
Improved Monitoring of Semi-Continuous Anaerobic Digestion of Sugarcane Waste: Effects of Increasing Organic Loading Rate on Methanogenic Community Dynamics


Abstract
:1. Introduction
2. Results and Discussion
2.1. Biogas Reactor Performance
2.2. Methanogenic Community Dynamics

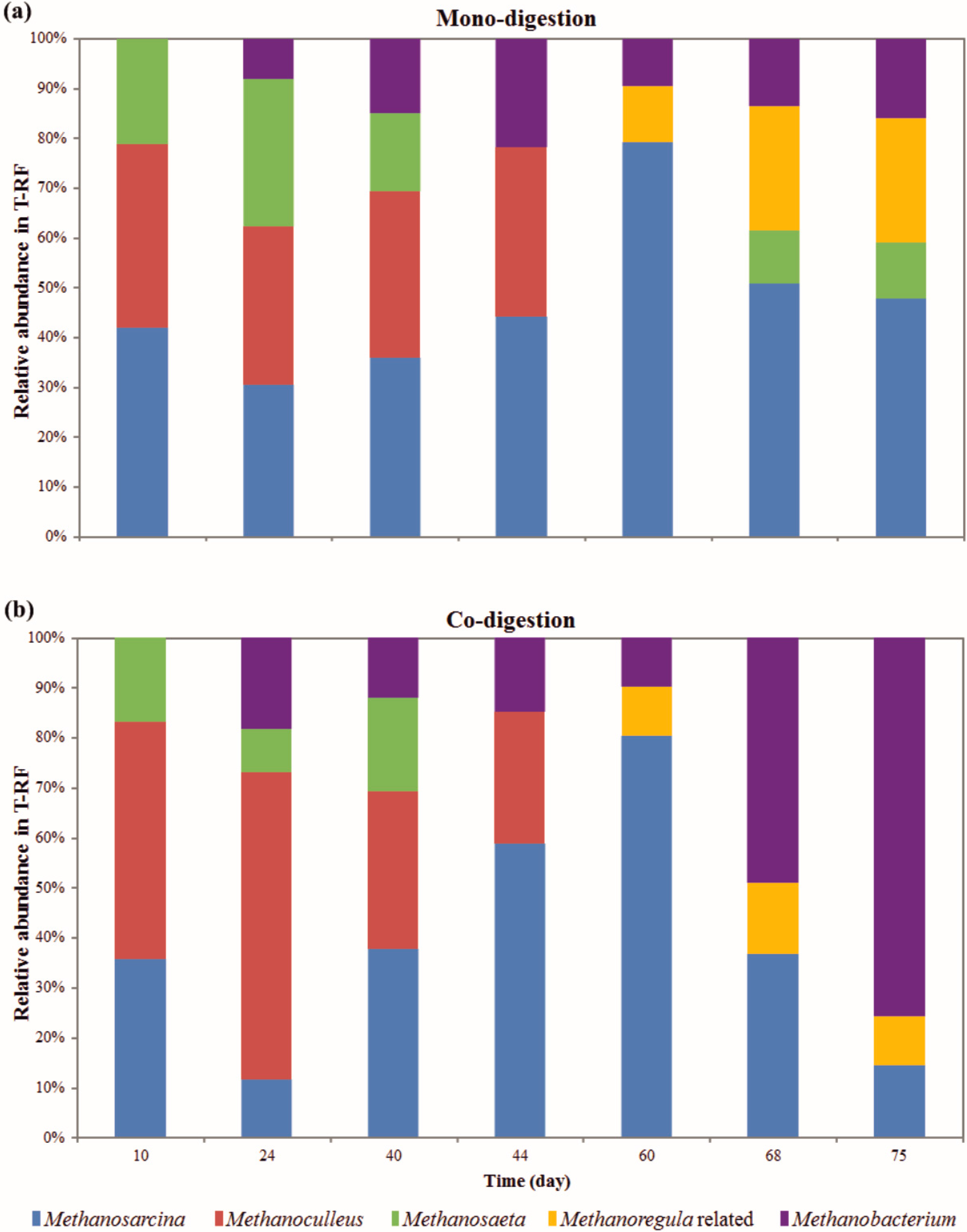{kind=link}
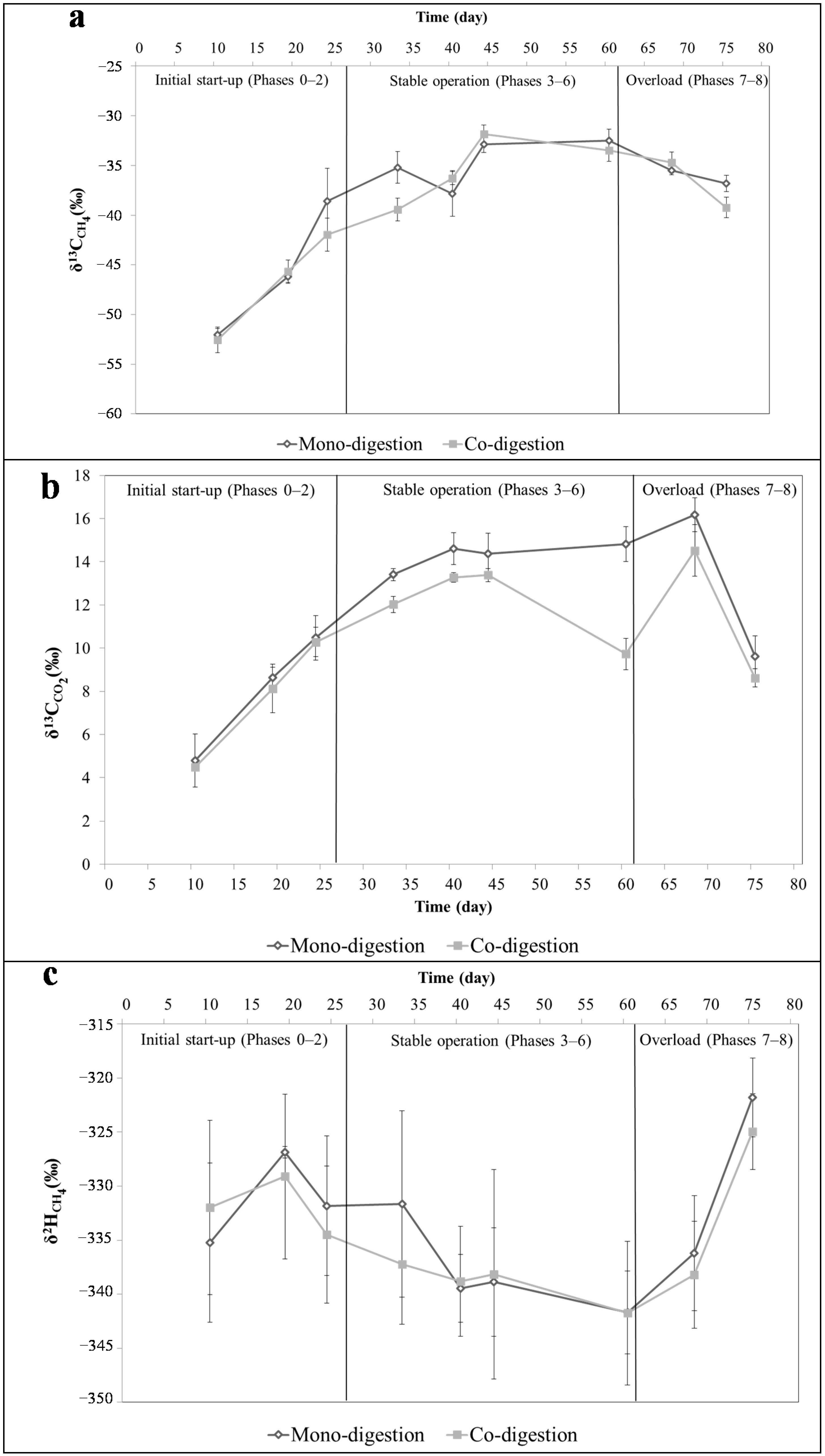{kind=link}
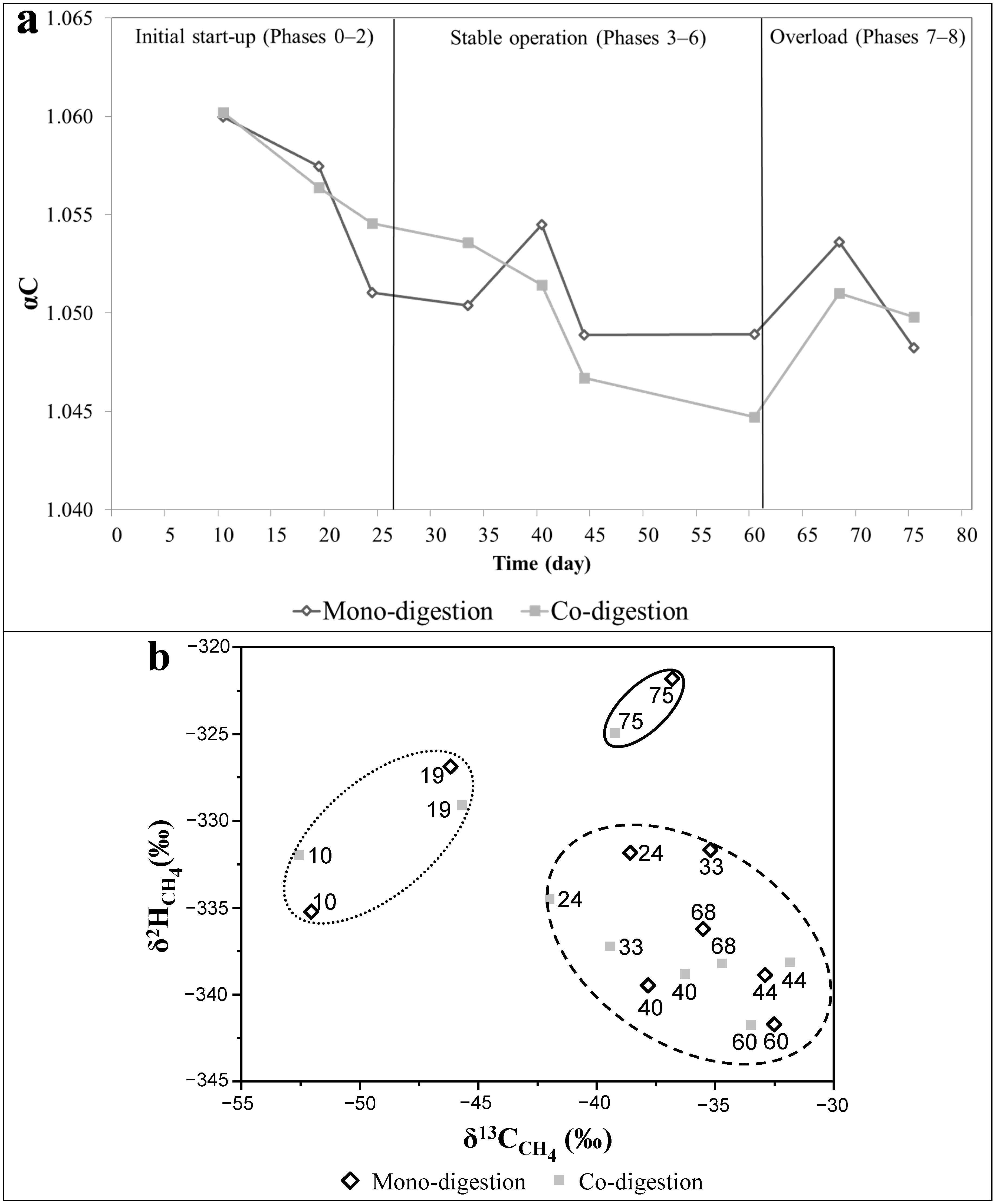{kind=link}
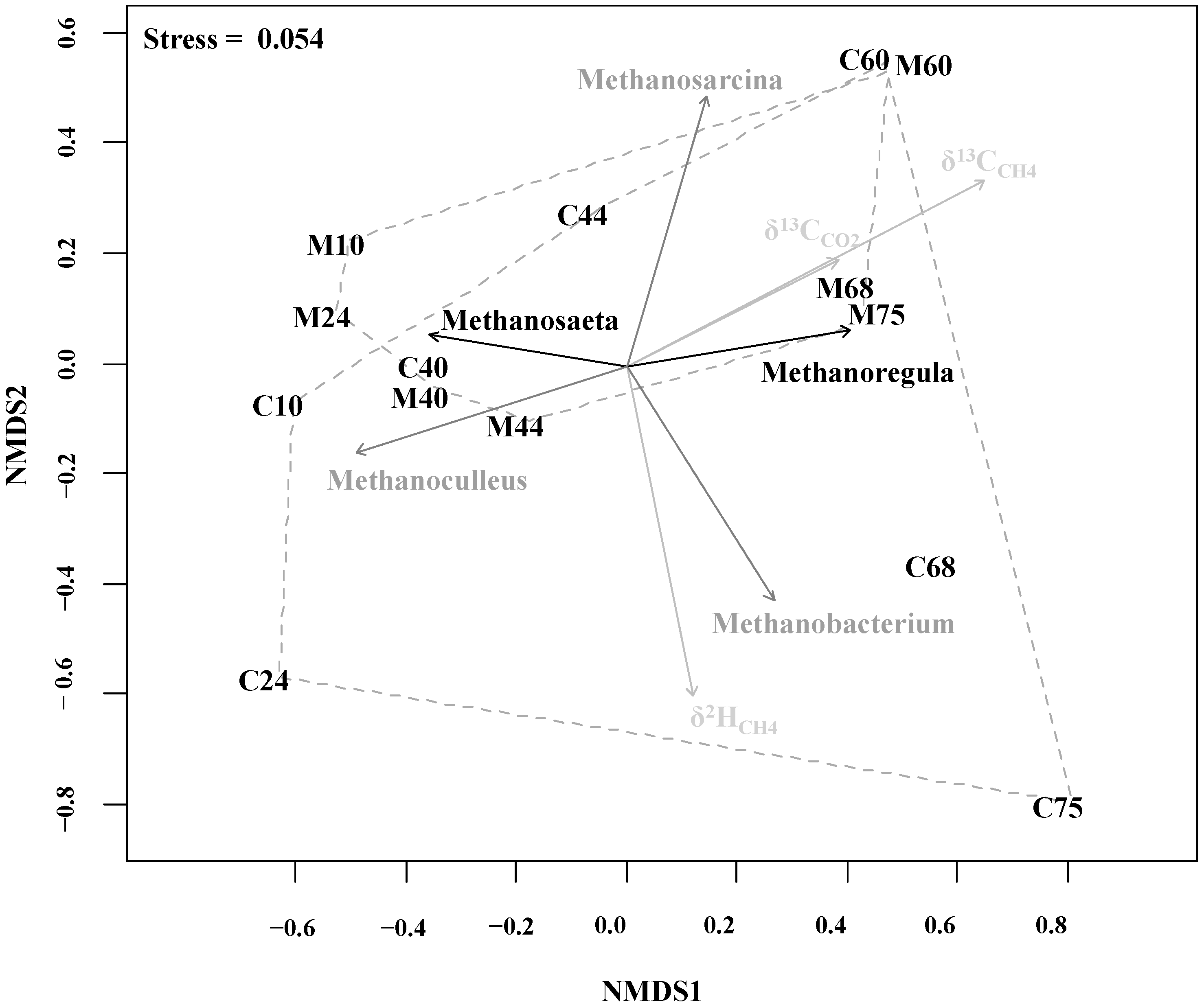{kind=link}
| Reactor’ Parameters | Phase 1 (sampling Day: 19) | Phase 2 (sampling day: 26) | Phase 3 (sampling day: 33) | Phase 4 (sampling day: 40) | Phase 5 (sampling day: 55) | Phase 6 (sampling day: 61) | Phase 7 (sampling day: 68) | Phase 8 (sampling day: 75) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | |
| Digestion | Digestion | Digestion | Digestion | Digestion | Digestion | Digestion | Digestion | |||||||||
| Biogas yield * (mL·gVS−1) | 1086.3 | 1198.2 | 506.9 | 541.0 | 283.9 | 409.6 | 292.8 | 329.2 | 368.3 | 410.9 | 397.6 | 251.2 | 127.9 | 58.3 | 69.7 | 33.7 |
| CH4§ (%) | 57.7 | 55.6 | 57.4 | 56.6 | 60.3 | 53.9 | 57.8 | 54.9 | 61.3 | 54.9 | 61.1 | na | na | na | na | na |
| CO2§ (%) | 42.3 | 44.4 | 42.6 | 43.4 | 39.7 | 46.1 | 42.2 | 45.1 | 38.7 | 45.1 | 38.9 | na | na | na | na | na |
| Acetic acid (mg·L−1) | 55.7 | 37.3 | 26.7 | 25.4 | 46.1 | 69.6 | 160.3 | 120.5 | 240.6 | 155.6 | 145.1 | 141.9 | 1003.0 | 914.2 | 1370.0 | 1334.1 |
| Propionic acid (mg·L−1) | 10.7 | 6.9 | 6.7 | 4.9 | 5.3 | 4.6 | 13.1 | 8.8 | 10.4 | 5.8 | 208.0 | 283.7 | 537.6 | 550.7 | 433.9 | 391.8 |
| n-Butyric acid (mg·L−1) | 4.7 | 2.5 | 3.1 | 2.0 | 2.2 | 1.8 | 19.5 | 12.1 | 5.3 | 1.8 | 26.0 | 6.7 | 428.9 | 403.3 | 1144.8 | 1193.0 |
| VOA (g·L−1) | 0.8 | 0.8 | 0.6 | 0.7 | 0.6 | 0.6 | 0.5 | 0.5 | 1.0 | 0.8 | 0.6 | 0.6 | 2.2 | 2.1 | na | na |
| VOA/TIC (gVOA·gCaCO3−1) | 0.2 | 0.2 | 0.2 | 0.1 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.7 | 0.6 | 3.1 | 3.1 | na | na |
| pH * | 7.5 | 7.5 | 7.4 | 7.3 | 7.5 | 7.5 | 7.2 | 7.1 | 7.2 | 6.9 | 7.0 | 6.5 | 6.3 | 5.7 | 5.4 | 5.2 |
| NH4-N (g·L−1) | 1.0 | 1.1 | 0.8 | 0.8 | 0.6 | 0.9 | 0.5 | 0.5 | 0.3 | 0.3 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.2 |
| TS (%) | 3.1 | 2.9 | 2.7 | 2.6 | 2.5 | 2.7 | 2.3 | 2.8 | 2.1 | 2.3 | 2.1 | 2.1 | 5.6 | na | 6.9 | 7.6 |
| VS (%) | 2.1 | 2.0 | 1.8 | 1.7 | 1.7 | 1.8 | 1.6 | 1.9 | 1.5 | 1.7 | 1.6 | 1.5 | 3.8 | na | 4.5 | 5.0 |

2.3. Isotopic Changes of the Produced Biogas

2.4. Methanogenic Pathways


3. Experimental Section
3.1. Biogas Reactors, Operation and Analytical Methods
3.2. Methanogenic Community Analysis
| Set-up-Technical Parameters | Phase 1 | Phase 2 | Phase 3 | Phase 4 | Phase 5 | Phase 6 | Phase 7 | Phase 8 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | Mono- | Co- | |
| Digestion | Digestion | Digestion | Digestion | Digestion | Digestion | Digestion | Digestion | |||||||||
| Experiment phase (day) | 11–19 | 20–26 | 27–33 | 34–40 | 41–55 | 56–61 | 62–68 | 69–75 | ||||||||
| Substrate (g fresh mass) | 2.4 | 1.5 | 4.9 | 3.0 | 7.3 | 4.5 | 9.7 | 6.0 | 12.2 | 7.5 | 14.6 | 9.0 | 10.7 | 10.5 | 12.2 | 12.0 |
| Water mixed with substrate (mL) | 20 | 25 | 30 | 35 | 45 | 45 | 50 | 55 | ||||||||
| Working volume (L) | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.5 | 0.8 | 0.5 | 0.8 | ||||||
| VS (g·day−1) | 0.4 | 0.8 | 1.2 | 1.6 | 2.0 | 2.4 | 1.8 | 2.8 | 2.0 | 3.2 | ||||||
| OLR (gVS·L−1·day−1) | 0.5 | 1.0 | 1.5 | 2.0 | 2.5 | 3.0 | 3.5 | 4.0 | ||||||||
| HRT (day) | 35.7 | 37.2 | 26.8 | 28.6 | 21.4 | 23.2 | 17.9 | 19.5 | 15.3 | 15.2 | 13.4 | 14.8 | 8.2 | 13.2 | 7.4 | 11.9 |
3.3. Stable Isotope Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Leite, A.F.; Janke, L.; Harms, H.; Zang, J.W.; Fonseca-Zang, W.A.; Stinner, W.; Nikolausz, M. Assessment of the variations in characteristics and methane potential of major waste products from the brazilian bioethanol industry along an operating season. Energy Fuels 2015, 29, 4022–4029. [Google Scholar] [CrossRef]
- Janke, L.; Leite, A.; Batista, K.; Weinrich, S.; Strauber, H.; Nikolausz, M.; Nelles, M.; Stinner, W. Optimization of hydrolysis and volatile fatty acids production from sugarcane filter cake: Effects of urea supplementation and sodium hydroxide pretreatment. Bioresour. Technol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Janke, L.; Leite, A.; Wedwitschka, H.; Schmidt, T.; Nikolausz, M.; Stinner, W. Biomethane production integrated to the brazilian sugarcane industry: The case study of são paulo state. In Proceedings of the 22nd European Biomass Conference and Exhibition, Hamburg, Germany, December 2014; pp. 1295–1299.
- Janke, L.; Leite, A.; Nikolausz, M.; Schmidt, T.; Liebetrau, J.; Nelles, M.; Stinner, W. Biogas production from sugarcane waste: Assessment on kinetic challenges for process designing. Int. J. Mol. Sci. 2015, 16, 20685–20703. [Google Scholar] [CrossRef]
- Nikolausz, M.; Walter, R.F.; Strauber, H.; Liebetrau, J.; Schmidt, T.; Kleinsteuber, S.; Bratfisch, F.; Gunther, U.; Richnow, H.H. Evaluation of stable isotope fingerprinting techniques for the assessment of the predominant methanogenic pathways in anaerobic digesters. Appl. Microbiol. Biotechnol. 2013, 97, 2251–2262. [Google Scholar] [CrossRef] [PubMed]
- Weiland, P. Biogas production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 85, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: A review. Rev. Environ. Sci. Biol. Technol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef] [PubMed]
- Briones, A.; Raskin, L. Diversity and dynamics of microbial communities in engineered environments and their implications for process stability. Curr. Opin. Biotechnol. 2003, 14, 270–276. [Google Scholar] [CrossRef]
- Karakashev, D.; Batstone, D.J.; Angelidaki, I. Influence of environmental conditions on methanogenic compositions in anaerobic biogas reactors. Appl. Environ. Microbiol. 2005, 71, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.; Kuchenbuch, A.; Fetzer, I.; Harms, H.; Kleinsteuber, S. Long-term monitoring reveals stable and remarkably similar microbial communities in parallel full-scale biogas reactors digesting energy crops. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Blume, F.; Bergmann, I.; Nettmann, E.; Schelle, H.; Rehde, G.; Mundt, K.; Klocke, M. Methanogenic population dynamics during semi-continuous biogas fermentation and acidification by overloading. J. Appl. Microbiol. 2010, 109, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Franke-Whittle, I.H.; Walter, A.; Ebner, C.; Insam, H. Investigation into the effect of high concentrations of volatile fatty acids in anaerobic digestion on methanogenic communities. Waste Manag. 2014, 34, 2080–2089. [Google Scholar] [CrossRef] [PubMed]
- Marchaim, U.; Krause, C. Propionic to acetic acid ratios in overloaded anaerobic digestion. Bioresour. Technol. 1993, 43, 195–203. [Google Scholar] [CrossRef]
- Steinberg, L.M.; Regan, J.M. Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl. Environ. Microbiol. 2008, 74, 6663–6671. [Google Scholar] [CrossRef] [PubMed]
- Talbot, G.; Topp, E.; Palin, M.; Masse, D. Evaluation of molecular methods used for establishing the interactions and functions of microorganisms in anaerobic bioreactors. Water Res. 2008, 42, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Chidthaisong, A.; Chin, K.-J.; Valentine, D.L.; Tyler, S.C. A comparison of isotope fractionation of carbon and hydrogen from paddy field rice roots and soil bacterial enrichments during CO2/H2 methanogenesis. Geochim. Cosmochim. Acta 2002, 66, 983–995. [Google Scholar] [CrossRef]
- Lv, Z.; Leite, A.F.; Harms, H.; Richnow, H.H.; Liebetrau, J.; Nikolausz, M. Influences of the substrate feeding regime on methanogenic activity in biogas reactors approached by molecular and stable isotope methods. Anaerobe 2014, 29, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Hu, M.; Harms, H.; Richnow, H.H.; Liebetrau, J.; Nikolausz, M. Stable isotope composition of biogas allows early warning of complete process failure as a result of ammonia inhibition in anaerobic digesters. Bioresour. Technol. 2014, 167, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Keppler, F.; Laukenmann, S.; Rinne, J.; Heuwinkel, H.; Greule, M.; Whiticar, M.; Lelieveld, J. Measurements of 13C/12C methane from anaerobic digesters: Comparison of optical spectrometry with continuous-flow isotope ratio mass spectrometry. Environ. Sci. Technol. 2010, 44, 5067–5073. [Google Scholar] [CrossRef]
- Nielsen, H.; Uellendahl, H.; Ahring, B. Regulation and optimization of the biogas process: Propionate as a key parameter. Biomass Bioenerg. 2007, 31, 820–830. [Google Scholar] [CrossRef]
- Prochazka, J.; Dolejs, P.; Maca, J.; Dohanyos, M. Stability and inhibition of anaerobic processes caused by insufficiency or excess of ammonia nitrogen. Appl. Microbiol. Biotechnol. 2012, 93, 439–447. [Google Scholar] [CrossRef]
- Jenkins, D.; Richard, M.G.; Daigger, G.T. Manual on the Causes and Control of Activated Sludge Bulking, Foaming, and Other Solids Separation Problems; IWA Publishing: London, UK, 2003; pp. 131–161. [Google Scholar]
- Zinder, S. Physiological ecology of methanogens. In Methanogenesis; Ferry, J., Ed.; Springer: New York, NY, USA, 1993; pp. 128–206. [Google Scholar]
- Griffin, M.E.; McMahon, K.D.; Mackie, R.I.; Raskin, L. Methanogenic population dynamics during start-up of anaerobic digesters treating municipal solid waste and biosolids. Biotechnol. Bioeng. 1998, 57, 342–355. [Google Scholar] [CrossRef]
- Yu, Y.; Kim, J.; Hwang, S. Use of real-time PCR for group-specific quantification of aceticlastic methanogens in anaerobic processes: Population dynamics and community structures. Biotechnol. Bioeng. 2006, 93, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; He, Q. Persistence of methanosaeta populations in anaerobic digestion during process instability. J. Ind. Microbiol. Biotechnol. 2015, 42, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, Y.; Sakai, S.; Ehara, M.; Miyazaki, M.; Yamaguchi, T.; Imachi, H. Methanoregula formicica sp. nov., a methane-producing archaeon isolated from methanogenic sludge. Int. J. Syst. Evol. Microbiol. 2011, 61, 53–59. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, M.H. Carbon isotopes in photosysnthesis. BioScience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Conrad, R. Quantification of methanogenic pathways using stable carbon isotopic signatures: A review and a proposal. Org. Geochem. 2005, 36, 739–752. [Google Scholar] [CrossRef]
- Galand, P.E.; Yrjälä, K.; Conrad, R. Stable carbon isotope fractionation during methanogenesis in three boreal peatland ecosystems. Biogeosciences 2010, 7, 3893–3900. [Google Scholar] [CrossRef]
- Whiticar, M.J.; Faber, E.; Schoell, M. Biogenic methane formation in marine and freshwater environments: CO2 reduction vs. Acetate fermentation—Isotope evidence. Geochim. Cosmochim. Acta 1986, 50, 693–709. [Google Scholar] [CrossRef]
- Schmidt, T.; Ziganshin, A.M.; Nikolausz, M.; Scholwin, F.; Nelles, M.; Kleinsteuber, S.; Pröter, J. Effects of the reduction of the hydraulic retention time to 1.5 days at constant organic loading in CSTR, ASBR, and fixed-bed reactors—Performance and methanogenic community composition. Biomass Bioenerg. 2014, 69, 241–248. [Google Scholar] [CrossRef]
- Tauber, T.; Berta, B.; Szabo, Z.; Kovacs, J.; Marialigeti, K.; Toth, E.M. A simple and novel volumetric method to metre low gas flows from laboratory-scale bioreactors and its application on laboratory sludge digesters. Appl. Microbiol. Biotechnol. 2011, 90, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Ziganshin, A.M.; Schmidt, T.; Scholwin, F.; Il’inskaya, O.N.; Harms, H.; Kleinsteuber, S. Bacteria and archaea involved in anaerobic digestion of distillers grains with solubles. Appl. Microbiol. Biotechnol. 2011, 89, 2039–2052. [Google Scholar] [CrossRef] [PubMed]
- Sträuber, H.; Schröder, M.; Kleinsteuber, S. Metabolic and microbial community dynamics during the hydrolytic and acidogenic fermentation in a leach-bed process. Energy Sustain. Soc. 2012, 2, 13. [Google Scholar] [CrossRef]
- Feisthauer, S.; Siegert, M.; Seidel, M.; Richnow, H.H.; Zengler, K.; Gründger, F.; Krüger, M. Isotopic fingerprinting of methane and CO2 formation from aliphatic and aromatic hydrocarbons. Org. Geochem. 2010, 41, 482–490. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, A.F.; Janke, L.; Lv, Z.; Harms, H.; Richnow, H.-H.; Nikolausz, M. Improved Monitoring of Semi-Continuous Anaerobic Digestion of Sugarcane Waste: Effects of Increasing Organic Loading Rate on Methanogenic Community Dynamics. Int. J. Mol. Sci. 2015, 16, 23210-23226. https://doi.org/10.3390/ijms161023210
Leite AF, Janke L, Lv Z, Harms H, Richnow H-H, Nikolausz M. Improved Monitoring of Semi-Continuous Anaerobic Digestion of Sugarcane Waste: Effects of Increasing Organic Loading Rate on Methanogenic Community Dynamics. International Journal of Molecular Sciences. 2015; 16(10):23210-23226. https://doi.org/10.3390/ijms161023210
Chicago/Turabian StyleLeite, Athaydes Francisco, Leandro Janke, Zuopeng Lv, Hauke Harms, Hans-Hermann Richnow, and Marcell Nikolausz. 2015. "Improved Monitoring of Semi-Continuous Anaerobic Digestion of Sugarcane Waste: Effects of Increasing Organic Loading Rate on Methanogenic Community Dynamics" International Journal of Molecular Sciences 16, no. 10: 23210-23226. https://doi.org/10.3390/ijms161023210
APA StyleLeite, A. F., Janke, L., Lv, Z., Harms, H., Richnow, H. -H., & Nikolausz, M. (2015). Improved Monitoring of Semi-Continuous Anaerobic Digestion of Sugarcane Waste: Effects of Increasing Organic Loading Rate on Methanogenic Community Dynamics. International Journal of Molecular Sciences, 16(10), 23210-23226. https://doi.org/10.3390/ijms161023210