
Connexin 43 Suppresses Tumor Angiogenesis by Down-Regulation of Vascular Endothelial Growth Factor via Hypoxic-Induced Factor-1α

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
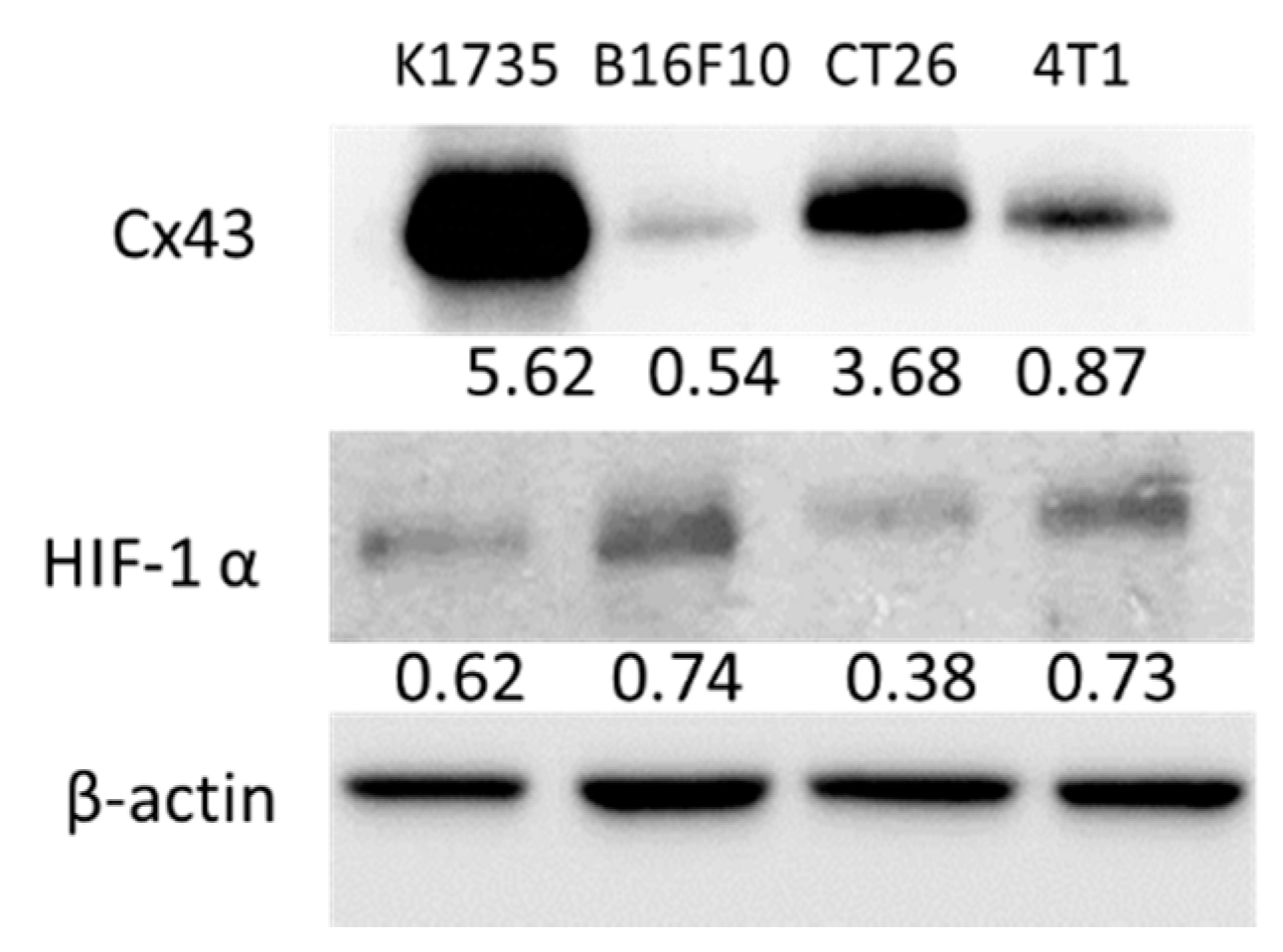
2.1. Murine Tumor Cells Express Cx43 and HIF-1α

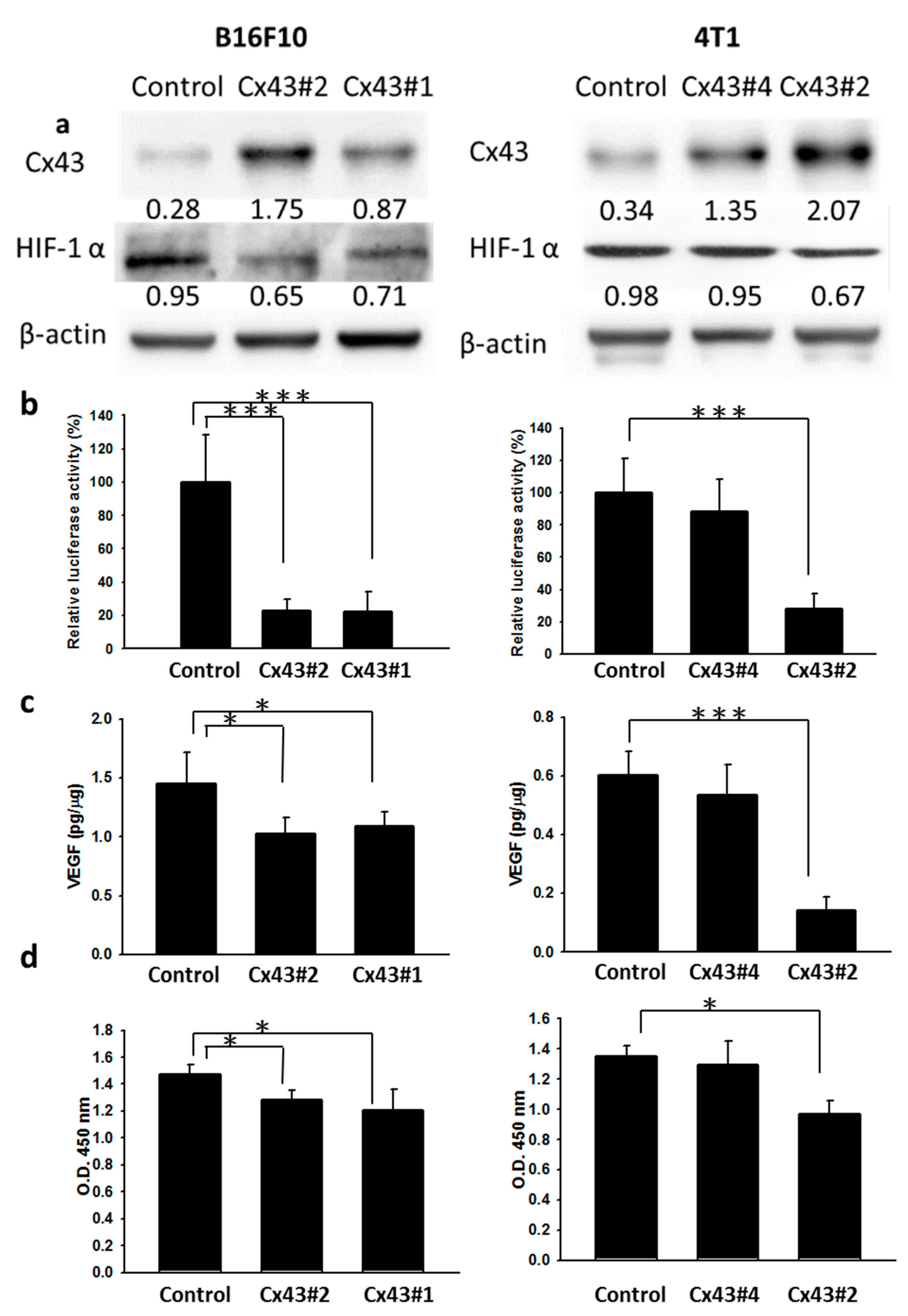
2.2. Cx43 Regulates the Expression of HIF-1α

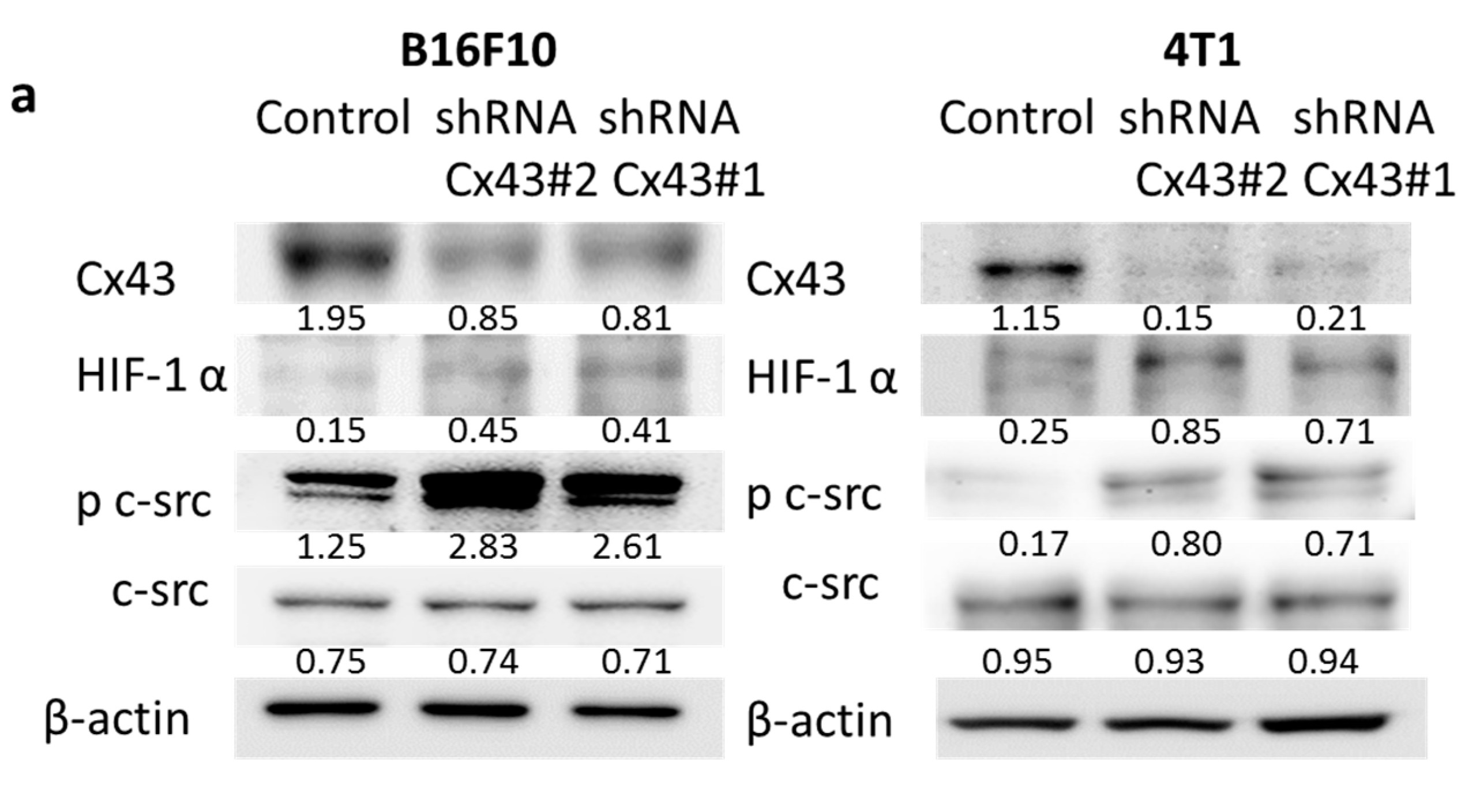
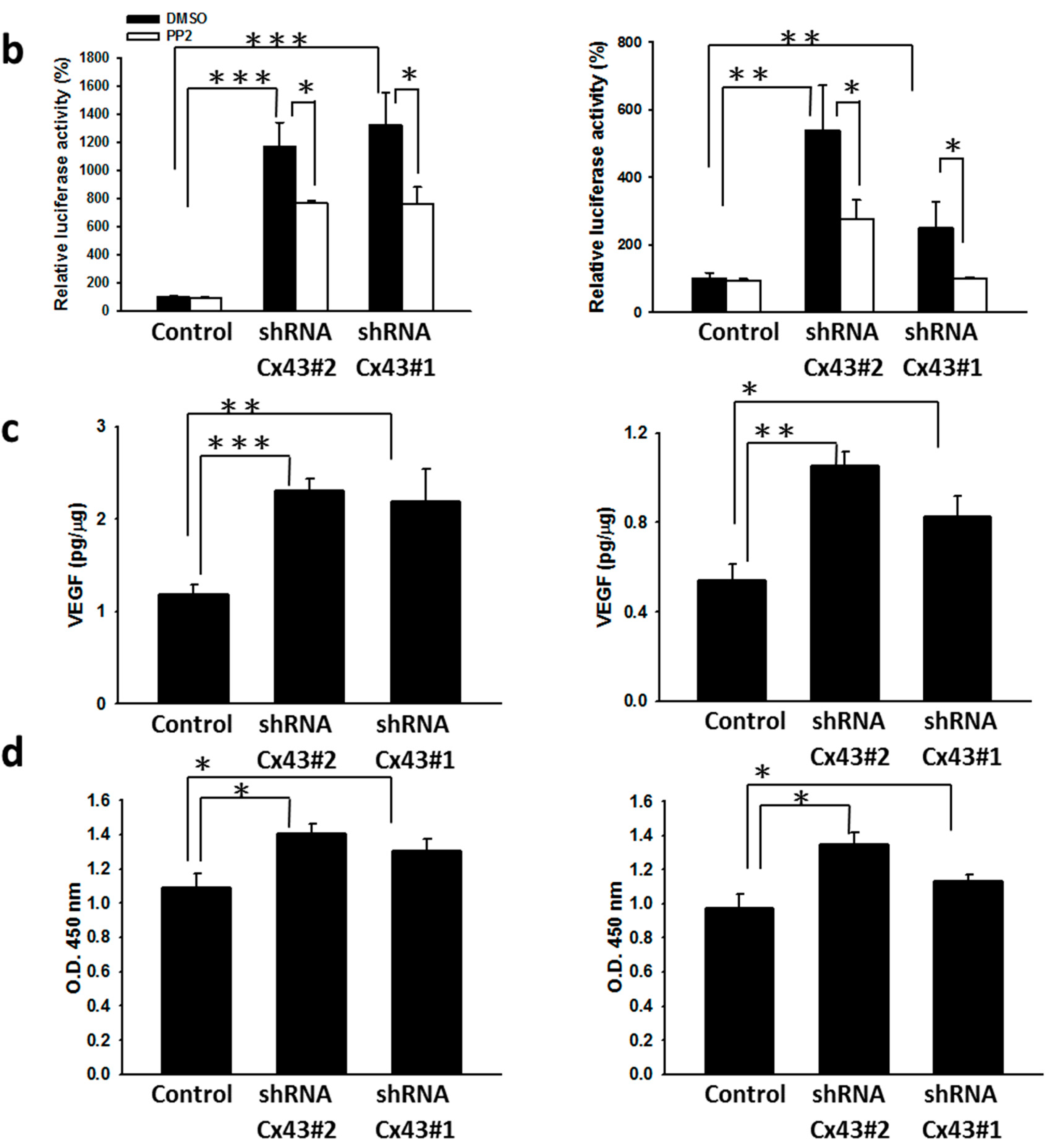
2.3. Reduction of Cx43 Expression by Cx43: Short Hairpin RNA (shRNA) Tumor Cells Are More Hypoxia-Responsive by Increasing the Transcriptional Activity and Stability of HIF-1α Protein


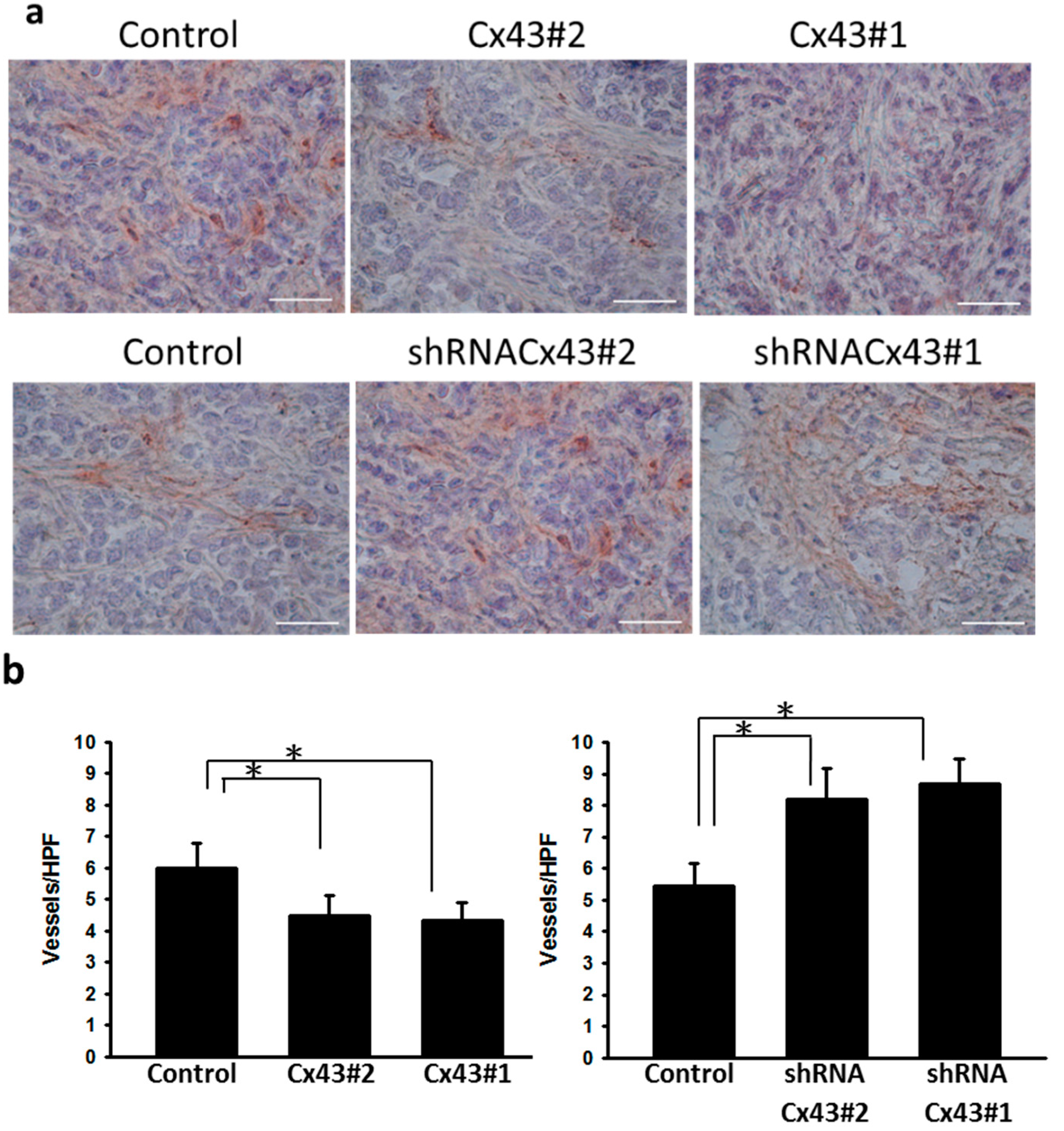
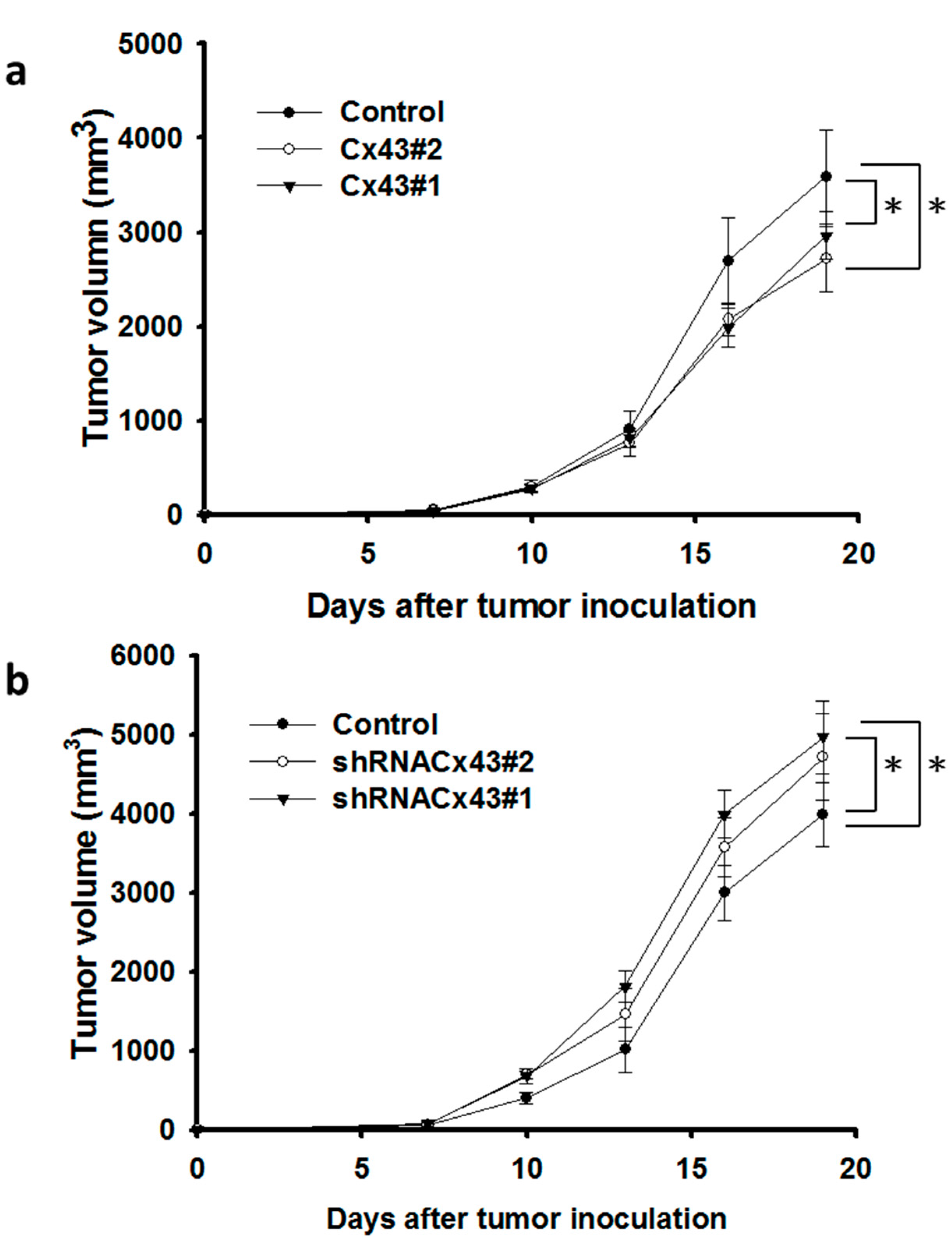
2.4. Inhibition of Tumor Angiogenesis in Cx43-Overexpressing Tumor Cells


3. Discussion
4. Experimental Section
4.1. Cell Lines, Reagents, Mice and Plasmid
4.2. Knockdown of Cx43 and Cx43 Overexpression
4.3. Analysis of Hypoxia-Inducible Transcriptional Activities
4.4. Immunoblot Analysis and Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Assay of Endothelial Proliferation
4.6. Immunohistochemistry
4.7. Animal Studies
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sharrow, A.C.; Li, Y.; Micsenyi, A.; Griswold, R.D.; Wells, A.; Monga, S.S.; Blair, H.C. Modulation of osteoblast gap junction connectivity by serum, TNF-α, and TRAIL. Exp. Cell Res. 2008, 314, 297–308. [Google Scholar] [PubMed]
- Wang, M.; Berthoud, V.M.; Beyer, E.C. Connexin 43 increases the sensitivity of prostate cancer cells to TNF-α-induced apoptosis. J. Cell Sci. 2007, 120, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Fukushima, M.; Maitani, Y. Non-viral delivery of the connexin 43 gene with histone deacetylase inhibitor to human nasopharyngeal tumor cells enhances gene expression and inhibits in vivo tumor growth. Int. J. Oncol. 2007, 30, 1427–1439. [Google Scholar] [PubMed]
- Huang, R.; Lin, Y.; Wang, C.C.; Gano, J.; Lin, B.; Shi, Q.; Boynton, A.; Burke, J.; Huang, R.P. Connexin 43 suppresses human glioblastoma cell growth by down-regulation of monocyte chemotactic protein 1, as discovered using protein array technology. Cancer Res. 2002, 62, 2806–2812. [Google Scholar] [PubMed]
- Ionta, M.; Ferreira, R.A.; Pfister, S.C.; Machado-Santelli, G.M. Exogenous Cx43 expression decrease cell proliferation rate in rat hepatocarcinoma cells independently of functional gap junction. Cancer Cell Int. 2009, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.W.; Lai, C.H.; Chen, M.C.; Liu, C.F.; Kuan, Y.D.; Lin, S.T.; Lee, C.H. Salmonella enhance chemosensitivity in tumor through connexin 43 upregulation. Int. J. Cancer 2013, 133, 1926–1935. [Google Scholar] [CrossRef] [PubMed]
- Ableser, M.J.; Penuela, S.; Lee, J.; Shao, Q.; Laird, D.W. Connexin43 reduces melanoma growth within a keratinocyte microenvironment and during tumorigenesis in vivo. J. Biol. Chem. 2014, 289, 1592–1603. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Hypoxia-induced cytosine deaminase gene expression for cancer therapy. Hum. Gene Ther. 2007, 18, 27–38. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, E.; Shao, Q.; Wang, H.L.; Langlois, S.; Laird, D.W. Connexins act as tumor suppressors in three-dimensional mammary cell organoids by regulating differentiation and angiogenesis. Cancer Res. 2006, 66, 9886–9894. [Google Scholar] [CrossRef]
- Neitzel, L.T.; Neitzel, C.D.; Magee, K.L.; Malafa, M.P. Angiogenesis correlates with metastasis in melanoma. Ann. Surg. Oncol. 1999, 6, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Quintieri, L.; Selmy, M.; Indraccolo, S. Metabolic effects of antiangiogenic drugs in tumors: Therapeutic implications. Biochem. Pharmacol. 2014, 89, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Systemic administration of attenuated Salmonella choleraesuis carrying thrombospondin-1 gene leads to tumor-specific transgene expression, delayed tumor growth and prolonged survival in the murine melanoma model. Cancer Gene Ther. 2005, 12, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Endostatin gene therapy delivered by Salmonella choleraesuis in murine tumor models. J. Gene Med. 2004, 6, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.W.; Kuan, Y.D.; Chen, M.C.; Lin, S.T.; Lee, C.H. Tracking of mouse breast cancer stem-like cells with Salmonella. Exp. Biol. Med. 2012, 237, 1189–1196. [Google Scholar] [CrossRef]
- Shan, Y.S.; Hsu, H.P.; Lai, M.D.; Yen, M.C.; Fang, J.H.; Weng, T.Y.; Chen, Y.L. Suppression of mucin 2 promotes interleukin-6 secretion and tumor growth in an orthotopic immune-competent colon cancer animal model. Oncol. Rep. 2014, 32, 2335–2342. [Google Scholar] [PubMed]
- Valle-Casuso, J.C.; González-Sánchez, A.; Medina, J.M.; Tabernero, A. HIF-1 and c-Src mediate increased glucose uptake induced by endothelin-1 and connexin43 in astrocytes. PLoS One 2012, 7, e32448. [Google Scholar] [CrossRef] [PubMed]
- Shiau, A.L.; Shen, Y.T.; Hsieh, J.L.; Wu, C.L.; Lee, C.H. Scutellaria barbata inhibits angiogenesis through downregulation of HIF-1α in lung tumor. Environ. Toxicol. 2014, 29, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Laderoute, K.R.; Alarcon, R.M.; Brody, M.D.; Calaoagan, J.M.; Chen, E.Y.; Knapp, A.M.; Yun, Z.; Denko, N.C.; Giaccia, A.J. Opposing effects of hypoxia on expression of the angiogenic inhibitor thrombospondin 1 and the angiogenic inducer vascular endothelial growth factor. Clin. Cancer Res. 2000, 6, 2941–2950. [Google Scholar] [PubMed]
- Salnikow, K.; An, W.G.; Melillo, G.; Blagosklonny, M.V.; Costa, M. Nickel-induced transformation shifts the balance between HIF-1 and p53 transcription factors. Carcinogenesis 1999, 20, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; de Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor-1α in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar] [PubMed]
- Zhang, Y.W.; Kaneda, M.; Morita, I. The gap junction-independent tumor-suppressing effect of connexin 43. J. Biol. Chem. 2003, 278, 44852–44856. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.X.; Gu, S. Gap junction and hemichannel-independent actions of connexins. Biochim. Biophys. Acta 2005, 1711, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Giepmans, B.N.; Hengeveld, T.; Postma, F.R.; Moolenaar, W.H. Interaction of c-Src with gap junction protein connexin-43: Role in the regulation of cell-cell communication. J. Biol. Chem. 2001, 276, 8544–8549. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.J.; Chang, M.Y.; Chang, W.W.; Wang, W.K.; Liu, C.F.; Lin, S.T.; Lee, C.H. Resveratrol enhances chemosensitivity in mouse melanoma model through connexin 43 upregulation. Environ. Toxicol. 2014. [CrossRef]
- Wang, W.K.; Kuan, Y.D.; Kuo, C.Y.; Lee, C.H. Connexin 43 gene therapy delivered by polymer-modified Salmonella in murine tumor models. Polymers 2014, 6, 1119–1128. [Google Scholar] [CrossRef]
- Lee, C.H.; Lin, S.T.; Liu, J.J.; Chang, W.W.; Hsieh, J.L.; Wang, W.K. Salmonella induce autophagy in melanoma by the downregulation of AKT/mTOR pathway. Gene Ther. 2014, 21, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, S.D.; Ou, H.C.; Lai, S.C.; Cheng, Y.J. Eicosapentaenoic acid protects against palmitic acid-induced endothelial dysfunction via activation of the AMPK/eNOS pathway. Int. J. Mol. Sci. 2014, 15, 10334–10349. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.W.; Liu, J.J.; Liu, C.F.; Liu, W.S.; Lim, Y.P.; Cheng, Y.J.; Lee, C.H. An extract of Rhodobacter sphaeroides reduces cisplatin-induced nephrotoxicity in mice. Toxins 2013, 5, 2353–2365. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.W.; Kuan, Y.D.; Chiu, K.H.; Wang, W.K.; Chang, F.H.; Liu, C.H.; Lee, C.H. The extract of Rhodobacter sphaeroides inhibits melanogenesis through the MEK/ERK signaling pathway. Mar. Drug 2013, 11, 1899–1908. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.-K.; Chen, M.-C.; Leong, H.-F.; Kuo, Y.-L.; Kuo, C.-Y.; Lee, C.-H. Connexin 43 Suppresses Tumor Angiogenesis by Down-Regulation of Vascular Endothelial Growth Factor via Hypoxic-Induced Factor-1α. Int. J. Mol. Sci. 2015, 16, 439-451. https://doi.org/10.3390/ijms16010439
Wang W-K, Chen M-C, Leong H-F, Kuo Y-L, Kuo C-Y, Lee C-H. Connexin 43 Suppresses Tumor Angiogenesis by Down-Regulation of Vascular Endothelial Growth Factor via Hypoxic-Induced Factor-1α. International Journal of Molecular Sciences. 2015; 16(1):439-451. https://doi.org/10.3390/ijms16010439
Chicago/Turabian StyleWang, Wei-Kuang, Man-Chin Chen, Hon-Fai Leong, Yu-Liang Kuo, Chun-Yu Kuo, and Che-Hsin Lee. 2015. "Connexin 43 Suppresses Tumor Angiogenesis by Down-Regulation of Vascular Endothelial Growth Factor via Hypoxic-Induced Factor-1α" International Journal of Molecular Sciences 16, no. 1: 439-451. https://doi.org/10.3390/ijms16010439
APA StyleWang, W.-K., Chen, M.-C., Leong, H.-F., Kuo, Y.-L., Kuo, C.-Y., & Lee, C.-H. (2015). Connexin 43 Suppresses Tumor Angiogenesis by Down-Regulation of Vascular Endothelial Growth Factor via Hypoxic-Induced Factor-1α. International Journal of Molecular Sciences, 16(1), 439-451. https://doi.org/10.3390/ijms16010439