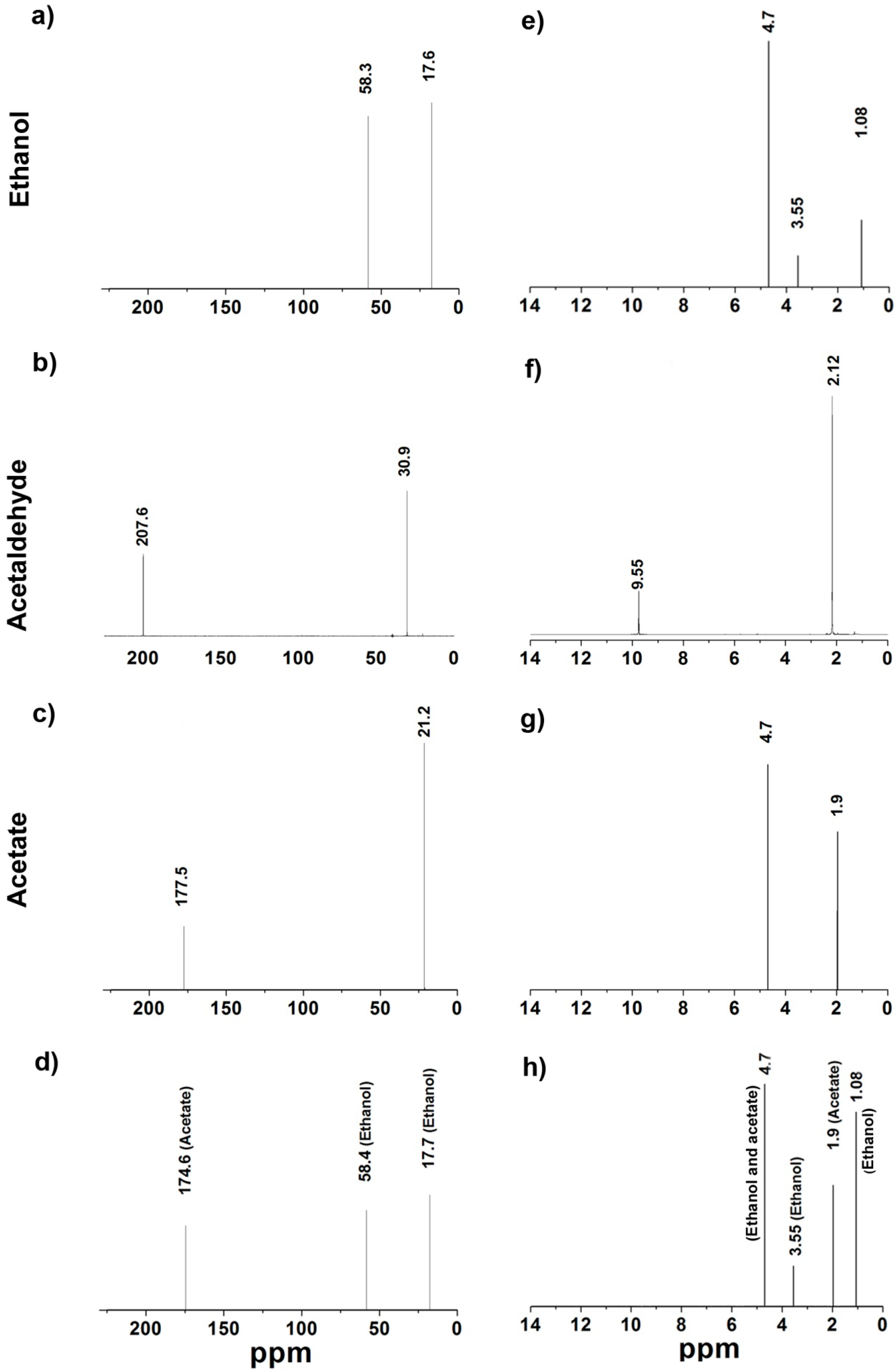
2.5.2. Nuclear Magnetic Resonance (NMR)

2.5.3. Acetaldehyde-Semicarbazone Formation with Radiolabeled Ethanol
| Assay | Acetaldehyde-Semicarbazone Complex (CPM) | Final Reaction Mixture Catalyzed by the ADHa (CPM) |
|---|---|---|
| Control (−) | 13,285 | 386,175 |
| Control (+) (ADH-NADH) | 89,640 | 310,360 |
| ADHa | 12,525 | 383,400 |

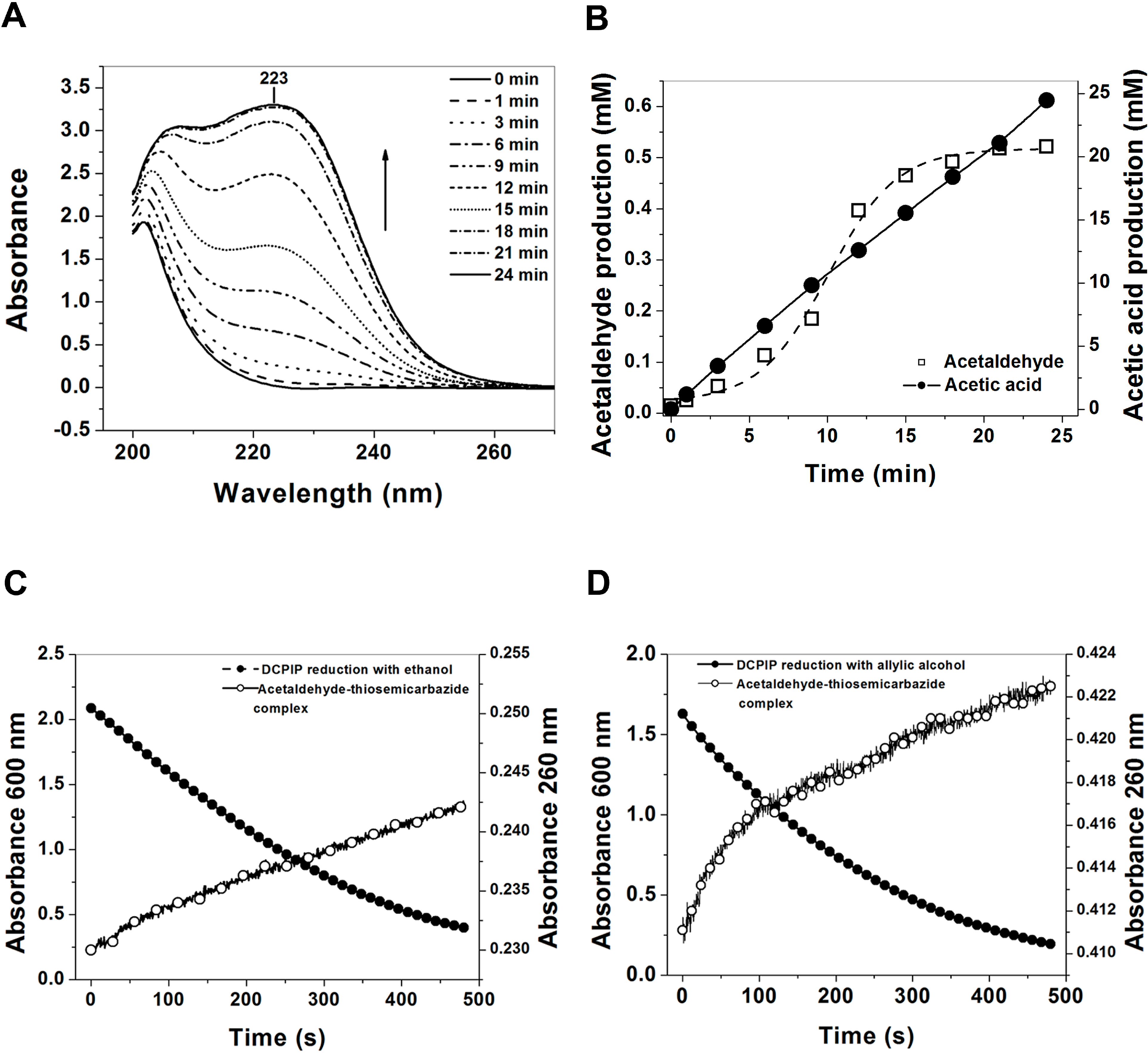
2.5.4. Coupled Assay of 2,6-Dichlorophenolindophenol (DCPIP) with Semicarbazide and Thiosemicarbazide
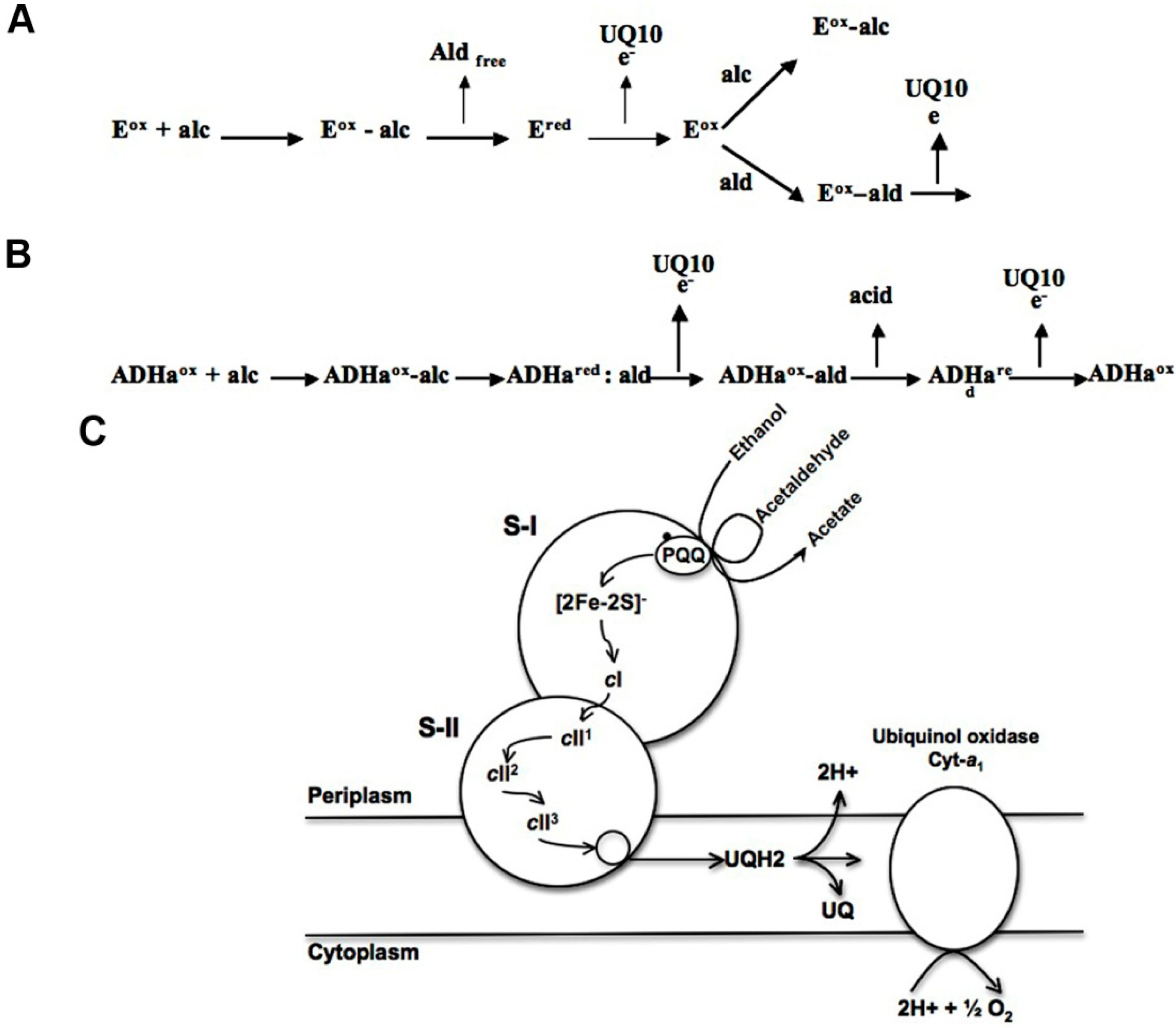
3. Discussion


4. Experimental Section
4.1. Strain and Growth Conditions
4.2. Purification of the ADHa from Ga. diazotrophicus
4.3. Spectroscopy Studies
4.4. Thermal Inactivation of the Purified Enzyme ADHa
4.5. Enzymatic Activity Assays
4.6. Determination of Ethanol and Acetate as Final Products of the Reaction
4.6.1. Gas Chromatography-Mass Spectrometry (GC-MS)
4.6.2. Nuclear Magnetic Resonance (NMR)
4.6.3. Acetaldehyde-Semicarbazone Formation with Radiolabeled Ethanol
4.6.4. Coupled Assay of 2,6-Dichlorophenolindophenol (DCPIP) with Carbazide and Thiosemicarbazide
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yuzo, Y.; Pattaraporn, Y. Genera and species in acetic acid bacteria. Int. J. Food. Microbiol. 2008, 125, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Sengun, I.Y.; Karabiyikli, S. Importance of acetic acid bacteria in food industry. Food Control 2011, 22, 647–656. [Google Scholar] [CrossRef]
- Gullo, M.; Verzelloni, E.; Canonico, M. Aerobic submerged fermentation by acetic acid bacteria for vinegar production: Process and biotechnological aspects. Proc. Biochem. 2014, 49, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Gillis, M.; Kersters, K.; Hoste, B.; Janssens, D.; Kroppenstedt, R.M.; Stephan, M.P.; Teixeria, K.R.S.; Dobereiner, J.; de Ley, J. Acetobacter diazotrophicus sp. nov., a nitrogen-fixing acetic bacterium associated with sugarcane. Int. J. Syst. Bacteriol. 1989, 39, 361–364. [Google Scholar] [CrossRef]
- Yamada, Y.; Hoshino, K.I.; Ishikawa, T. The phylogeny of acetic acid bacteria based on the partial sequences of 16S ribosomal RNA: The elevation of the subgenus Gluconoacetobacter to the generic level. Biosci. Biotechnol. Biochem. 1997, 61, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Flores-Encarnación, M.; Contreras-Zentella, M.; Soto-Urzúa, L.; Aguilar, G.R.; Baca, B.E.; Escamilla, J.E. The respiratory system and diazotrophic activity of Acetobacter diazotrophicus PAL5. J. Bacteriol. 1999, 181, 6987–6995. [Google Scholar] [PubMed]
- Matsushita, K.; Takaki, Y.; Shinagawa, E.; Ameyama, M.; Adachi, O. Ethanol oxidase respiratory chain of acetic acid bacteria. Reactivity with ubiquinone of pyrroloquinoline quinone-dependent alcohol dehydrogenases purified from Acetobacter aceti and Gluconobacter suboxydans. Biosci. Biotechnol. Biochem. 1992, 56, 304–310. [Google Scholar]
- Matsushita, K.; Yakushi, T.; Takaki, Y.; Toyama, H.; Adachi, O. Generation mechanism and purification of an inactive form convertible in vivo to the active form of quinoprotein alcohol dehydrogenase in Gluconobacter suboxydans. J. Bacteriol. 1995, 177, 6552–6559. [Google Scholar] [PubMed]
- Carrell, A.A.; Frank, A.C. Pinus flexilis and Picea engelmannii share a simple and consistent needle endophyte microbiota with a potential role in nitrogen fixation. Front. Microbiol. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schrover, J.M.; Frank, J.; van Wielink, J.E.; Duine, J.A. Quaternary structure of quinoprotein ethanol dehydrogenase from Pseudomonas aeruginosa and its reoxidation with a novel cytochrome c from this organism. Biochem. J. 1993, 290, 123–127. [Google Scholar] [PubMed]
- Jongejan, A.; Jongejan, J.A.; Duine, J.A. Homology model of the quinohaemoprotein alcohol dehydrogenase from Comamonas testosteroni. Protein Eng. 1998, 11, 185–98. [Google Scholar] [CrossRef] [PubMed]
- Groen, B.W.; van Kleef, M.A.G.; Duine, J.A. Quinohaemoprotein alcohol dehydrogenase apoenzyme from Pseudomonas testosteroni. Biochem. J. 1986, 234, 611–615. [Google Scholar] [PubMed]
- De Jong, G.A.H.; Geerlof, A.; Stoorvogel, J.; Jongejan, J.A.; de Vries, S.; Duine, J.A. Quinohaemoprotein ethanol dehydrogenase from Comamonas testosteroni purification, characterization, and reconstitution of the apoenzyme with pyrroloquinoline quinone analogues. Eur. J. Biochem. 1995, 230, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Manzo, S.; del Arenal-Mena, I.P.; Escamilla, J.E. The inactive and active forms of the pyrroloquinoline quinone-alcohol dehydrogenase of Gluconacetobacter diazotrophicus: A comparative study. Acetic. Acid Bact. 2013, 2. [Google Scholar] [CrossRef]
- Tayama, K.; Fukaya, M.; Okumura, H.; Kawamura, Y.; Beppu, T. Purification and characterization of membrane-bound alcohol dehydogenase from Acetobacter polyoxogenes. Appl. Microbiol. Biotechnol. 1989, 32, 181–185. [Google Scholar] [CrossRef]
- Chávez-Pacheco, J.L.; Contreras-Zentella, M.L.; Membrillo-Hernández, J.; Arreguín-Espinoza, R.; Mendoza-Hernandez, G.; Gómez-Manzo, S.; Escamilla, J.E. The quinohaemoprotein alcohol dehydrogenase from Gluconacetobacter xylinus: Molecular and catalytic properties. Archiv. Microbiol. 2010, 192, 703–713. [Google Scholar] [CrossRef]
- Shinagawa, E.; Toyama, H.; Matsushita, K.; Tuitemwong, P.; Theeragool, G.; Adachi, O. A novel type of formaldehyde-oxidizing enzyme from the membrane of Acetobacter sp. SKU 14. Biosci. Biotechnol. Biochem. 2006, 70, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Manzo, S.; Contreras-Zentella, M.; González-Valdez, A.; Sosa-Torres, M.; Arreguín-Espinoza, R.; Escamilla-Marván, E. The PQQ-alcohol deshidrogenase from Gluconacetobacter diazotrophicus. Int. J. Food Microbiol. 2008, 125, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Kanchanarach, W.; Theeragool, G.; Yakushi, T.; Toyama, H.; Adachi, O.; Matsushita, K. Characterization of thermotolerant Acetobacter pasteurianus strains and their quinoprotein alcohol dehydrogenases. Appl. Microbiol. Biotechnol. 2010, 85, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Manzo, S.; González-Valdez, A.A.; Oria-Hernández, J.; Reyes-Vivas, H.; Arreguín-Espinosa, R.; Kroneck, P.M.H.; Sosa-Torres, M.E.; Escamilla, J.E. The inactive and active forms of the PQQ-alcohol dehydrogenase of Gluconacetobacter diazotrophicus differ in the redox state of their respective prosthetic groups. FEMS. Microbiol. Lett. 2012, 328, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Manzo, S.; Solano-Peralta, A.; Saucedo-Vázquez, J.P.; Escamilla-Marván, J.E.; Kroneck, P.M.H.; Sosa-Torres, M.E. The membrane-bound quinohemoprotein alcohol dehydrogenase from Gluconacetobacter diazotrophicus PAL5 carries a [2Fe–2S] cluster. Biochemistry 2010, 49, 2409–2415. [Google Scholar] [CrossRef] [PubMed]
- Djurdjić, V.; Stojanović, S. A method for spectrophotometric determination of acetaldehyde with thiosemicarbazide in blood. Facta Univ. 1998, 1, 129–134. [Google Scholar]
- Ameyama, M.; Adachi, O. Alcohol dehydrogenase from acetic acid bacteria membrane-bound. Methods Enzymol. 1982, 89, 450–457. [Google Scholar]
- Adachi, O.; Miyagawa, E.; Shinagawa, E.; Matsushita, K.; Ameyama, M. Purification and propierties of particulate alcohol dehydrogenase from Acetobacter aceti. Agric. Biol. Chem. 1978, 42, 2331–2340. [Google Scholar] [CrossRef]
- Gómez-Manzo, S.; Chavez-Pacheco, J.L.; Contreras-Zentella, M.; Sosa-Torres, M.E.; Arreguín-Espinosa, R.; Pérez de la Mora, M.; Membrillo-Hernández, J.; Escamilla, J.E. Molecular and catalytic properties of the aldehyde dehydrogenase of Gluconacetobacter diazotrophicus, a quinoheme protein containing pyrroloquinoline quinone, cytochrome b, and cytochrome c. J. Bacteriol. 2010, 192, 5718–5724. [Google Scholar] [CrossRef] [PubMed]
- Ameyama, M.; Osada, K.; Shinagawa, E.; Matsushita, K.; Adachi, O. Purification and characterization of aldehyde dehydrogenase of Acetobacter aceti. Agric. Biol. Chem. 1981, 45, 1889–1890. [Google Scholar] [CrossRef]
- Adachi, O.; Tayama, K.; Shinagawa, E.; Matsushita, K.; Ameyama, M. Purification and characterization of membrane-bound aldehyde dehydrogenase from Gluconobacter suboxydans. Agric. Biol. Chem. 1980, 44, 503–515. [Google Scholar] [CrossRef]
- Hommel, R.; Kleber, H.P. Properties of the quinoprotein aldehyde dehydrogenase from Acetobacter rances. J. Gen. Microbiol. 1990, 136, 1705–1711. [Google Scholar] [CrossRef]
- Nair, R.V.; Bennett, G.N.; Papoutsakis, E.T. Molecular characterization of an aldehyde/alcohol dehydrogenase gene from Clostridium acetobutylicum ATCC 824. J. Bacteriol. 1994, 176, 871–885. [Google Scholar] [PubMed]
- Goodlove, P.; Cunningham, P.R.; Parker, J.; Clark, D.P. Cloning and sequence analysis of the fermentative alcohol-dehydrogenase-encoding gene of Escherichia coli. Gene 1989, 85, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Membrillo-Hernández, J.; Echave, P.; Cabiscol, E.; Tamarit, J.; Ros, J.; Lin, E.C.C. Evolution of the adhE gene product of Escherichia coli from a functional reductase to a dehydrogenase. J. Biol. Chem. 2000, 275, 33869–33875. [Google Scholar] [CrossRef] [PubMed]
- Atteia, A.; van Lis, R.; Mendoza-Hernández, G.; Henze, K.; Martin, W.; Riveros-Rosas, H.; González-Halphen, D. Bifunctional aldehyde/alcohol dehydrogenase (ADHE) in chlorophyte algal mitochondria. Plant Mol. Biol. 2003, 53, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.K.; Jeong, D.W.; Lee, J.M.; Kim, M.J.; Lee, J.H.; Chang, H.C.; Kim, J.H.; Lee, H.J. Cloning and characterization of the bifunctional alcohol/acetaldehyde dehydrogenase gene (adhE) in Leuconostoc mesenteroides isolated from kimchi. Biotechnol. Lett. 2005, 27, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Reis, V.M.; Olivares, F.L.; Döbereiner, J. Improved methodology for isolation of Acetobacter diazotrophicus and confirmation of its endophytic habitat. World J. Microbiol. Biotechnol. 1994, 10, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, J.E.; Ramirez, R.; del Arenal, I.P.; Zarzosa, G.; Linares, V. Expression of cytochrome oxidases in Bacillus cereus; effects of oxygen tension and carbon source. J. Gen. Microbiol. 1987, 133, 3549–3555. [Google Scholar]
- Goodhew, C.F.; Brown, K.R.; Pettigrew, G.W. Haem staining in gels, as useful tool in the study of bacterial c-type cytochromes. Biochim. Biophys. Acta 1986, 852, 288–294. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; Haas, S.M.; Tolbert, N.E.; Bieber, L.L. Protein determination in membrane and lipoprotein samples: Manual and automated procedures. Methods Enzymol. 1981, 72, 296–303. [Google Scholar] [PubMed]
- Duine, J.A.; Jongejan, J.A. Quinoproteins, enzymes with pyrrolo-quinoline quinone as cofactor. Annu. Rev. Biochem. 1989, 58, 403–426. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Manzo, S.; Escamilla, J.E.; González-Valdez, A.; López-Velázquez, G.; Vanoye-Carlo, A.; Marcial-Quino, J.; De la Mora-de la Mora, I.; Garcia-Torres, I.; Enríquez-Flores, S.; Contreras-Zentella, M.L.; et al. The Oxidative Fermentation of Ethanol in Gluconacetobacter diazotrophicus Is a Two-Step Pathway Catalyzed by a Single Enzyme: Alcohol-Aldehyde Dehydrogenase (ADHa). Int. J. Mol. Sci. 2015, 16, 1293-1311. https://doi.org/10.3390/ijms16011293
Gómez-Manzo S, Escamilla JE, González-Valdez A, López-Velázquez G, Vanoye-Carlo A, Marcial-Quino J, De la Mora-de la Mora I, Garcia-Torres I, Enríquez-Flores S, Contreras-Zentella ML, et al. The Oxidative Fermentation of Ethanol in Gluconacetobacter diazotrophicus Is a Two-Step Pathway Catalyzed by a Single Enzyme: Alcohol-Aldehyde Dehydrogenase (ADHa). International Journal of Molecular Sciences. 2015; 16(1):1293-1311. https://doi.org/10.3390/ijms16011293
Chicago/Turabian StyleGómez-Manzo, Saúl, José E. Escamilla, Abigail González-Valdez, Gabriel López-Velázquez, América Vanoye-Carlo, Jaime Marcial-Quino, Ignacio De la Mora-de la Mora, Itzhel Garcia-Torres, Sergio Enríquez-Flores, Martha Lucinda Contreras-Zentella, and et al. 2015. "The Oxidative Fermentation of Ethanol in Gluconacetobacter diazotrophicus Is a Two-Step Pathway Catalyzed by a Single Enzyme: Alcohol-Aldehyde Dehydrogenase (ADHa)" International Journal of Molecular Sciences 16, no. 1: 1293-1311. https://doi.org/10.3390/ijms16011293
APA StyleGómez-Manzo, S., Escamilla, J. E., González-Valdez, A., López-Velázquez, G., Vanoye-Carlo, A., Marcial-Quino, J., De la Mora-de la Mora, I., Garcia-Torres, I., Enríquez-Flores, S., Contreras-Zentella, M. L., Arreguín-Espinosa, R., Kroneck, P. M. H., & Sosa-Torres, M. E. (2015). The Oxidative Fermentation of Ethanol in Gluconacetobacter diazotrophicus Is a Two-Step Pathway Catalyzed by a Single Enzyme: Alcohol-Aldehyde Dehydrogenase (ADHa). International Journal of Molecular Sciences, 16(1), 1293-1311. https://doi.org/10.3390/ijms16011293