
Genotypic Variation under Fe Deficiency Results in Rapid Changes in Protein Expressions and Genes Involved in Fe Metabolism and Antioxidant Mechanisms in Tomato Seedlings (Solanum lycopersicum L.)



Abstract
:1. Introduction

2. Results
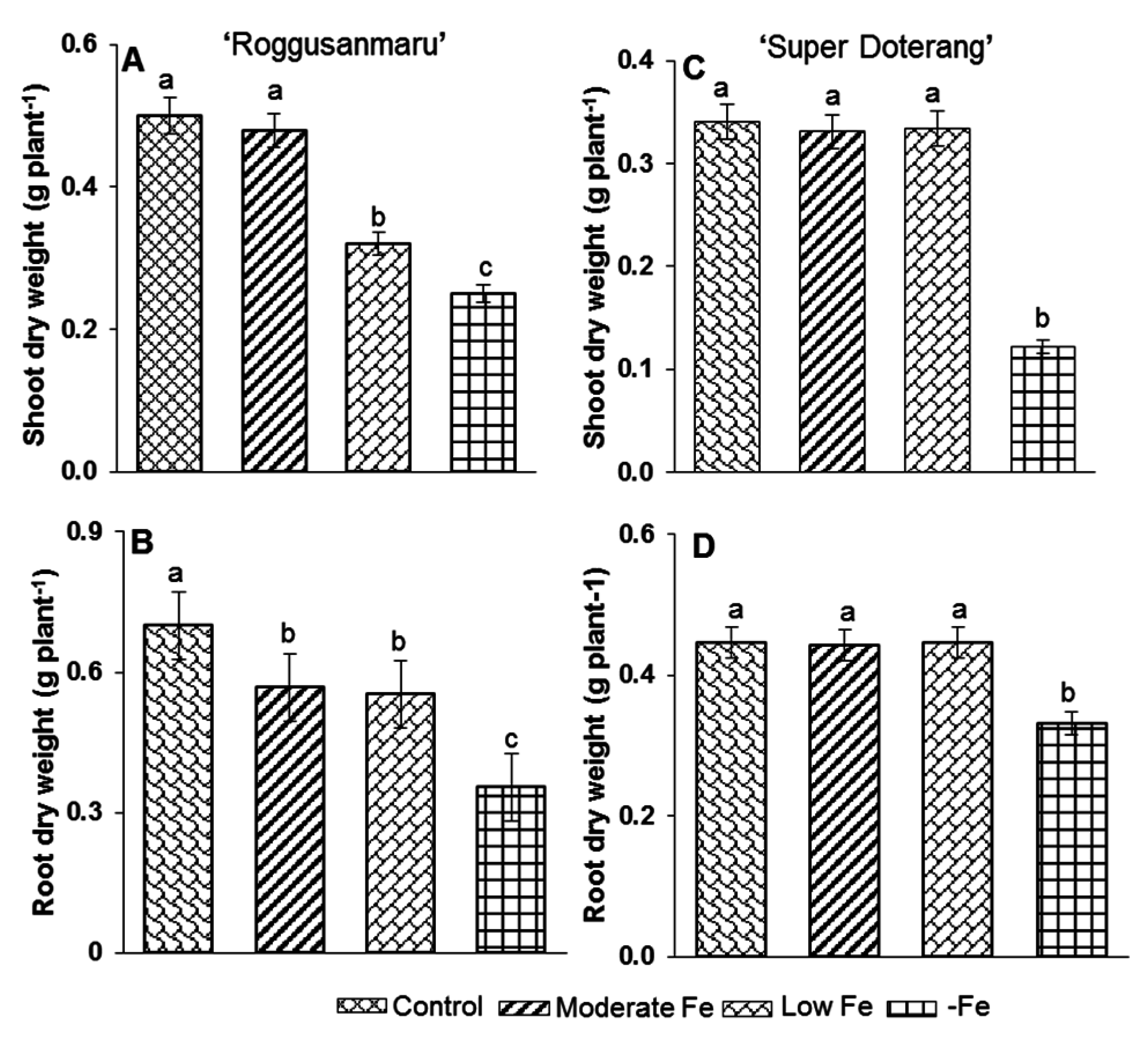
2.1. Plant Growth Parameters

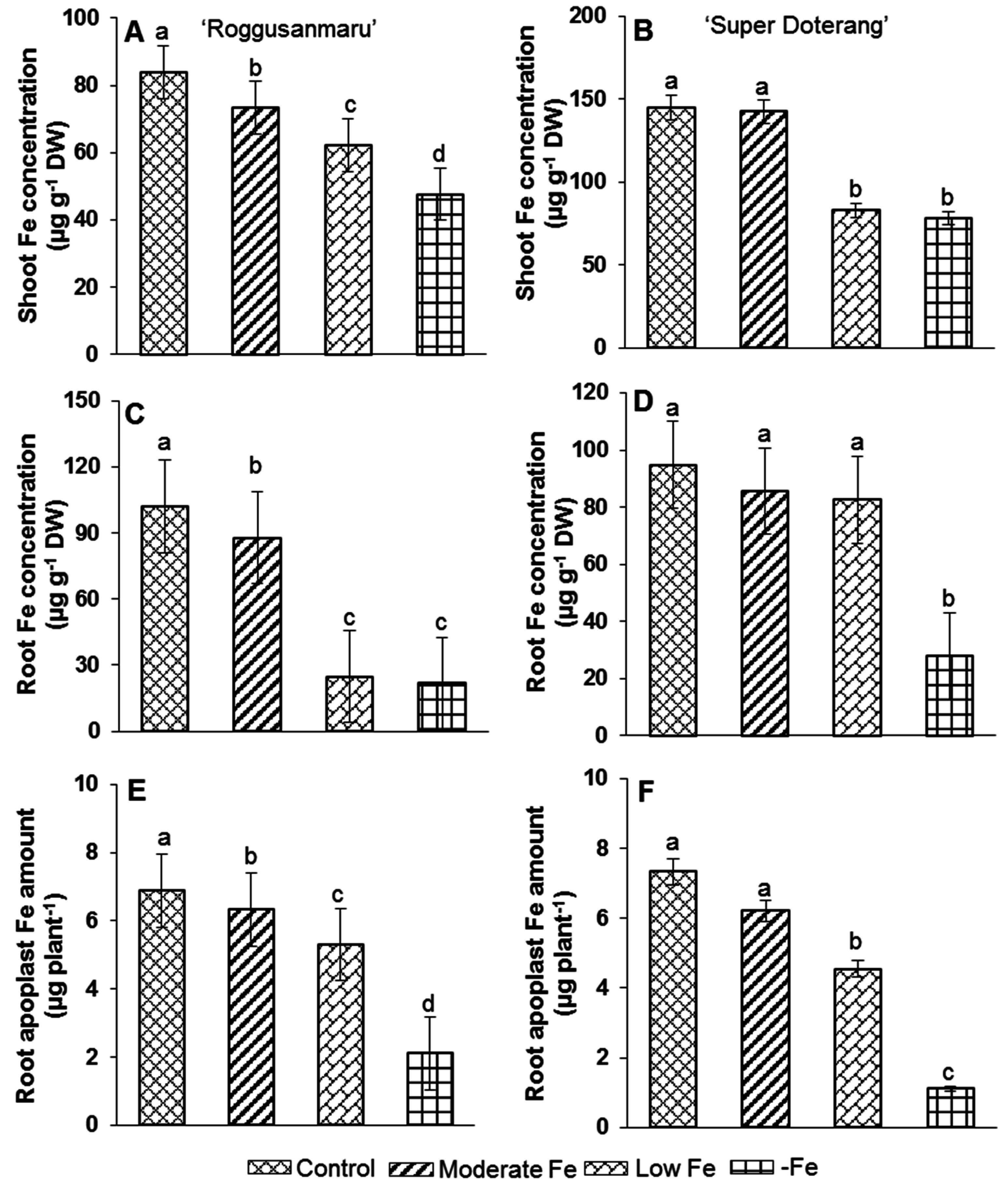
2.2. Total Fe Concentration and Apoplastic Fe Amount

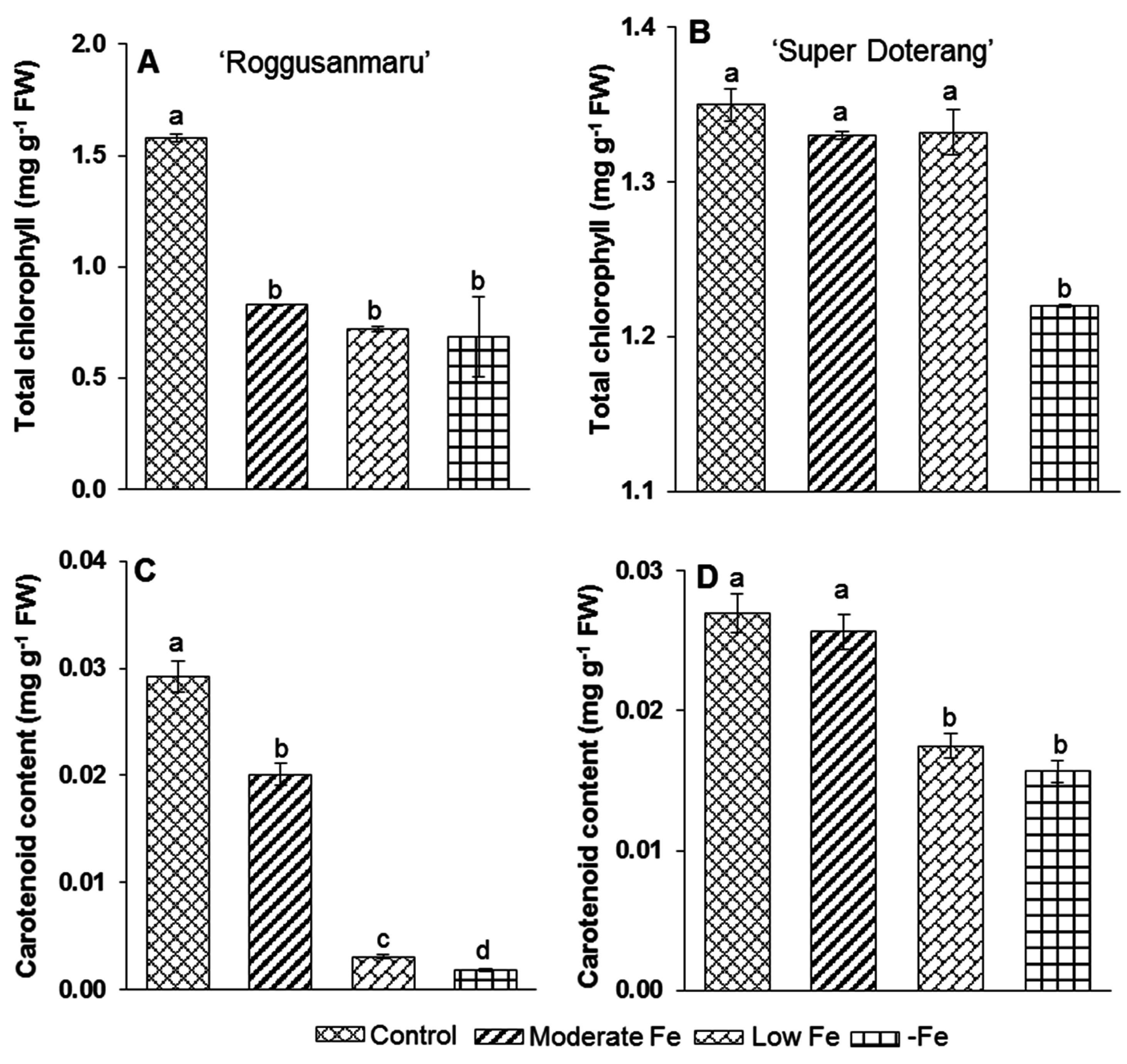
2.3. Pigment Analysis

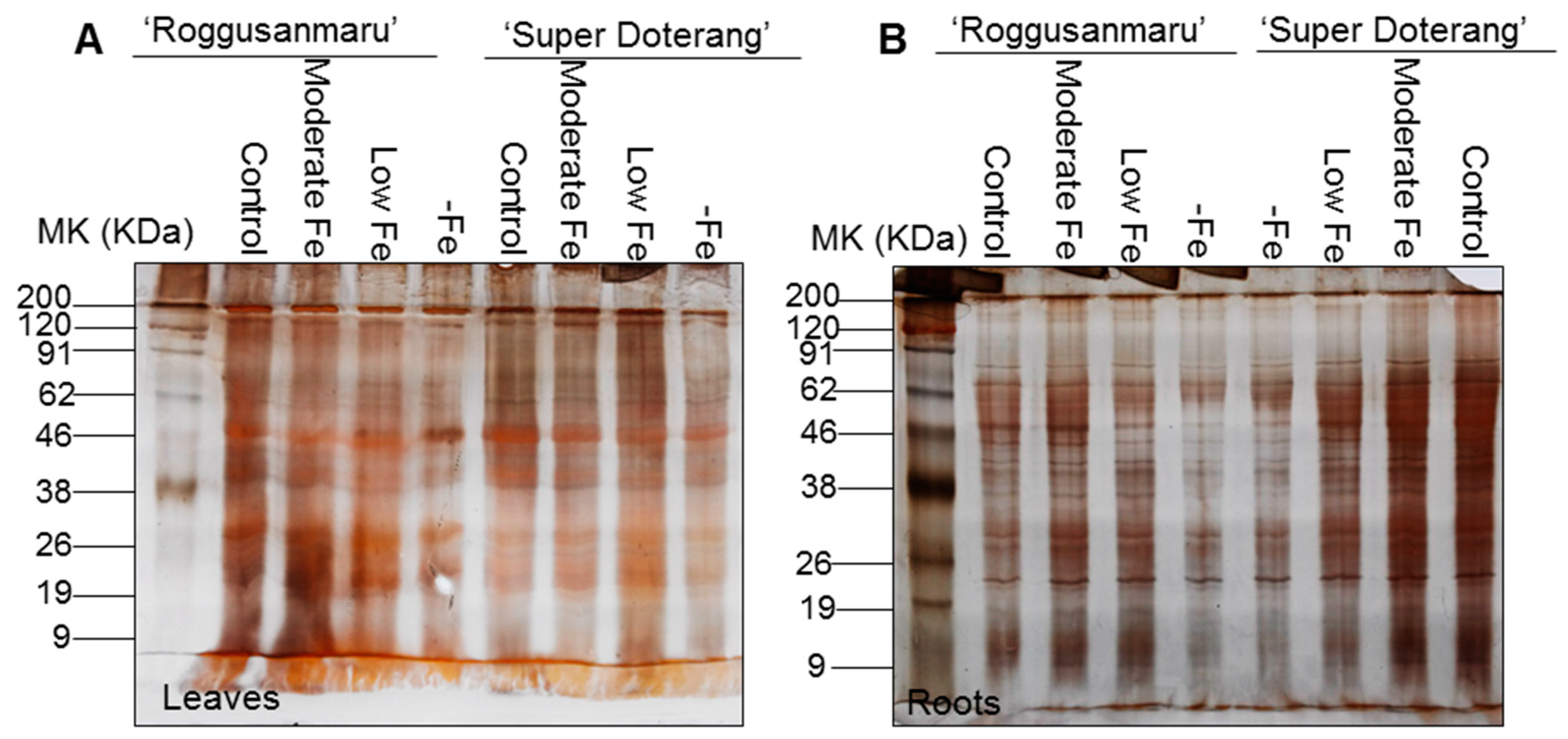
2.4. Total Protein Profile Content

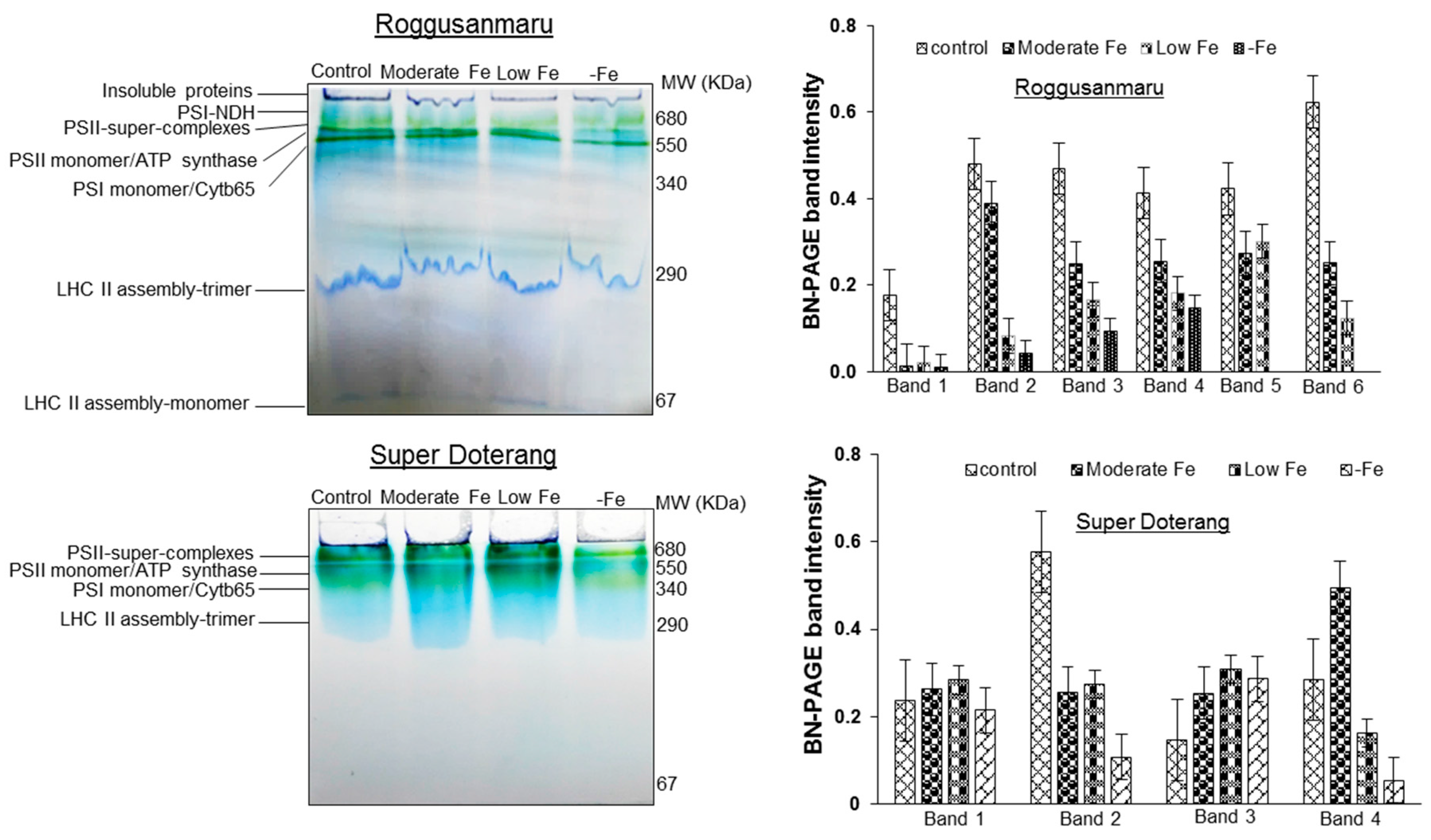
2.5. Changes in Multiprotein Complex Proteins (MCPs) (1D-BN-PAGE)

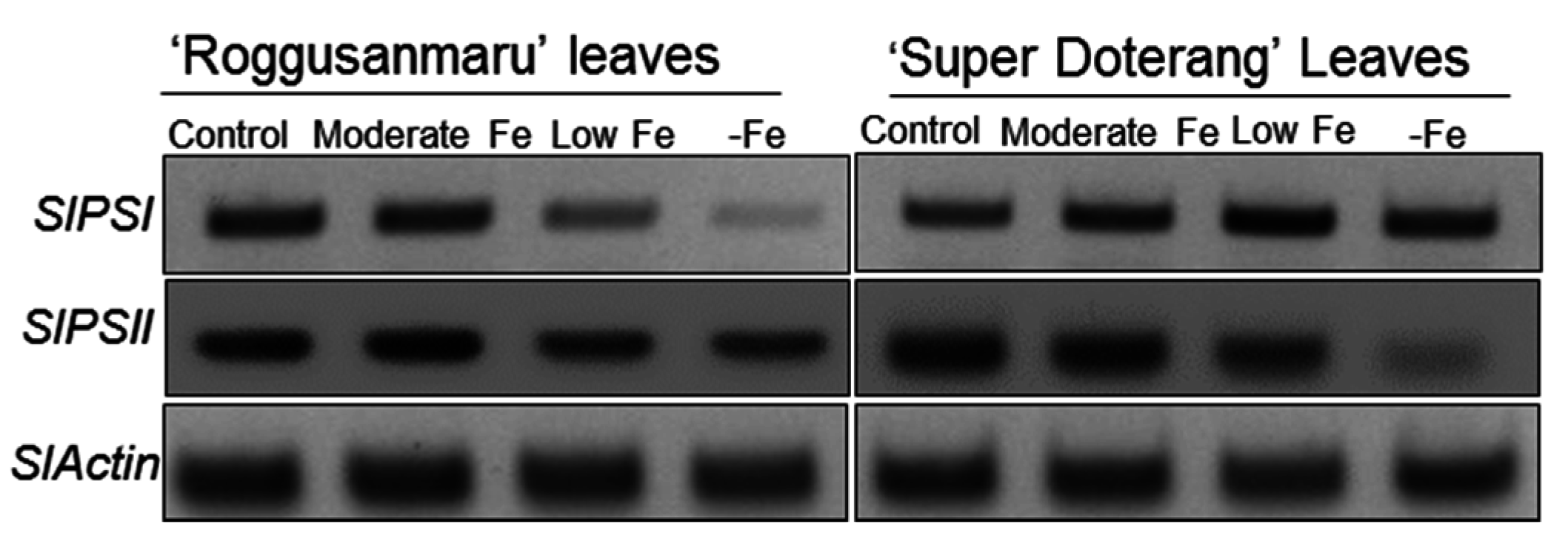
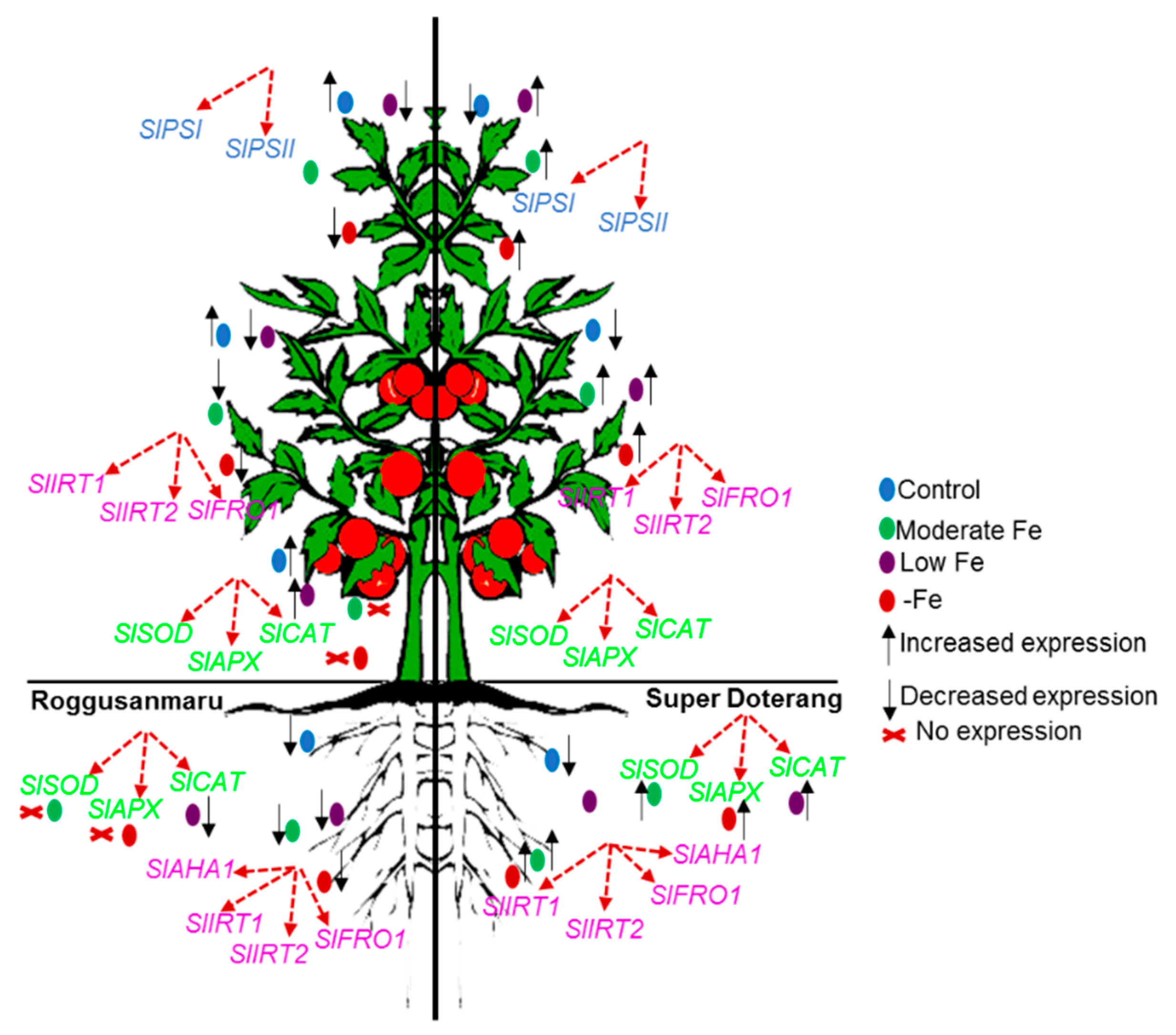
2.6. Effect of Fe-Deficiency on SlPSI and SlPSII
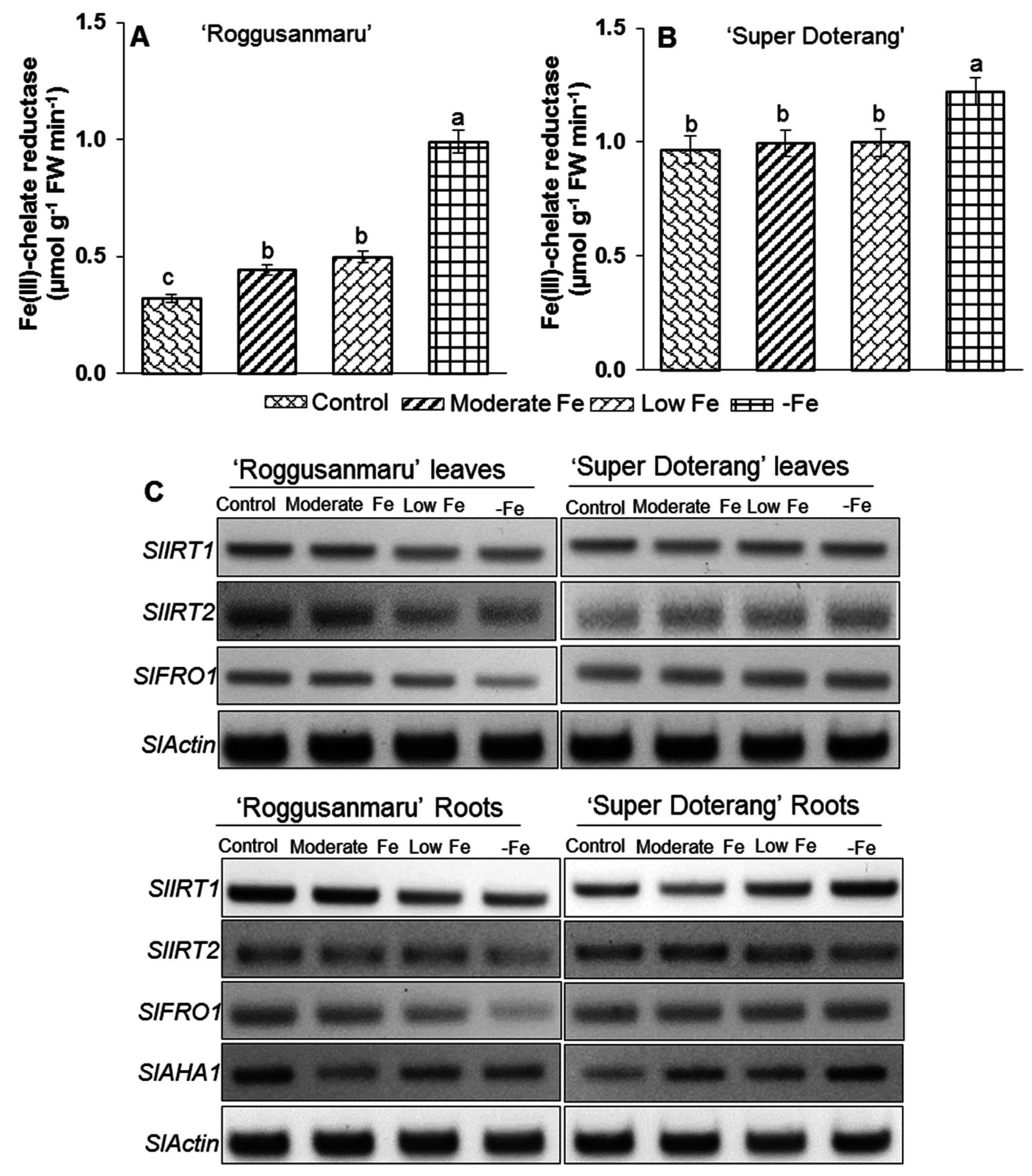
2.7. Effect of Fe-Deficiency on Fe-Transport and Fe(III)-Chelate Activity


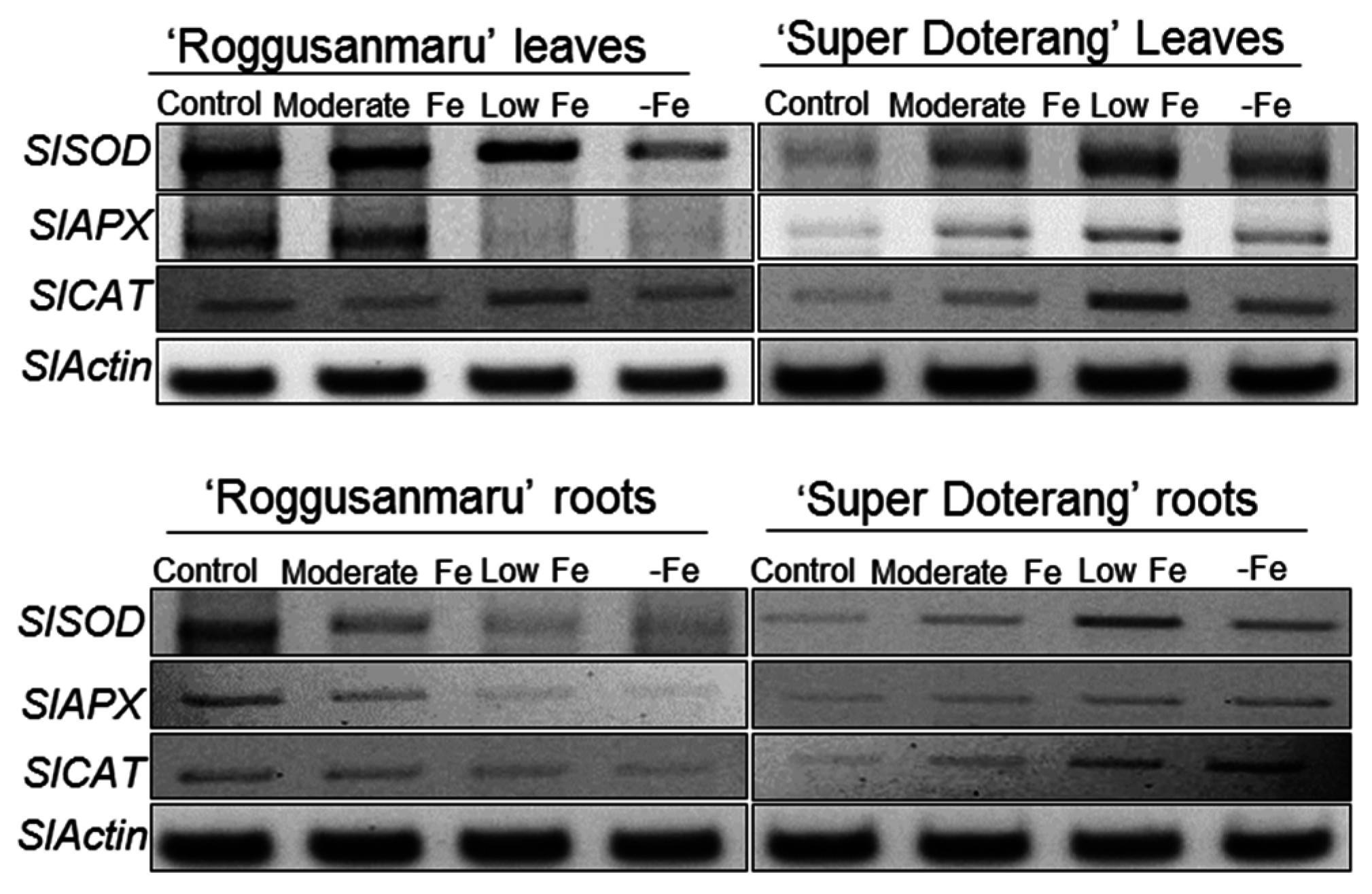
2.8. Effect of Fe-Deficiency on Antioxidant Mechanisms

3. Discussion

4. Material and Methods
4.1. Plant Materials and Growth Conditions
4.2. Biomass Analysis
4.3. Analysis of Fe Concentration and Root Apoplastic Fe-Amount
4.4. Pigment Determination
4.5. Total Protein Profile by SDS-PAGE (Sodium Dodecyl Sulfate Polyacryalamide Gel Elctrophoresis)
4.6. Analysis of Multiprotein Complex Proteins in Thylakoids (MCPs) (First-Dimensional Blue Native Page) (1D-BN-PAGE)
4.7. Image Analysis
4.8. Determination of Fe3+-EDTA Reduction Activity
4.9. Isolation of RNA, cDNA Preparation and Semi-Quantitative RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|
| SlPSI | DQ525206.1 | 5′-ATAAGGGTCTTTATGACACA-3′ | 5′-TATAATGCAGCTTGAGTAGT-3′ |
| SlPSII | NM_001247113.2 | 5′-ATGGCAAGCACAGTAATGAG-3′ | 5′-TCTCAAGGCCATGCTTCCAT-3′ |
| SlIRT1 | AF136579.1 | 5′-TATTTTGTAAAATCCAGATA-3′ | 5′-ACAATATTTTTTGAATAGTG-3′ |
| SlIRT2 | AF136580.1 | 5′-ATTCCTACTCAAAATTCAAA-3′ | 5′-ACAAACTCCGATCATACTAG-3′ |
| SlFRO1 | NM_001247400.1 | 5′-ATGGCTCAAACATCCTCTTC-3′ | 5′-GAGAGGAAATGTGTTCATCA-3′ |
| SlAHA1 | M60166.1 | 5′-AAATCTTGTCCTCTTTTCTT-3′ | 5′-AAAATCCTGCCAGTCCGGTG-3′ |
| SlAPX | NM001247702 | 5′-AAGGCCACATTCTGTCATCC-3′ | 5′-TCAAATTTCAGCCACTGCAC-3′ |
| SlSOD | M37151 | 5′-CTGGACTTCACGGGTTTCAT-3′ | 5′-CCCGGAGAGGAGGGTAAATA-3′ |
| SlCAT | NM001247898 | 5′-TGCATTGAAACCAAATCCAA-3′ | 5′-CAACACCAATCGACCAACTG-3′ |
| SlActin | FJ532351.1 | 5′-ATGACTCAAATCATGTTTGAG-3′ | 5′-GCAGCATGAAGATTAAGGTA-3′ |
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Balk, J.; Pilon, M. Ancient and essential: the assembly of iron–sulfur clusters in plants. Trends Plant Sci. 2011, 16, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Paolacci, A.R.; Celletti, S.; Catarcione, G.; Hawkesford, M.J.; Astolfi, S.; Ciaffi, M. Iron deprivation results in a rapid but not sustained increase of the expression of genes involved in iron metabolism and sulfate uptake in tomato (Solanum lycopersicum L.) seedlings. J. Int. Plant Biol. 2014, 56, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Hakeem, K.R.; Mohamed, R.; Lee, J.H. Cadmium toxicity induced alterations in the root proteome of green gram in contrasting response towards iron supplement. Int. J. Mol. Sci. 2014, 15, 6343–6355. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, W.L.; Schwab, A.P. The chemistry of iron in soils and its availability to plants. J. Plant Nutr. 1982, 5, 821–840. [Google Scholar] [CrossRef]
- Muneer, S.; Kim, T.H.; Qureshi, M.I. Fe modulates Cd-induced oxidative stress and the expression of stress responsive proteins in the nodules of Vigna radiata. Plant Growth Regul. 2012, 68, 421–433. [Google Scholar] [CrossRef]
- Mori, S. Iron acquisition by plants. Curr. Opin. Plant Biol. 1999, 2, 250–253. [Google Scholar] [CrossRef]
- Muneer, S.; Lee, B.R.; Kim, K.Y.; Park, S.H.; Zhang, Q.; Kim, T.H. Involvement of sulphur nutrition in modulating iron deficiency responses in photosynthetic organelles of oilseed rape (Brassica napus L.). Photosynth. Res. 2014, 119, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Römheld, V.; Kissel, M. Different strategies in higher plants in mobilization and uptake of iron. J. Plant Nutr. 1986, 9, 695–713. [Google Scholar] [CrossRef]
- Schaaf, G.; Ludewig, U.; Erenoglu, B.E.; Mori, S.; Kitahara, T.; von Wiren, N. ZmYS1 functions as a proton-coupled symporter for phytosiderophore- and nicotianamine-chelated metals. J. Biol. Chem. 2004, 279, 9091–9096. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Nakanishi, H.; Nishizawa, N.K. Characterizing the crucial components of iron homeostasis in the Maize Mutants ys1 and ys3. PLoS ONE 2013, 8, 62567. [Google Scholar] [CrossRef] [PubMed]
- Waters, B.M.; Blevins, D.G.; Eide, D.J. Characterization of FRO1, a pea ferric-chelate reductase involved in root iron acquisition. Plant Physiol. 2002, 129, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Connolly, E.L.; Campbell, N.H.; Grotz, N.; Prichard, C.L.; Guerinot, M.L. Overexpression of the FRO2 ferric chelate reductase confers tolerance to growth on low iron and uncovers posttranscriptional control. Plant Physiol. 2003, 133, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Zocchi, G. Metabolic changes in iron stressed dicotyledonous plants. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Barton, L.L., Abadía, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 359–370. [Google Scholar]
- Zuchi, S.; Cesco, S.; Varanini, Z.; Pinton, R.; Astolfi, S. Sulphur deprivation limits Fe-deficiency responses in tomato plants. Planta 2009, 230, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, R.; Brumbarova, T.; Bauer, P. Fitting into the harsh reality: Regulation of iron-deficiency responses in dicotyledonous plants. Mol. Plant 2009, 5, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.J.; Procter, C.M.; Connolly, E.L.; Guerinot, M.L. A ferric-chelate reductase for iron uptake from soils. Nature 1999, 397, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Lee, B.R.; Bae, D.W.; Kim, T.H. Changes in expression of proteins involved in alleviation of Fe-deficiency by sulfur nutrition in Brassica napus L. Acta Physiol. Plant. 2013, 35, 3037–3045. [Google Scholar] [CrossRef]
- Timperio, A.M.; D’Amici, G.M.; Barta, C.; Loreto, F.; Zolla, L. Proteomics, pigment composition, and organization of thylakoid membranes in iron-deficient spinach leaves. J. Exp. Bot. 2007, 58, 3695–3710. [Google Scholar] [CrossRef] [PubMed]
- Andaluz, S.; López-Millán, A.F.; Rivas, J.D.L.; Aro, E.M.; Abadía, J.; Abadía, A. Proteomic profiles of thylakoid membranes and changes in response to iron deficiency. Photosynth. Res. 2006, 89, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Briat, J.F.; Curie, C.; Gaymard, F. Iron utilization and metabolism in plants. Curr. Opin. Plant Biol. 2007, 10, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Szaciłowski, K.; Chmura, A.; Stasicka, Z. Interplay between iron complexes, nitric oxide and sulfur ligands: Structure, (photo) reactivity and biological importance. Coord. Chem. Rev. 2005, 249, 2408–2436. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Graziano, M.; Lamattina, L. Nitric oxide and iron in plants: An emerging and converging story. Trends Plant Sci. 2005, 10, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Ksouri, R.; Gharsalli, M.; Lachaâl, M. Differences in responses to iron deficiency between two legumes: Lentil (Lens culinaris) and chickpea (Cicer arietinum). J. Plant Physiol. 2005, 162, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Connolly, E.L. Iron uptake mechanisms in plants: Functions of the FRO family of ferric reductases. Plant Sci. 2009, 176, 709–714. [Google Scholar] [CrossRef]
- Andaluz, S.; Rodríguez-Celma, J.; Abadía, A.; Abadía, J.; López-Millán, A.F. Time course induction of several key enzymes in Medicago truncatula roots in response to Fe deficiency. Plant Physiol. Biochem. 2009, 47, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Jeong, B.R.; Kim, T.H.; Lee, J.H.; Soundararajan, P. Transcriptional and physiological changes in relation to Fe uptake under conditions of Fe-deficiency and Cd-toxicity in roots of Vigna radiata L. J. Plant Res. 2014, 127, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Choudhary, M.K.; Bhushan, D.; Chattopadhyay, A.; Chakraborty, S.; Datta, A.; Chakroborty, N. The Nuclear proteome of chick pea (Cicer arietinum L.) reveals predicted and un-expected proteins. J. Proteome Res. 2006, 5, 3301–3311. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Ishida, H.; Makino, A.; Mae, T. Fe2+−catalyzed site specific cleavage of the large subunit of ribulose 1,5-bisphosphatecarboxylase close to the active site. J. Biol. Chem. 2002, 277, 12382–12387. [Google Scholar] [CrossRef] [PubMed]
- Sandstrom, S.; Ivanov, A.G.; Park, Y.I.; Öquist, G.; Gustafsson, P. Iron stress responses in the cyanobacterium Synechococcus sp. PCC 7942. Physiol. Plant. 2002, 116, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, E.; Romera, F.J.; Cañete, M.; de la Huardia, M.I. Effects of heavy metals on both induction and function of root Fe(III) reductase in Fe-deficient cucumber (Cucumis sativus L.) plants. J. Exp. Bot. 1994, 45, 1893–1898. [Google Scholar] [CrossRef]
- Henriques, R.; Jásik, J.; Klein, M.; Martinoia, E.; Feller, U.; Schell, J.; Pais, M.S.; Koncz, C. Knock-out of Arabidopsis metal transporter gene IRT1 results in iron deficiency accompanied by cell differentiation defects. Plant Mol. Biol. 2002, 50, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Hahn, E.J.; Paek, K.Y. Effects of temperature on oxidative stress defense systems, lipid peroxidation and lipoxygenase activity in Phalaenopsis. Plant Physiol. Biochem. 2005, 43, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.R.; Jung, W.J.; Jin, Y.L.; Avice, J.C.; Ourry, A.; Kim, T.H. Water deficit-induced oxidative stress and the activation of antioxidative enzymes in white clover leaves. Biol. Plant. 2009, 53, 505–510. [Google Scholar] [CrossRef]
- Molassiotis, A.N.; Diamantidis, G.C.; Therios, I.N.; Tsirakoglou, V.; Dimassi, K.N. Oxidative stress, antioxidant activity and Fe(III)-chelate reductase activity of five Prunus rootstocks explants in response to Fe deficiency. Plant Growth Regul. 2005, 46, 69–78. [Google Scholar] [CrossRef]
- Ranieri, A.; Castagna, A.; Baldan, B.; Soldatini, G.F. Iron deficiency differently affects peroxidase isoforms in sunflower. J. Exp. Bot. 2001, 52, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef] [PubMed]
- Grill, C.J.; Cohick, W.S.; Sherman, A.R. Post pubertal development of the rat mammary gland is preserved during iron deficiency. J. Nutr. 2001, 131, 1444–1448. [Google Scholar] [PubMed]
- Bienfait, H.F.; van den Briel, W.; Mesland-Mul, N.T. Free space iron pools in roots: Generation and mobilization. Plant Physiol. 1985, 78, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principal of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes complex proteins and photosynthetic activity under different intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed]
- Sárvári, E.; Mihailova, G.; Solti, Á.; Keresztes, Á.; Velitchkov, M.; Georgieva, K. Comparison of thylakoid structure and organization in sun and shade Haberlea rhodopensis populations under desiccation and rehydration. J. Plant Physiol. 2014, 171, 1591–1600. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, W. Root-mediated ferric reduction: responses to iron deficiency, exogenously induced changes in hormonal balance and inhibition of protein synthesis. J. Exp. Bot. 1994, 45, 725–731. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muneer, S.; Jeong, B.R. Genotypic Variation under Fe Deficiency Results in Rapid Changes in Protein Expressions and Genes Involved in Fe Metabolism and Antioxidant Mechanisms in Tomato Seedlings (Solanum lycopersicum L.). Int. J. Mol. Sci. 2015, 16, 28022-28037. https://doi.org/10.3390/ijms161226086
Muneer S, Jeong BR. Genotypic Variation under Fe Deficiency Results in Rapid Changes in Protein Expressions and Genes Involved in Fe Metabolism and Antioxidant Mechanisms in Tomato Seedlings (Solanum lycopersicum L.). International Journal of Molecular Sciences. 2015; 16(12):28022-28037. https://doi.org/10.3390/ijms161226086
Chicago/Turabian StyleMuneer, Sowbiya, and Byoung Ryong Jeong. 2015. "Genotypic Variation under Fe Deficiency Results in Rapid Changes in Protein Expressions and Genes Involved in Fe Metabolism and Antioxidant Mechanisms in Tomato Seedlings (Solanum lycopersicum L.)" International Journal of Molecular Sciences 16, no. 12: 28022-28037. https://doi.org/10.3390/ijms161226086
APA StyleMuneer, S., & Jeong, B. R. (2015). Genotypic Variation under Fe Deficiency Results in Rapid Changes in Protein Expressions and Genes Involved in Fe Metabolism and Antioxidant Mechanisms in Tomato Seedlings (Solanum lycopersicum L.). International Journal of Molecular Sciences, 16(12), 28022-28037. https://doi.org/10.3390/ijms161226086