
Specificity Protein 1 Regulates Gene Expression Related to Fatty Acid Metabolism in Goat Mammary Epithelial Cells

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
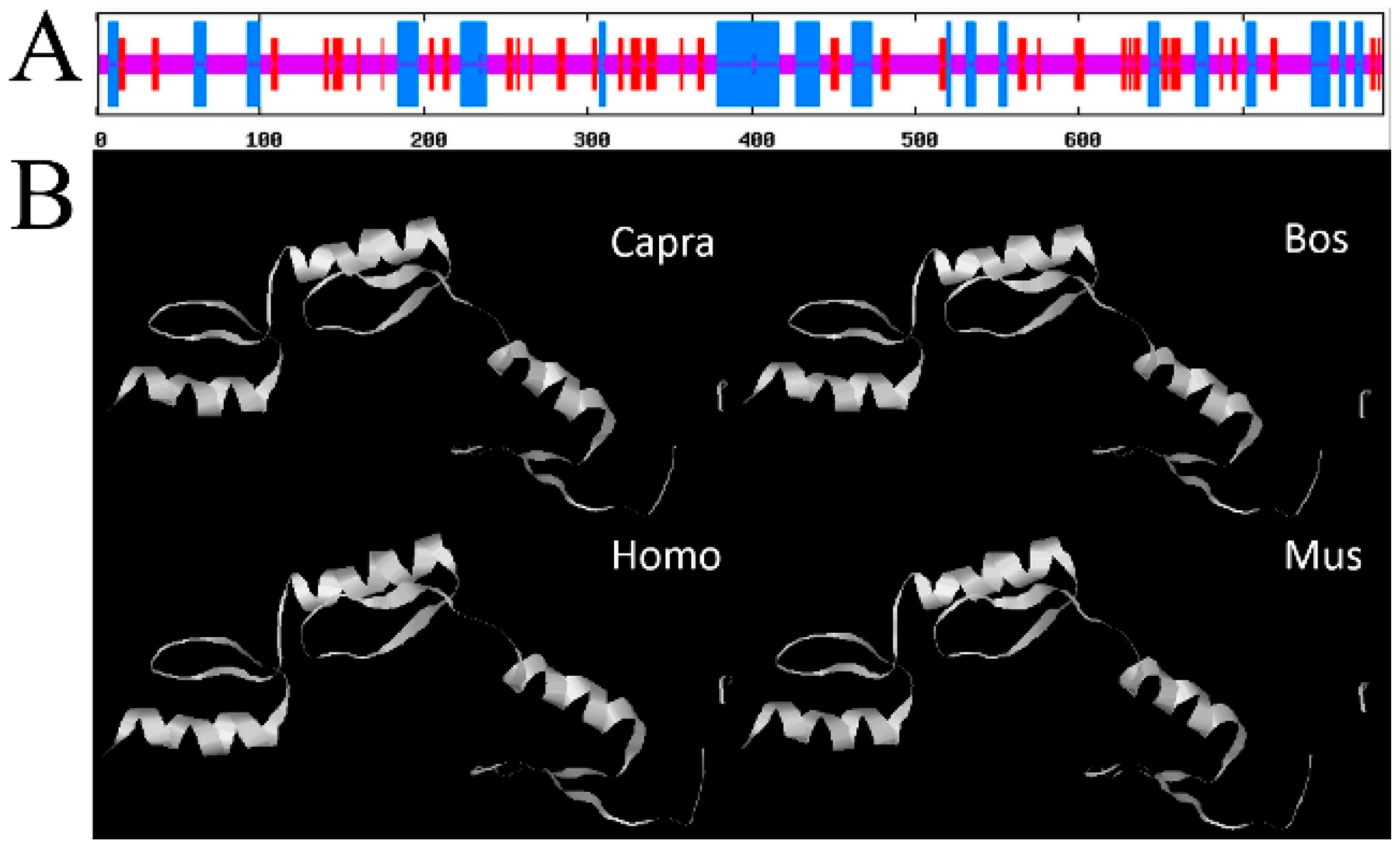
2.1. Characterization of SP1 cDNA from Goat Mammary Gland

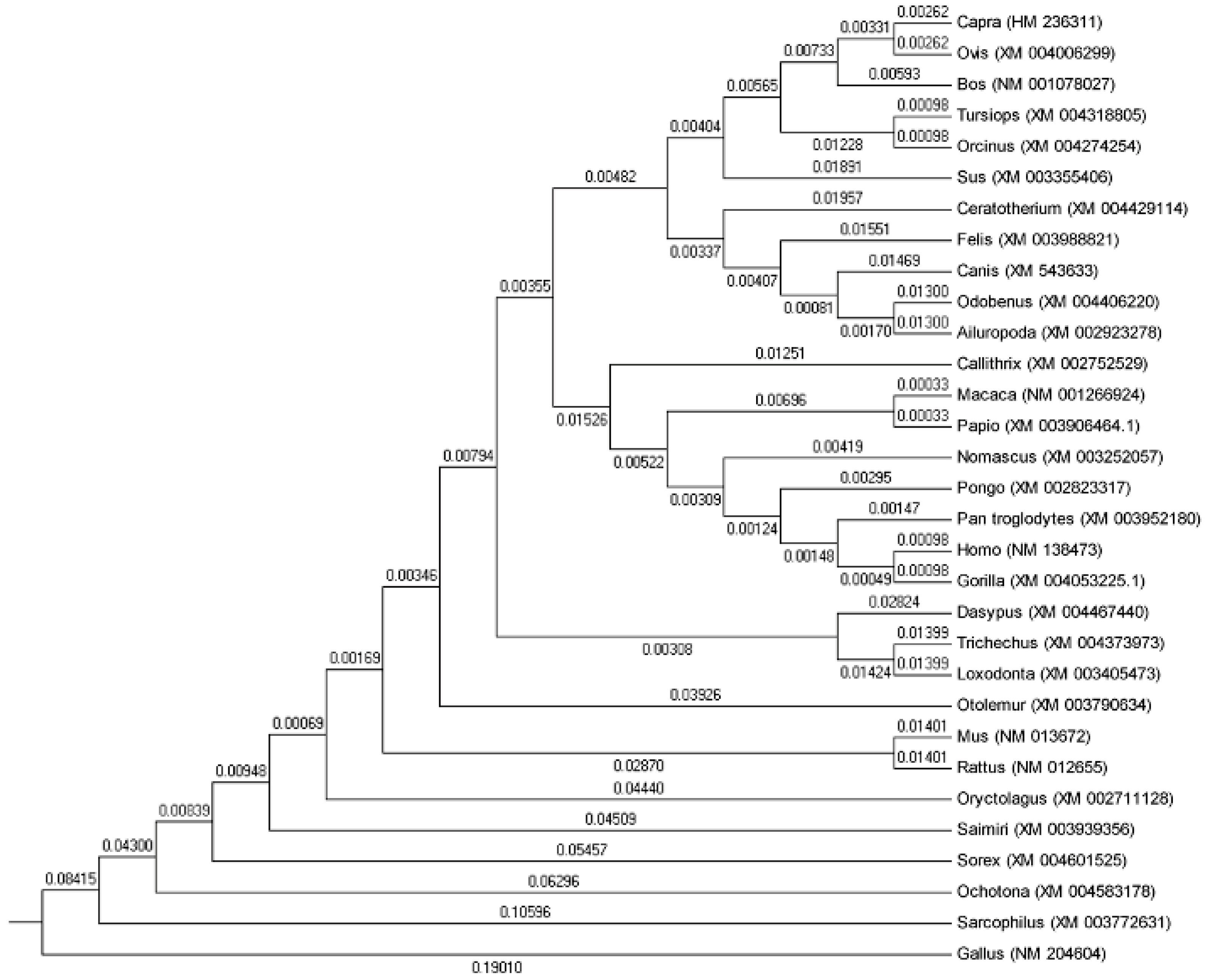
2.2. Phylogenetic Analysis Based on SP1 Amino Acid Sequences

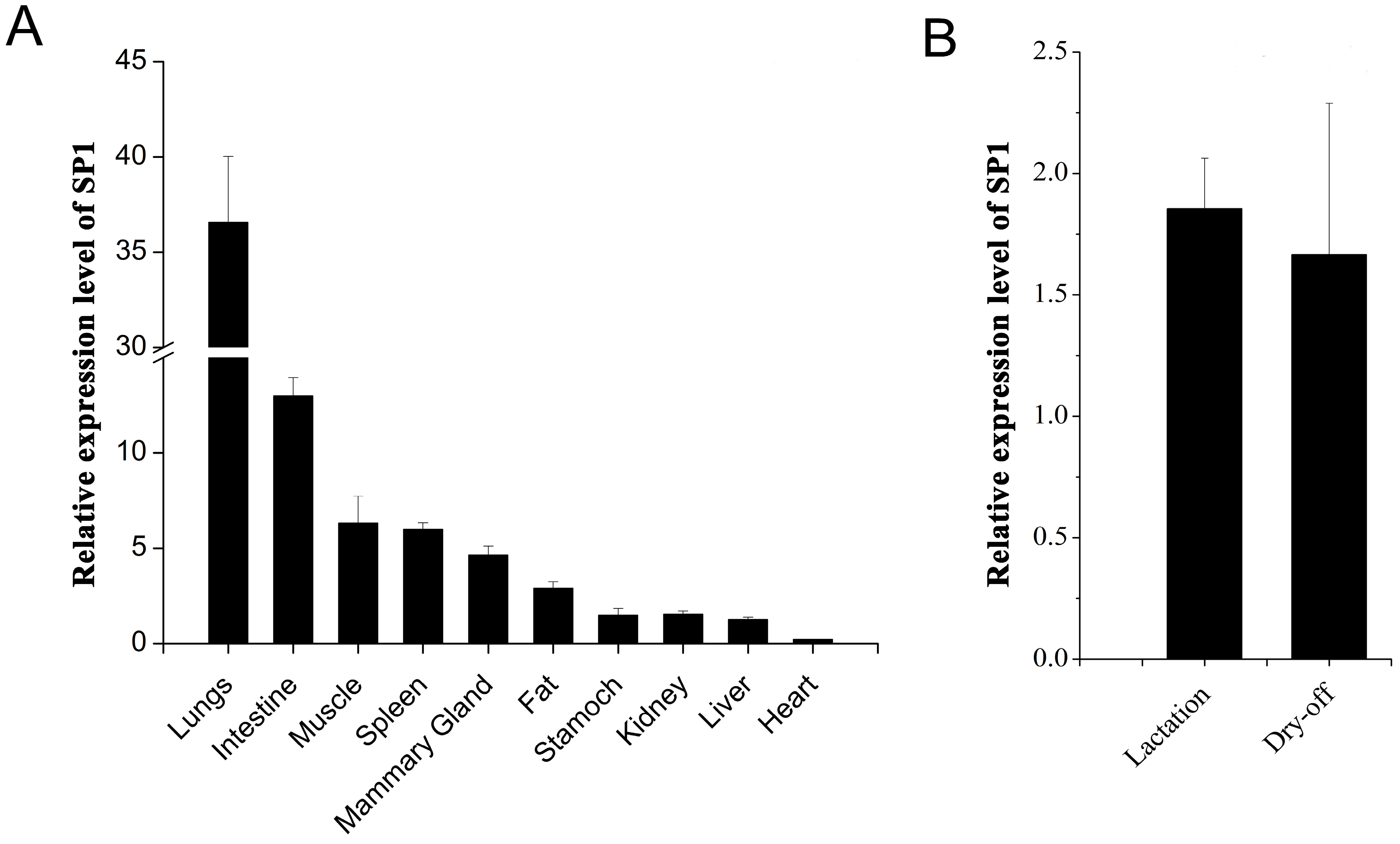
2.3. Analysis of the Expression Profile of Goat SP1

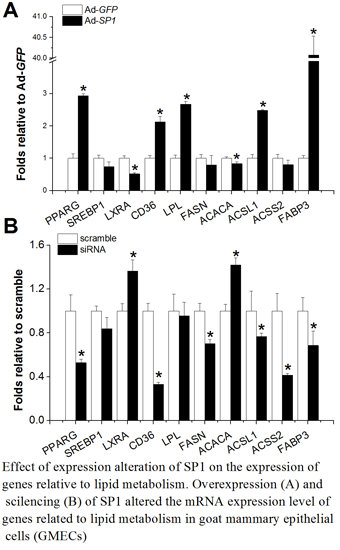
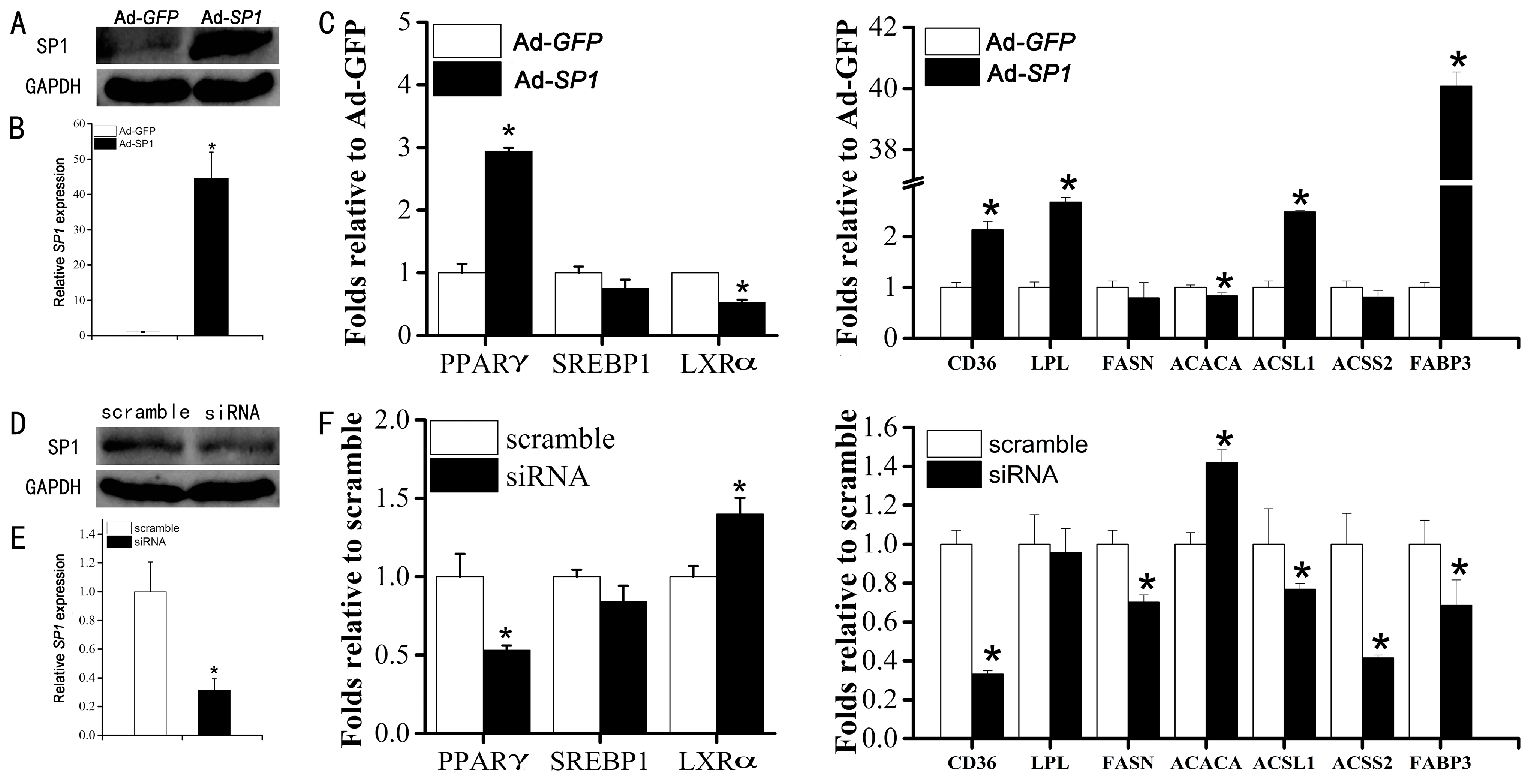
2.4. SP1 Is Involved in Lipid Metabolism Regulation

3. Discussion
4. Experimental Section
4.1. Animals and Samples Collection
4.2. Total RNA Extraction and Real-Time Quantitative PCR (RT-qPCR)
4.3. cDNA Cloning of SP1 from Dairy Goat Mammary Gland
4.4. Sequence Analyses
4.5. siRNA Synthesis and Adenovirus Generation
4.6. Cell Culture and Treatment
4.7. Western Blot Analyses
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Haenlein, G.F.W. Goat milk in human nutrition. Small Rumin. Res. 2004, 51, 155–163. [Google Scholar] [CrossRef]
- Williams, C. Dietary fatty acids and human health. Ann. Zootech. 2000, 49, 165–180. [Google Scholar] [CrossRef]
- Shi, H.; Luo, J.; Zhu, J.; Li, J.; Sun, Y.; Lin, X.; Zhang, L.; Yao, D.; Shi, H. PPARG regulates genes involved in triacylglycerol synthesis and secretion in mammary gland epithelial cells of dairy goats. PPAR Res. 2013, 2013, 310948:1–310948:10. [Google Scholar] [CrossRef]
- Harvatine, K.J.; Boisclair, Y.R.; Bauman, D.E. Recent advances in the regulation of milk fat synthesis. Animal 2009, 3, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Oppi-Williams, C.; Suagee, J.K.; Corl, B.A. Regulation of lipid synthesis by liver X receptor a and sterol regulatory element-binding protein 1 in mammary epithelial cells. J. Dairy Sci. 2013, 96, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.S.; Majumdar, G.; Martinez-Hernandez, A.; Raghow, R. A critical role of sp1 transcription factor in regulating gene expression in response to insulin and other hormones. Life Sci. 2008, 83, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xie, K. Crosstalk of Sp1 and Stat3 signaling in pancreatic cancer pathogenesis. Cytokine Growth Factor Rev. 2012, 23, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Abdelrahim, M. Sp transcription factor family and its role in cancer. Eur. J. Cancer 2005, 41, 2438–2448. [Google Scholar] [CrossRef] [PubMed]
- Bouwman, P.; Philipsen, S. Regulation of the activity of Sp1-related transcription factors. Mol. Cell. Endocrinol. 2002, 195, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Gill, G.; Pascal, E.; Tseng, Z.H.; Tjian, R. A glutamine-rich hydrophobic patch in transcription factor SP1 contacts the dTAFII110 component of the drosophila TFIID complex and mediates transcriptional activation. Proc. Natl. Acad. Sci. USA 1994, 91, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Hoey, T.; Weinzierl, R.O.; Gill, G.; Chen, J.L.; Dynlacht, B.D.; Tjian, R. Molecular cloning and functional analysis of drosophila TAF110 reveal properties expected of coactivators. Cell 1993, 72, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.L.; Coleman, R.A.; Grob, P.; King, D.S.; Florens, L.; Washburn, M.P.; Geles, K.G.; Yang, J.L.; Ramey, V.; Nogales, E.; et al. Structural changes in TAF4b-TFIID correlate with promoter selectivity. Mol. Cell 2008, 29, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.-N.; Kim, Y.-S.; Choi, W.-I.; Koh, D.-I.; Kim, M.-K.; Yoon, J.-H.; Kim, M.-Y.; Hur, B.; Paik, P.D.-H.; Hur, M.-W. Kr-pok increases FASN expression by modulating the DNA binding of SREBP-1c and Sp1 at the proximal promoter. J. Lipid Res. 2012, 53, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Archer, M.C. Sp1 coordinately regulates de novo lipogenesis and proliferation in cancer cells. Int. J. Cancer 2010, 126, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Iritani, N.; Noguchi, T. Transcriptional regulatory regions for expression of the rat fatty acid synthase. FEBS Lett. 1997, 406, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.D.; Charos, A.E.; Szekely, A.M.; Weissman, S.M.; Snyder, M. Genome-wide occupancy of SREBP1 and its partners NFY and SP1 reveals novel functional roles and combinatorial regulation of distinct classes of genes. PLoS Genet. 2008, 4, e1000133. [Google Scholar] [CrossRef] [PubMed]
- Alberts, A.W.; Vagelos, P.R. Acetyl CoA carboxylase. I. Requirement for two protein fractions. Proc. Natl. Acad. Sci. USA 1968, 59, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.A.; Kim, K.S.; Cho, U.H.; Yoon, D.J.; Park, S.W. Characterization of regulatory elements on the promoter region of human ATP-citrate lyase. Exp. Mol. Med. 1999, 31, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Daniel, S.; Kim, K.H. Sp1 mediates glucose activation of the acetyl-CoA carboxylase promoter. J. Biol. Chem. 1996, 271, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Krey, G.; Mahfoudi, A.; Wahli, W. Functional interactions of peroxisome proliferator-activated receptor, retinoid-X receptor, and Sp1 in the transcriptional regulation of the acyl-coenzyme-A oxidase promoter. Mol. Endocrinol. 1995, 9, 219–231. [Google Scholar] [PubMed]
- Schmitz, G.; Langmann, T. Transcriptional regulatory networks in lipid metabolism control ABCA1 expression. Biochim. Biophys. Acta 2005, 1735, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kaczynski, J.; Cook, T.; Urrutia, R. Sp1- and kruppel-like transcription factors. Genome Biol. 2003, 4, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Azumano, M.; Uwatoko, C.; Itoh, K.; Kuwahara, J. Role of zinc finger structure in nuclear localization of transcription factor Sp1. Biochem. Biophys. Res. Commun. 2009, 380, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Ferro, T.J. Sp1: Regulation of gene expression by phosphorylation. Gene 2005, 348, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Samson, S.L.; Wong, N.C. Role of Sp1 in insulin regulation of gene expression. J. Mol. Endocrinol. 2002, 29, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Han, I.; Kudlow, J.E. Reduced O glycosylation of Sp1 is associated with increased proteasome susceptibility. Mol. Cell. Biol. 1997, 17, 2550–2258. [Google Scholar] [PubMed]
- Wells, L.; Vosseller, K.; Hart, G.W. Glycosylation of nucleocytoplasmic proteins: Signal transduction and O-GlcNAc. Science 2001, 291, 2376–2378. [Google Scholar] [CrossRef]
- Roos, M.D.; Su, K.; Baker, J.R.; Kudlow, J.E. O glycosylation of an Sp1-derived peptide blocks known Sp1 protein interactions. Mol. Cell. Biol. 1997, 17, 6472–6480. [Google Scholar] [PubMed]
- Bionaz, M.; Loor, J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genomics 2008, 9, 366. [Google Scholar] [CrossRef]
- Degrelle, S.A.; Murthi, P.; Evain-Brion, D.; Fournier, T.; Hue, I. Expression and localization of DLX3, PPARG and Sp1 in bovine trophoblast during binucleated cell differentiation. Placenta 2011, 32, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Sassa, Y.; Hata, Y.; Aiello, L.P.; Taniguchi, Y.; Kohno, K.; Ishibashi, T. Bifunctional properties of peroxisome proliferator-activated receptor γ1 in KDR gene regulation mediated via interaction with both Sp1 and Sp3. Diabetes 2004, 53, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Yellaturu, C.; Cagen, L.; Wilcox, H.G.; Park, E.A.; Raghow, R.; Elam, M.B. Expression of the rat sterol regulatory element-binding protein-1c gene in response to insulin is mediated by increased transactivating capacity of specificity protein 1 (Sp1). J. Biol. Chem. 2007, 282, 17517–17529. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, L.; Roy, R.; Pampin, S.; Zaragoza, P.; Osta, R.; Rodriguez-Rey, J.C.; Rodellar, C. The g.763G>C SNP of the bovine FASN gene affects its promoter activity via Sp-mediated regulation: Implications for the bovine lactating mammary gland. Physiol. Genomics 2008, 34, 144–148. [Google Scholar] [CrossRef] [PubMed]
- McFadden, J.W.; Corl, B.A. Activation of liver X receptor (LXR) enhances de novo fatty acid synthesis in bovine mammary epithelial cells. J. Dairy Sci. 2010, 93, 4651–4658. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Seyfert, H.M. Promoter II of the bovine acetyl-coenzyme A carboxylase-α-encoding gene is widely expressed and strongly active in different cells. Biochim. Biophys. Acta 2002, 1576, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Yang, J.; Horton, J.D.; Hammer, R.E.; Goldstein, J.L.; Brown, M.S. Diminished hepatic response to fasting/refeeding and liver X receptor agonists in mice with selective deficiency of sterol regulatory element-binding protein-1c. J. Biol. Chem. 2002, 277, 9520–9528. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.S.; Hu, S.L.; Yu, K.; Wang, H.; Wang, W.; Loor, J.; Luo, J. Lipoprotein lipase, tissue expression and effects on genes related to fatty acid synthesis in goat mammary epithelial cells. Int. J. Mol. Sci. 2014, 15, 22757–22771. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lan, X.; Guo, W.; Sun, J.; Huang, Y.; Wang, J.; Huang, T.; Lei, C.; Fang, X.; Chen, H. Comparative transcriptome profiling of dairy goat micrornas from dry period and peak lactation mammary gland tissues. PLoS One 2012, 7, e52388. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Gu, Z.; Heid, B.; Akers, R.M.; Jiang, H. Identification and characterization of the bovine G protein-coupled receptor GPR41 and GPR43 genes. J. Dairy Sci. 2009, 92, 2696–2705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.D.; Liu, X.; Li, Y.S.; Ding, J.P.; Zhang, X.R.; Zhang, Y.H. Reference gene screening for analyzing gene expression across goat tissue. Asian-Australas. J. Anim. Sci. 2013, 26, 1665–1671. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-Z.; Luo, J.; Zhang, L.-P.; Wang, W.; Shi, H.-B.; Zhu, J.-J. miR-27a suppresses triglyceride accumulation and affects gene mRNA expression associated with fat metabolism in dairy goat mammary gland epithelial cells. Gene 2013, 521, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luo, J.; Wang, H.; Shi, H.; Zhu, J.; Sun, Y.; Yu, K.; Yao, D. Adipose triglyceride lipase regulates lipid metabolism in dairy goat mammary epithelial cells. Gene 2015, 554, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.J.; Luo, J.; Wang, W.; Yu, K.; Wang, H.B.; Shi, H.B.; Sun, Y.T.; Lin, X.Z.; Li, J. Inhibition of FASN reduces the synthesis of medium-chain fatty acids in goat mammary gland. Animal 2014, 8, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Kadegowda, A.K.; Bionaz, M.; Thering, B.; Piperova, L.S.; Erdman, R.A.; Loor, J.J. Identification of internal control genes for quantitative polymerase chain reaction in mammary tissue of lactating cows receiving lipid supplements. J. Dairy Sci. 2009, 92, 2007–2019. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.B.; Luo, J.; Yao, D.W.; Zhu, J.J.; Xu, H.F.; Shi, H.P.; Loor, J.J. Peroxisome proliferator-activated receptor-γ stimulates the synthesis of monounsaturated fatty acids in dairy goat mammary epithelial cells via the control of stearoyl-coenzyme A desaturase. J. Dairy Sci. 2013, 96, 7844–7853. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Shi, H.; Luo, J.; Wang, W.; Haile, A.B.; Xu, H.; Li, J. Establishment and characterization of a dairy goat mammary epithelial cell line with human telomerase (ht-MECs). Anim. Sci. J. 2014, 85, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Luo, J.; Zhong, Y.; Lin, X.-Z.; Shi, H.-B.; Zhu, J.-J.; Li, J.; Sun, Y.-T.; Zhao, W.-S. Goat liver X receptor α, molecular cloning, functional characterization and regulating fatty acid synthesis in epithelial cells of goat mammary glands. Gene 2012, 505, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.G.; Matitashvili, E.A.; Bauman, D.E. The inhibitory effect of trans-10, cis-12 CLA on lipid synthesis in bovine mammary epithelial cells involves reduced proteolytic activation of the transcription factor SREBP-1. J. Nutr. 2004, 134, 2523–2527. [Google Scholar] [PubMed]
- Kang, Y.; Hengbo, S.; Jun, L.; Jun, L.; Wangsheng, Z.; Huibin, T.; Huaiping, S. PPARG modulated lipid accumulation in dairy gmec via regulation of ADRP gene. J. Cell. Biochem. 2015, 116, 192–201. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Sun, Y.; Luo, J.; Wu, M.; Li, J.; Cao, Y. Specificity Protein 1 Regulates Gene Expression Related to Fatty Acid Metabolism in Goat Mammary Epithelial Cells. Int. J. Mol. Sci. 2015, 16, 1806-1820. https://doi.org/10.3390/ijms16011806
Zhu J, Sun Y, Luo J, Wu M, Li J, Cao Y. Specificity Protein 1 Regulates Gene Expression Related to Fatty Acid Metabolism in Goat Mammary Epithelial Cells. International Journal of Molecular Sciences. 2015; 16(1):1806-1820. https://doi.org/10.3390/ijms16011806
Chicago/Turabian StyleZhu, Jiangjiang, Yuting Sun, Jun Luo, Min Wu, Jianhua Li, and Yanhong Cao. 2015. "Specificity Protein 1 Regulates Gene Expression Related to Fatty Acid Metabolism in Goat Mammary Epithelial Cells" International Journal of Molecular Sciences 16, no. 1: 1806-1820. https://doi.org/10.3390/ijms16011806
APA StyleZhu, J., Sun, Y., Luo, J., Wu, M., Li, J., & Cao, Y. (2015). Specificity Protein 1 Regulates Gene Expression Related to Fatty Acid Metabolism in Goat Mammary Epithelial Cells. International Journal of Molecular Sciences, 16(1), 1806-1820. https://doi.org/10.3390/ijms16011806