
Cancer Microenvironment: What Can We Learn from the Stem Cell Niche

 ,
, 

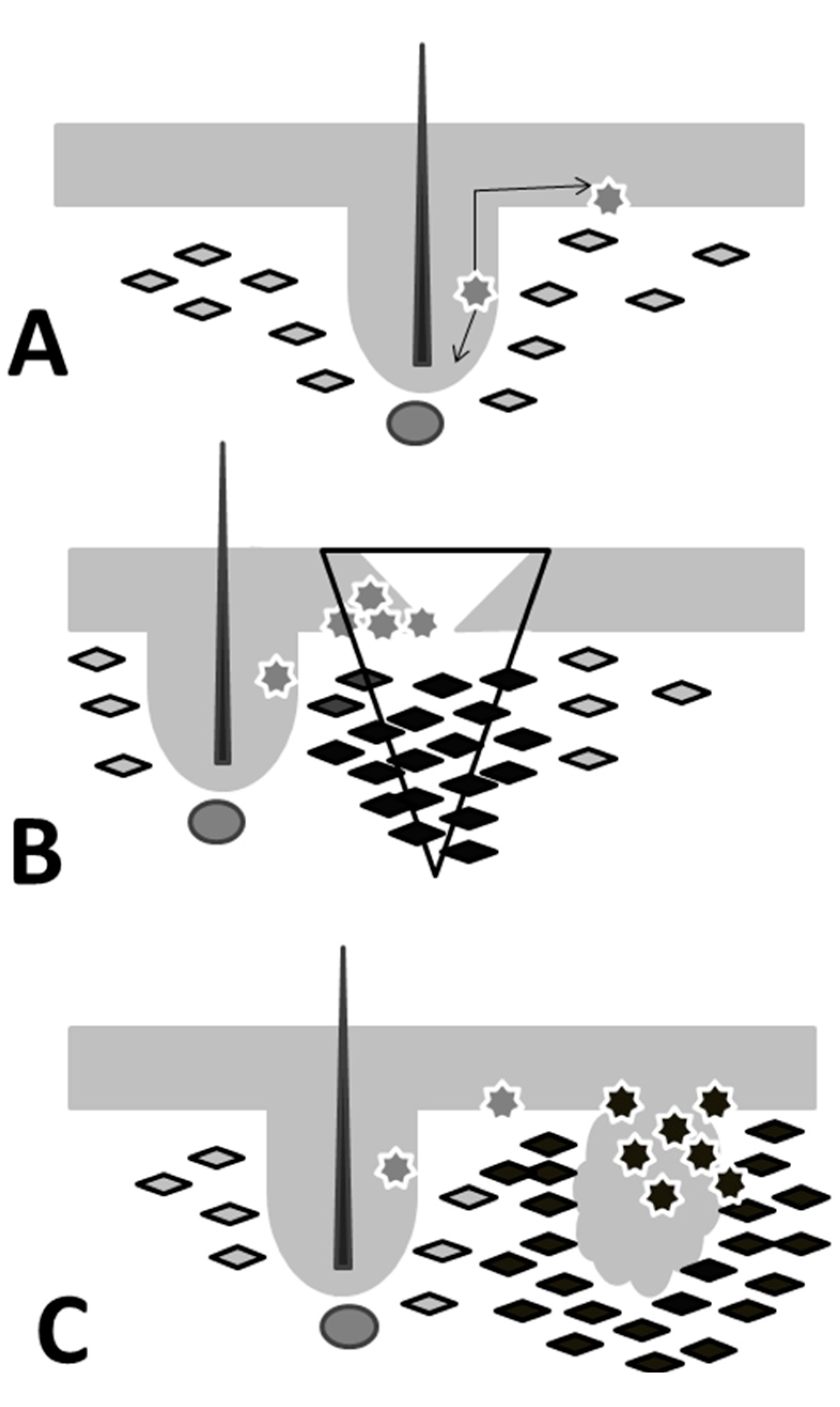{kind=link}
Abstract
:1. Stem Cells: Clinical Expectation and Reality
2. ESCs and Their Niche
3. ESCs and Wound Healing

4. ESCs and Cancer
5. Tumor as Complex Organ with CSC Activity Stimulating Microenvironment
6. CAFs as the Master Cell Type in Tumor Stroma
7. Malignant Melanoma: Microenvironment in Non-Keratinocytic Cutaneous Tumors
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weisdorf, D.; Chao, N.; Waselenko, J.K.; Dainiak, N.; Armitage, J.O.; McNiece, I.; Confer, D. Acute radiation injury: Contingency planning for triage, supportive care, and transplantation. Biol. Blood Marrow Transplant. 2006, 12, 672–682. [Google Scholar]
- Abbasalizadeh, S.; Baharvand, H. Technological progress and challenges towards cGMP manufacturing of human pluripotent stem cells based therapeutic products for allogeneic and autologous cell therapies. Biotechnol. Adv. 2013, 31, 1600–1623. [Google Scholar] [CrossRef] [PubMed]
- Healy, L.; Young, L.; Stacey, G.N. Stem cell banks: Preserving cell lines, maintaining genetic integrity, and advancing research. Methods Mol. Biol. 2011, 767, 15–27. [Google Scholar] [PubMed]
- Bravery, C.A.; French, A. Reference materials for cellular therapeutics. Cytotherapy 2014, 16, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Grasset, N.; Barrandon, Y. Clinical application of autologous epithelial stem cells in disorders of squamous epithelia in translational stem cell research. In Issues beyond the Debate on the Moral Status of the Human Embryo; Hug, K., Hermén, G., Eds.; Springer Science & Business Media: New York, NY, USA, 2010; pp. 45–53. [Google Scholar]
- Rheinwald, J.G.; Green, H. Serial cultivation of strains of human epidermal keratinocytes: The formation of keratinizing colonies from single cells. Cell 1975, 63, 331–343. [Google Scholar] [CrossRef]
- Green, H.; Rheinwald, J.G.; Sun, T.T. Properties of an epithelial cell type in culture: The epidermal keratinocyte and its dependence on products of the fibroblast. Prog. Clin. Biol. Res. 1977, 17, 493–500. [Google Scholar] [PubMed]
- Dvoránková, B.; Smetana, K., Jr.; Königová, R.; Singerová, H.; Vacík, J.; Jelínková, M.; Kapounková, Z.; Zahradník, M. Cultivation and grafting of human keratinocytes on a poly(hydroxyethyl methacrylate) support to the wound bed: A clinical study. Biomaterials 1998, 19, 141–146. [Google Scholar] [CrossRef]
- Dvoránková, B.; Holíková, Z.; Vacík, J.; Königová, R.; Kapounková, Z.; Michálek, J.; Prádný, M.; Smetana, K., Jr. Reconstruction of epidermis by grafting of keratinocytes cultured on polymer support-clinical study. Int. J. Dermatol. 2003, 42, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Labský, J.; Dvoránková, B.; Smetana, K., Jr.; Holíková, Z.; Broz, L.; Gabius, H.-J. Mannosides as crucial part of bioactive supports for cultivation of human epidermal keratinocytes without feeder cells. Biomaterials 2003, 24, 863–872. [Google Scholar] [CrossRef]
- Vacík, J.; Dvoránková, B.; Michálek, J.; Prádný, M.; Krumbholcová, E.; Fenclová, T.; Smetana, K., Jr. Cultivation of human keratinocytes without feeder cells on polymer carriers containing ethoxyethyl methacrylate: In vitro study. J. Mater. Sci. Mater. Med. 2008, 19, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Mcheik, J.N.; Barrault, C.; Levard, G.; Morel, F.; Bernard, F.X.; Lecron, J.C. Epidermal healing in burns: Autologous keratinocyte transplantation as a standard procedure: Update and perspective. Plast. Reconstr. Surg. Glob. Open 2014, 2, e218. [Google Scholar] [CrossRef] [PubMed]
- Bickenbach, J.R. Identification and behavior of label-retaining cells in oral mucosa and skin. J. Dent. Res. 1981, 60, 611–620. [Google Scholar] [CrossRef]
- Williams, S.E.; Beronja, S.; Pasolli, H.A.; Fuchs, E. Asymmetric cell divisions promote Notch-dependent epidermal differentiation. Nature 2011, 470, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Tumbar, T.; Guasch, G.; Greco, V.; Blanpain, C.; Lowry, W.E.; Rendl, M.; Fuchs, E. Defining the epithelial stem cell niche in skin. Science 2004, 303, 359–363. [Google Scholar] [CrossRef]
- Watt, F.M. Mammalian skin cell biology: At the interface between laboratory and clinic. Science 2014, 346, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Alcolea, M.P.; Jones, P.H. Lineage analysis of epidermal stem cells. Cold Spring Harb. Perspect. Med. 2014, 4, a015206. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Fuchs, E. Epidermal stem cells of the skin. Annu. Rev. Cell Dev. Biol. 2006, 22, 339–373. [Google Scholar] [CrossRef] [PubMed]
- Purba, T.S.; Haslam, I.S.; Poblet, E.; Jiménez, F.; Gandarillas, A.; Izeta, A.; Paus, R. Human epithelial hair follicle stem cells and their progeny: Current state of knowledge, the widening gap in translational research and future challenges. Bioessays 2014, 36, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Sieber-Blum, M.; Grim, M. The adult hair follicle: Cradle for pluripotent neural crest stem cells. Birth Defects Res. C Embryo Today Rev. 2004, 72, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Krejčí, E.; Grim, M. Isolation and characterization of neural crest stem cells from adult human hair follicles. Folia Biol. 2010, 56, 149–157. [Google Scholar]
- Tanimura, S.; Tadokoro, Y.; Inomata, K.; Binh, N.T.; Nishie, W.; Yamazaki, S.; Nakauchi, H.; Tanaka, Y.; McMillan, J.R.; Sawamura, D.; et al. Hair follicle stem cells provide a functional niche for melanocyte stem cells. Cell Stem Cell 2011, 8, 177–187. [Google Scholar] [PubMed]
- Chang, C.Y.; Pasolli, H.A.; Giannopoulou, E.G.; Guasch, G.; Gronostajski, R.M.; Elemento, O.; Fuchs, E. NFIB is a governor of epithelial-melanocyte stem cell behaviour in a shared niche. Nature 2013, 495, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.; Horsley, V. Home sweet home: Skin stem cell niches. Cell. Mol. Life Sci. 2012, 69, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Greco, V. Stem cell dynamics in the hair follicle niche. Semin. Cell Dev. Biol. 2014, 25–26, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Alonso, L.; Okada, H.; Pasolli, H.A.; Wakeham, A.; You-Ten, A.I.; Mak, T.W.; Fuchs, E. Sgk3 links growth factor signaling to maintenance of progenitor cells in the hair follicle. J. Cell Biol. 2005, 170, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Folgueras, A.R.; Guo, X.; Pasolli, H.A.; Stokes, N.; Polak, L.; Zheng, D.; Fuchs, E. Architectural niche organization by LHX2 is linked to hair follicle stem cell function. Cell Stem Cell 2013, 133, 314–327. [Google Scholar] [CrossRef]
- Fuchs, E. Finding one’s niche in the skin. Cell Stem Cell 2009, 4, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Harries, M.J.; Meyer, K.; Chaudhry, I.E.; Kloepper, J.; Poblet, E.; Griffiths, C.E.; Paus, R. Lichen planopilaris is characterized by immune privilege collapse of the hair follicle’s epithelial stem cell niche. J. Pathol. 2013, 231, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Bagutti, C.; Hutter, C.; Chiquet-Ehrismann, R.; Fässler, R.; Watt, F.M. Dermal fibroblast-derived growth factors restore the ability of β1integrin-deficient embryonal stem cells to differentiate into keratinocytes. Dev. Biol. 2001, 231, 321–333. [Google Scholar] [CrossRef]
- Rendl, M.; Lewis, L.; Fuchs, E. Molecular dissection of mesenchymal-epithelial interactions in the hair follicle. PLoS Biol. 2005, 311, e331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellheyer, K.; Krahl, D. Skin mesenchymal stem cells: Prospects for clinical dermatology. J. Am. Acad. Dermatol. 2010, 63, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Cotsarelis, G. Review of hair follicle dermal cells. J. Dermatol. Sci. 2010, 57, 2–11. [Google Scholar] [CrossRef]
- Tucker, R.P.; Ferralli, J.; Schittny, J.C.; Chiquet-Ehrismann, R. Tenascin-C and tenascin-W in whisker follicle stem cell niches: Possible roles in regulating stem cell proliferation and migration. J. Cell Sci. 2013, 126, 5111–5115. [Google Scholar] [CrossRef] [PubMed]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Solanas, G.; Benitah, S.A. Regenerating the skin: A task for the heterogeneous stem cell pool and surrounding niche. Nat. Rev. Mol. Cell Biol. 2013, 14, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Sottocornola, R.; Lo Celso, C. Dormancy in the stem cell niche. Stem Cell Res. Ther. 2012, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Rawles, M.E. Tissue interactions in scale and feather development as studied in dermal-epidermal recombinations. J. Embryol. Exp. Morphol. 1963, 11, 765–789. [Google Scholar] [PubMed]
- Ferraris, C.; Chevalier, G.; Favier, B.; Jahoda, C.A.; Dhouailly, D. Adult corneal epithelium basal cells possess the capacity to activate epidermal, pilosebaceous and sweat gland genetic programs in response to embryonic dermal stimuli. Development 2000, 127, 5487–5495. [Google Scholar] [PubMed]
- Blazejewska, E.A.; Schlötzer-Schrehardt, U.; Zenkel, M.; Bachmann, B.; Chankiewitz, E.; Jacobi, C.; Kruse, F.E. Corneal limbal microenvironment can induce transdifferentiation of hair follicle stem cells into corneal epithelial-like cells. Stem Cells 2009, 27, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.E.; Mooney, D.J.; Zandstra, P.W. Growth factors, matrices, and forces combine and control stem cells. Science 2009, 324, 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Das, R.K.; Zouani, O.F. A review of the effects of the cell environment physicochemical nanoarchitecture on stem cell commitment. Biomaterials 2014, 35, 5278–5293. [Google Scholar] [CrossRef] [PubMed]
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Morasso, M.I.; Tomic-Canic, M. Epidermal stem cells: The cradle of epidermal determination, differentiation and wound healing. Biol. Cell 2005, 97, 173–183. [Google Scholar] [CrossRef]
- Bickenbach, J.R.; Stern, M.M.; Grinnell, K.L.; Manuel, A.; Chinnathambi, S. Epidermal stem cells have the potential to assist in healing damaged tissues. J. Investig. Dermatol. Symp. Proc. 2006, 11, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Chai, L.; Chen, L.; Chen, W.; Ge, L.; Li, X.; Li, H.; Li, S.; Cao, C. Stromal cell-derived factor 1 (SDF-1) accelerated skin wound healing by promoting the migration and proliferation of epidermal stem cells. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 5768–5785. [Google Scholar] [CrossRef] [PubMed]
- Ghahary, A.; Ghaffari, A. Role of keratinocyte-fibroblast cross-talk in development of hypertrophic scar. Wound Repair Regen. 2007, 15, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Laverdet, B.; Micallef, L.; Lebreton, C.; Mollard, J.; Lataillade, J.J.; Coulomb, B.; Desmoulière, A. Use of mesenchymal stem cells for cutaneous repair and skin substitute elaboration. Pathol. Biol. 2014, 62, 108–117. [Google Scholar] [CrossRef]
- Stojadinovic, O.; Pastar, I.; Nusbaum, A.G.; Vukelic, S.; Krzyzanowska, A.; Tomic-Canic, M. Deregulation of epidermal stem cell niche contributes to pathogenesis of nonhealing venous ulcers. Wound Repair Regen. 2014, 22, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Wu, C.S.; Chao, D.; Wu, C.H.; Li, C.C.; Chen, G.S.; Lan, C.C. High-glucose cultivated peripheral blood mononuclear cells impaired keratinocyte function via reduced IL-22 expression: Implications on impaired diabetic wound healing. Exp. Dermatol. 2015, 24, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Dvořánková, B.; Szabo, P.; Lacina, L.; Gal, P.; Uhrova, J.; Zima, T.; Kaltner, H.; André, S.; Gabius, H.-J.; Sykova, E.; et al. Human galectins induce conversion of dermal fibroblasts into myofibroblasts and production of extracellular matrix: Potential application in tissue engineering and wound repair. Cells Tissues Organs 2011, 194, 469–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mifková, A.; Kodet, O.; Szabo, P.; Kučera, J.; Dvořánková, B.; André, S.; Koripelly, G.; Gabius, H.-J.; Lehn, J.-M.; Smetana, K., Jr. Synthetic polyamine BPA-C8 inhibits TGF-β1-mediated conversion of human dermal fibroblast to myofibroblasts and establishment of galectin-1-rich extracellular matrix in vitro. ChemBioChem 2014, 15, 1465–1470. [Google Scholar] [CrossRef] [PubMed]
- Van De Water, L.; Varney, S.; Tomasek, J.J. Mechanoregulation of the myofibroblast in wound contraction, scarring, and fibrosis: Opportunities for new therapeutic intervention. Adv. Wound Care 2013, 2, 122–141. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Iredale, J.P.; Fallowfield, J.A. Resolution of liver fibrosis: Basic mechanisms and clinical relevance. Semin. Liver Dis. 2015, 35, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Kolář, M.; Szabo, P.; Dvořánková, B.; Lacina, L.; Gabius, H.-J.; Strnad, H.; Sáchová, J.; Vlček, C.; Plzák, J.; Chovanec, M.; et al. Upregulation of IL-6, IL-8 and CXCL-1 production in dermal fibroblasts by normal/malignant epithelial cells in vitro: Immunohistochemical and transcriptomic analyses. Biol. Cell 2012, 104, 738–751. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.Z.; Pourreyron, C.; Salas-Alanis, J.C.; Dayal, J.H.; Cepeda-Valdes, R.; Yan, W.; Wright, S.; Chen, M.; Fine, J.D.; Hogg, F.J.; et al. Fibroblast-derived dermal matrix drives development of aggressive cutaneous squamous cell carcinoma in patients with recessive dystrophic epidermolysis bullosa. Cancer Res. 2012, 72, 3522–3534. [Google Scholar] [CrossRef] [PubMed]
- Rybinski, B.; Franco-Barraza, J.; Cukierman, E. The wound healing, chronic fibrosis, and cancer progression triad. Physiol. Genom. 2014, 46, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, A.W.; Salmon, S.E. Primary bioassay of human tumor stem cells. Science 1977, 197, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Motlík, J.; Klíma, J.; Dvoránková, B.; Smetana, K., Jr. Porcine epidermal stem cells as a biomedical model for wound healing and normal/malignant epithelial cell propagation. Theriogenology 2007, 67, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Perez-Losada, J.; Balmain, A. Stem-cell hierarchy in skin cancer. Nat. Rev. Cancer 2003, 3, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Song, I.Y.; Balmain, A. Cellular reprogramming in skin cancer. Semin. Cancer Biol. 2015, 32, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, Y.; Mao, J.H.; Brown, K.; Girardi, M.; Balmain, A. Promotion of Hras-induced squamous carcinomas by a polymorphic variant of the Patched gene in FVB mice. Nature 2007, 445, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Varjosalo, M.; Taipale, J. Hedgehog: Functions and mechanisms. Genes Dev. 2008, 22, 2454–2472. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.D.; Bazzi, H.; Fantauzzo, K.A.; Waters, J.M.; Crawford, H.; Hynd, P.; Christiano, A.M.; Jahoda, C.A. KGF and EGF signalling block hair follicle induction and promote interfollicular epidermal fate in developing mouse skin. Development 2009, 136, 2153–2164. [Google Scholar] [CrossRef] [PubMed]
- Nitzki, F.; Becker, M.; Frommhold, A.; Schulz-Schaeffer, W.; Hahn, H. Patched knockout mouse models of Basal cell carcinoma. J. Skin Cancer 2012, 2012, 907543. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, C.; Vogelstein, B. Cancer etiology. Variation in cancer risk among tissues can be explained by the number of stem cell divisions. Science 2015, 347, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Lombard, D.B.; Chua, K.F.; Mostoslavsky, R.; Franco, S.; Gostissa, M.; Alt, F.W. DNA repair, genome stability, and aging. Cell 2005, 120, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Sharpless, N.E.; DePinho, R.A. How stem cells age and why this makes us grow old. Nat. Rev. Mol. Cell Biol. 2007, 8, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Adjaye, J.; Akamatsu, H.; Moe-Behrens, G.; Niemann, C. Human skin stem cells and the ageing process. Exp. Gerontol. 2008, 43, 986–997. [Google Scholar] [CrossRef] [PubMed]
- Mackenbach, J.P. Political conditions and life expectancy in Europe, 1900–2008. Soc. Sci. Med. 2013, 82, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Smetana, K., Jr.; Dvořánková, B.; Lacina, L. Phylogeny, regeneration, ageing and cancer: Role of microenvironment and possibility of its therapeutic manipulation. Folia Biol. 2013, 59, 207–216. [Google Scholar]
- Egeblad, M.; Nakasone, E.S.; Werb, Z. Tumors as organs: Complex tissues that interface with the entire organism. Dev. Cell 2010, 18, 884–901. [Google Scholar] [CrossRef] [PubMed]
- Plaks, V.; Kong, N.; Werb, Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F. Tumors: Wounds that do not heal. Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [PubMed]
- Plzák, J.; Lacina, L.; Chovanec, M.; Dvoránková, B.; Szabo, P.; Cada, Z.; Smetana, K., Jr. Epithelial-stromal interaction in squamous cell epithelium-derived tumors: An important new player in the control of tumor biological properties. Anticancer Res. 2010, 30, 455–462. [Google Scholar] [PubMed]
- Karagiannis, G.S.; Petraki, C.; Prassas, I.; Saraon, P.; Musrap, N.; Dimitromanolakis, A.; Diamandis, E.P. Proteomic signatures of the desmoplastic invasion front reveal collagen type XII as a marker of myofibroblastic differentiation during colorectal cancer metastasis. Oncotarget 2012, 3, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Medler, T.R.; Coussens, L.M. Duality of the immune response in cancer: Lessons learned from skin. J. Investig. Dermatol. 2014, 134, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Champer, J.; Beynet, D.; Kim, J.; Friedman, A.J. The role of the cutaneous microbiome in skin cancer: Lessons learned from the gut. J. Drugs Dermatol. 2015, 14, 461–465. [Google Scholar] [PubMed]
- Muranushi, C.; Olsen, C.M.; Pandeya, N.; Green, A.C. Aspirin and nonsteroidal anti-inflammatory drugs can prevent cutaneous squamous cell carcinoma: A systematic review and meta-analysis. J. Investig. Dermatol. 2015, 135, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Etrych, T.; Subr, V.; Strohalm, J.; Sírová, M.; Ríhová, B.; Ulbrich, K. HPMA copolymer-doxorubicin conjugates: The effects of molecular weight and architecture on biodistribution and in vivo activity. J. Control. Release 2012, 164, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Polyak, K.; Haviv, I.; Campbell, I.G. Co-evolution of tumor cells and their microenvironment. Trends Genet. 2009, 25, 30–38. [Google Scholar] [CrossRef] [PubMed]
- De Wever, O.; Demetter, P.; Mareel, M.; Bracke, M. Stromal myofibroblasts are drivers of invasive cancer growth. Int. J. Cancer 2008, 123, 2229–2238. [Google Scholar] [CrossRef] [PubMed]
- Petersen, O.W.; Nielsen, H.L.; Gudjonsson, T.; Villadsen, R.; Rank, F.; Niebuhr, E.; Bissell, M.J.; Rønnov-Jessen, L. Epithelial to mesenchymal transition in human breast cancer can provide a nonmalignant stroma. Am. J. Pathol. 2003, 162, 391–402. [Google Scholar] [CrossRef]
- Smetana, K., Jr.; Dvoránková, B.; Lacina, L.; Cada, Z.; Vonka, V. Human hair follicle and interfollicular keratinocyte reactivity to mouse HPV16-transformed cells: An in vitro study. Oncol. Rep. 2008, 20, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Dvořánková, B.; Smetana, K., Jr.; Říhová, B.; Kučera, J.; Mateu, R.; Szabo, P. Cancer-associated fibroblasts are not formed from cancer cells by epithelial-to-mesenchymal transition in nu/nu mice. Histochem. Cell Biol. 2015, 143, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Hu, M.; Sridhar, A.; Opeskin, K.; Fox, S.; Shipitsin, M.; Trivett, M.; Thompson, E.R.; Ramakrishna, M.; Gorringe, K.L.; et al. No evidence of clonal somatic genetic alterations in cancer-associated fibroblasts from human breast and ovarian carcinomas. Nat. Genet. 2008, 40, 650–655. [Google Scholar] [CrossRef]
- Rhim, A.D.; Oberstein, P.E.; Thomas, D.H.; Mirek, E.T.; Palermo, C.F.; Sastra, S.A.; Dekleva, E.N.; Saunders, T.; Becerra, C.P.; Tattersall, I.W.; et al. Stromal elements act to restrain, rather than support, pancreatic ductal adenocarcinoma. Cancer Cell 2014, 25, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Lacina, L.; Dvoránkova, B.; Smetana, K., Jr.; Chovanec, M.; Plzák, J.; Tachezy, R.; Kideryová, L.; Kucerová, L.; Cada, Z.; Boucek, J.; et al. Marker profiling of normal keratinocytes identifies the stroma from squamous cell carcinoma of the oral cavity as a modulatory microenvironment in co-culture. Int. J. Radiat. Biol. 2007, 83, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Lacina, L.; Smetana, K., Jr.; Dvoránková, B.; Pytlík, R.; Kideryová, L.; Kucerová, L.; Plzáková, Z.; Stork, J.; Gabius, H.-J.; André, S. Stromal fibroblasts from basal cell carcinoma affect phenotype of normal keratinocytes. Br. J. Dermatol. 2007, 156, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Kideryová, L.; Lacina, L.; Dvoránková, B.; Stork, J.; Cada, Z.; Szabo, P.; André, S.; Kaltner, H.; Gabius, H.-J.; Smetana, K., Jr. Phenotypic characterization of human keratinocytes in coculture reveals differential effects of fibroblasts from benign fibrous histiocytoma (dermatofibroma) as compared to cells from its malignant form and to normal fibroblasts. J. Dermatol. Sci. 2009, 55, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Strnad, H.; Lacina, L.; Kolár, M.; Cada, Z.; Vlcek, C.; Dvoránková, B.; Betka, J.; Plzák, J.; Chovanec, M.; Sáchová, J.; et al. Head and neck squamous cancer stromal fibroblasts produce growth factors influencing phenotype of normal human keratinocytes. Histochem. Cell Biol. 2010, 133, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Jarkovska, K.; Dvorankova, B.; Halada, P.; Kodet, O.; Szabo, P.; Gadher, S.J.; Motlik, J.; Kovarova, H.; Smetana, K., Jr. Revelation of fibroblast protein commonalities and differences and their possible roles in wound healing and tumourigenesis using co-culture models of cells. Biol. Cell 2014, 106, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Szabo, P.; Valach, J.; Smetana, K., Jr.; Dvořánková, B. Comparative analysis of IL-8 and CXCL-1 production by normal and cancer stromal fibroblasts. Folia Biol. 2013, 59, 134–137. [Google Scholar]
- Jiang, L.; Gonda, T.A.; Gamble, M.V.; Salas, M.; Seshan, V.; Tu, S.; Twaddell, W.S.; Hegyi, P.; Lazar, G.; Steele, I.; et al. Global hypomethylation of genomic DNA in cancer-associated myofibroblasts. Cancer Res. 2008, 68, 9900–9908. [Google Scholar] [CrossRef] [PubMed]
- Wenner, C.E.; Yan, S. Biphasic role of TGF-beta1 in signal transduction and crosstalk. J. Cell. Physiol. 2003, 196, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Bierie, B.; Stover, D.G.; Abel, T.W.; Chytil, A.; Gorska, A.E.; Aakre, M.; Forrester, E.; Yang, L.; Wagner, K.U.; Moses, H.L. Transforming growth factor-beta regulates mammary carcinoma cell survival and interaction with the adjacent microenvironment. Cancer Res. 2008, 68, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Smetana, K., Jr.; Dvořánková, B.; Lacina, L.; Strnad, H.; Kolář, M.; Chovanec, M.; Plzák, J.; Čada, Z.; Vlček, Č.; Szabo, P.; et al. Combination of Antibodies or Their Fab Fragmennts for Use as Therapeutics and Pharmaceutic Tool Containing These Antibodies or their Fab Fragments. Czech Patent No. 303227, 22 April 2012. [Google Scholar]
- Karnoub, A.E.; Dash, A.B.; Vo, A.P.; Sullivan, A.; Brooks, M.W.; Bell, G.W.; Richardson, A.L.; Polyak, K.; Tubo, R.; Weinberg, R.A. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature 2007, 449, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Tsukuda, K.; Tsuji, H.; Kunitomo, T.; Aokage, K.; Miyake, T.; Nakahara, S.; Masuda, H. Breast cancer with cartilaginous and/or osseous metaplasia diagnosed by lymph nodal metastasis: A case report. Acta Med. Okayama 2009, 63, 367–371. [Google Scholar] [PubMed]
- Szabo, P.; Kolář, M.; Dvořánková, B.; Lacina, L.; Štork, J.; Vlček, Č.; Strnad, H.; Tvrdek, M.; Smetana, K., Jr. Mouse 3T3 fibroblasts under the influence of fibroblasts isolated from stroma of human basal cell carcinoma acquire properties of multipotent stem cells. Biol. Cell 2011, 103, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Berndt, A.; Richter, P.; Kosmehl, H.; Franz, M. Tenascin-C and carcinoma cell invasion in oral and urinary bladder cancer. Cell Adhes. Migr. 2015, 9, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Nishi, Y.; Sano, H.; Kawashima, T.; Okada, T.; Kuroda, T.; Kikkawa, K.; Kawashima, S.; Tanabe, M.; Goto, T.; Matsuzawa, Y.; et al. Role of galectin-3 in human pulmonary fibrosis. Allergol. Int. 2007, 56, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Valach, J.; Fík, Z.; Strnad, H.; Chovanec, M.; Plzák, J.; Cada, Z.; Szabo, P.; Sáchová, J.; Hroudová, M.; Urbanová, M.; et al. Smooth muscle actin-expressing stromal fibroblasts in head and neck squamous cell carcinoma: Increased expression of galectin-1 and induction of poor prognosis factors. Int. J. Cancer 2012, 131, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Peržeľová, V.; Varinská, L.; Dvořánková, B.; Szabo, P.; Spurný, P.; Valach, J.; Mojžiš, J.; André, S.; Gabius, H.-J.; Smetana, K., Jr.; et al. Extracellular matrix of galectin-1-exposed dermal and tumor-associated fibroblasts favors growth of human umbilical vein endothelial cells in vitro: A short report. Anticancer Res. 2014, 34, 3991–3996. [Google Scholar] [PubMed]
- Smetana, K., Jr.; Szabo, P.; Gal, P.; André, S.; Gabius, H.-J.; Kodet, O.; Dvořánková, B. Emerging role of tissue lectins as microenvironmental effectors in tumors and wounds. Histol. Histopathol. 2015, 30, 293–309. [Google Scholar] [PubMed]
- Díez-Torre, A.; Andrade, R.; Eguizábal, C.; López, E.; Arluzea, J.; Silió, M.; Aréchaga, J. Reprogramming of melanoma cells by embryonic microenvironments. Int. J. Dev. Biol. 2009, 53, 1563–1568. [Google Scholar] [CrossRef]
- Haass, N.K.; Ripperger, D.; Wladykowski, E.; Dawson, P.; Gimotty, P.A.; Blome, C.; Fischer, F.; Schmage, P.; Moll, I.; Brandner, J.M. Melanoma progression exhibits a significant impact on connexin expression patterns in the epidermal tumor microenvironment. Histochem. Cell Biol. 2010, 133, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Kodet, O.; Lacina, L.; Krejčí, E.; Dvořánková, B.; Grim, M.; Štork, J.; Kodetová, D.; Vlček, Č.; Šáchová, J.; Kolář, M.; et al. Melanoma cells influence the differentiation pattern of human epidermal keratinocytes. Mol. Cancer 2015, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Suh, E.K.; Han, I.O.; Oh, E.S. Keratinocyte-derived laminin-332 promotes adhesion and migration in melanocytes and melanoma. J. Biol. Chem. 2011, 286, 13438–13447. [Google Scholar] [CrossRef] [PubMed]
- Dvořánková, B.; Szabo, P.; Lacina, L.; Kodet, O.; Matoušková, E.; Smetana, K., Jr. Fibroblasts prepared from different types of malignant tumors stimulate expression of luminal marker keratin 8 in the EM-G3 breast cancer cell line. Histochem. Cell Biol. 2012, 137, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Kodet, O.; Dvořánková, B.; Krejčí, E.; Szabo, P.; Dvořák, P.; Štork, J.; Krajsová, I.; Dundr, P.; Smetana, K., Jr.; Lacina, L. Cultivation-dependent plasticity of melanoma phenotype. Tumor Biol. 2013, 34, 3345–3355. [Google Scholar] [CrossRef] [PubMed]
- Kučera, J.; Dvořánková, B.; Smetana, K., Jr.; Szabo, P.; Kodet, O. Fibroblasts isolated from the malignant melanoma influence phenotype of normal human keratinocytes. J. Appl. Biomed. 2015, 13, 195–198. [Google Scholar] [CrossRef]
- Trylcova, J.; Busek, P.; Smetana, K., Jr.; Balaziova, E.; Dvorankova, B.; Mifkova, A.; Sedo, A. Effect of cancer-associated fibroblasts on the migration of glioma cells in vitro. Tumor Biol. 2015, 36, 5873–5879. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacina, L.; Plzak, J.; Kodet, O.; Szabo, P.; Chovanec, M.; Dvorankova, B.; Smetana Jr., K. Cancer Microenvironment: What Can We Learn from the Stem Cell Niche. Int. J. Mol. Sci. 2015, 16, 24094-24110. https://doi.org/10.3390/ijms161024094
Lacina L, Plzak J, Kodet O, Szabo P, Chovanec M, Dvorankova B, Smetana Jr. K. Cancer Microenvironment: What Can We Learn from the Stem Cell Niche. International Journal of Molecular Sciences. 2015; 16(10):24094-24110. https://doi.org/10.3390/ijms161024094
Chicago/Turabian StyleLacina, Lukas, Jan Plzak, Ondrej Kodet, Pavol Szabo, Martin Chovanec, Barbora Dvorankova, and Karel Smetana Jr. 2015. "Cancer Microenvironment: What Can We Learn from the Stem Cell Niche" International Journal of Molecular Sciences 16, no. 10: 24094-24110. https://doi.org/10.3390/ijms161024094
APA StyleLacina, L., Plzak, J., Kodet, O., Szabo, P., Chovanec, M., Dvorankova, B., & Smetana Jr., K. (2015). Cancer Microenvironment: What Can We Learn from the Stem Cell Niche. International Journal of Molecular Sciences, 16(10), 24094-24110. https://doi.org/10.3390/ijms161024094