
The Importance of G Protein-Coupled Receptor Kinase 4 (GRK4) in Pathogenesis of Salt Sensitivity, Salt Sensitive Hypertension and Response to Antihypertensive Treatment


Abstract
:1. Introduction
2. Dopamine, G-Protein-Coupled Receptors and GRK4


3. The Role of GRK4 in the Pathogenesis of Salt Sensitivity in Animal Models
4. The Role of GRK4 Variants on Na Excretion in Humans
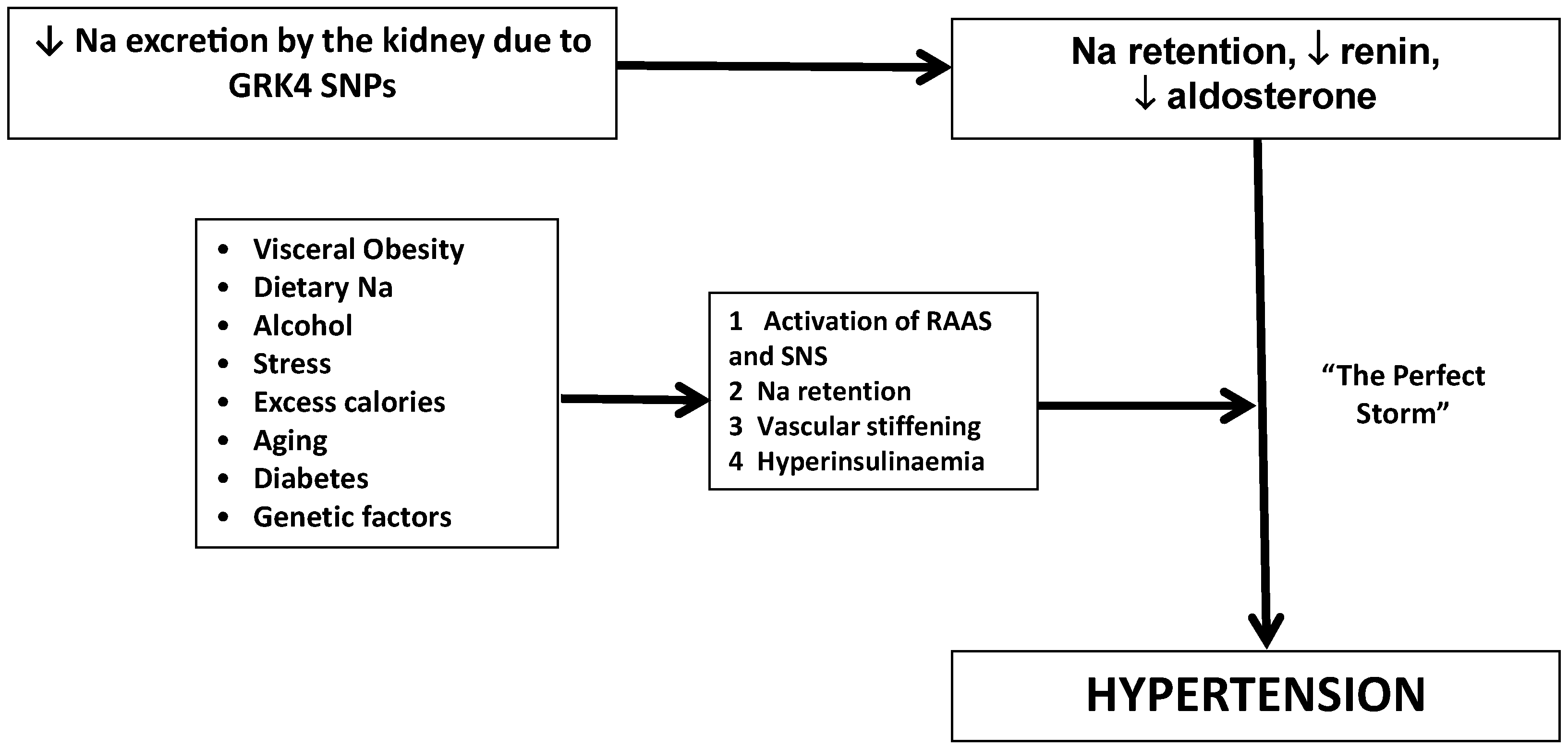
5. GRK4 Variants and Hypertension

6. GRK4 Variants and Response to Dietary and Pharmacological Intervention

{kind=link}
{kind=link}
{kind=link}
| GRK Variant | Parameter | Comment |
|---|---|---|
| p.Arg65Leu, p.Ala142Val, and p.Val486Ala | Hypertension in rats | p.Ala142Val shows greatest activity |
| p.Ala142Val | Enhanced activity of AT1 in smooth muscle | – |
| p.Arg65Leu | Impaired stress related Na excretion in normotensive blacks | – |
| p.Ala142Val | Impaired incremental Na excretion in healthy men, and lower aldosterone levels | p.Ala142Val significantly more common in Blacks compared to Whites |
| p.Arg65Leu, p.Ala142Val, and p.Val486Ala | Impaired Na excretion and higher BNP levels in healthy Japanese with two or more variants | – |
| p.Val486Ala | Association with hypertension in Australian population | – |
| p.Arg65Leu, p.Ala142Val, and p.Val486Ala | 94.4% predictive of salt sensitive renin hypertension in Japanese population | p.Ala142Val 78.4% predictive |
| p.Arg65Leu, p.Ala142Val | BP response to Na restriction in black South Africans with hypertension | – |
| p.142Ala genotype | In AASK males less likely to respond to metoprolol | Especially if co-inherited the p.65Leu variant |
| 65Leu/142Val haplotype | In two major hypertension studies predicted a reduced response to atenolol and increased cardiovascular outcomes | – |
7. Conclusions
Author Contributions
Conflicts of Interest
References
- Weinberger, M.H. Salt sensitivity of blood pressure in humans. Hypertension 1996, 27, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Luft, F.C.; Grim, C.E.; Higgins, J.T., Jr.; Weinberger, M.H. Differences in response to sodium administration in normotensive white and black subjects. J. Lab. Clin. Med. 1977, 90, 555–562. [Google Scholar] [PubMed]
- Mensah, G.A.; Mokdad, A.H.; Ford, E.S.; Greenlund, K.J.; Croft, J.B. State of disparities in cardiovascular health in the United States. Circulation 2005, 1111, 1233–1241. [Google Scholar] [CrossRef]
- Redmond, N.; Baer, H.J.; Hicks, L.S. Health behaviors and racial disparity in blood pressure control in the National Health and Nutrition Examination Survey. Hypertension 2011, 57, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Eckert, G.J.; Hannon, T.S.; Liu, H.; Pratt, L.M.; Wagner, M.A.; DiMeglio, L.A.; Jung, J.; Pratt, J.H. Racial differences in sensitivity of blood pressure to aldosterone. Hypertension 2014, 63, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Rayner, B.L.; Meyers, J.E.; Opie, L.H.; Trinder, Y.A.; Davidson, J.S. Screening for primary aldosteronism—Normal ranges for aldosterone and renin in three South African groups. S. Afr. Med. J. 2001, 91, 594–599. [Google Scholar] [PubMed]
- Bailey, M.A.; Shirley, D.G.; Unwin, R.J. Renal physiology. In Comprehensive Clinical Nephrology, 5th ed.; Johnson, R.J., Feehally, J., Floege, J., Eds.; Elsevier, Sanders: Philadelphia, PA, USA, 2014; Volume Chapter 2, pp. 14–27. [Google Scholar]
- Gross, P.; Heduschka, P. Inherited disorders of sodium and water metabolism. In Comprehensive Clinical Nephrology, 5th ed.; Johnson, R.J., Feehally, J., Floege, J., Eds.; Elsevier, Sanders: Philadelphia, PA, USA, 2014; Volume Chapter 49, pp. 579–589. [Google Scholar]
- Bochud, M.; Staessen, J.A.; Maillard, M.; Mazeko, M.J.; Kuznetsova, T.; Woodiwiss, A.; Richart, T.; Norton, G.; Thijs, L.; Elston, R.; et al. Ethnic differences in proximal and distal tubular sodium reabsorbtion are heritable in black and white populations. J. Hypertens. 2009, 27, 606–612. [Google Scholar]
- Aperia, A.C. Intrarenal dopamine: A key signal in the interactive regulation of sodium metabolism. Annu. Rev. Physiol. 2000, 62, 621–647. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Eisner, G.M.; Felder, R.A. Dopamine and the kidney: A role in hypertension? Curr. Opin. Nephrol. Hypertens. 2003, 12, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Eisner, G.M.; Felder, R.A. Role of dopamine receptors in the kidney in the regulation of blood pressure. Curr. Opin. Nephrol. Hypertens. 2002, 11, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Sanada, H.; Yatabe, J.; Midorikawa, S.; Katoh, T.; Hashimoto, S.; Watanabe, T.; Xu, J.; Luo, Y.; Wang, X.; Zeng, C.; et al. Amelioration of genetic hypertension by suppression of renal G protein-coupled receptor kinase type 4 expression. Hypertension 2006, 47, 1131–1139. [Google Scholar]
- Felder, R.; Sanada, H.; Xu, J.; Wang, Z.; Watanabe, H.; Asico, L.D.; Wang, W.; Zheng, S.; Yamaguchi, I.; Williams, S.M.; et al. G protein-coupled receptor kinase 4 variants in human essential hypertension. Proc. Natl. Acad. Sci. USA 2002, 99, 3872–3877. [Google Scholar]
- Chen, K.; Fu, C.; Chen, C.; Liu, L.; Ren, H.; Han, Y.; Yang, J.; He, D.; Zhou, L.; Yang, Z.; et al. Role of GRK4 in the Regulation of Arterial AT1 Receptor in Hypertension. Hypertension 2014, 63, 289–296. [Google Scholar]
- Zhu, H.; Lu, Y.; Wang, X.; Snieder, H.; Treiber, F.A.; Harshfield, G.A.; Dong, Y. The G protein-coupled receptor kinase 4 gene modulates stress-induced sodium excretion in black normotensive adolescents. Pediatr. Res. 2006, 60, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Rayner, B.; Musekiwa, A.; Lombard, C.; Ramesar, R. The A142V polymorphism of the G protein-coupled receptor kinase 4 gene predicts natriuretic response to saline challenge in young normotensive lean Black and White South African men. Nephrol. Rev. 2011, 3, e9. [Google Scholar] [CrossRef]
- Yatabe, Y.; Yatabe, M.S.; Yoneda, M.; Felder, R.A.; Jose, P.A.; Sanada, H. Hypertension-related gene polymorphisms of G-protein-coupled receptor kinase 4 are associated with NT-proBNP concentration in normotensive healthy adults. Int. J. Hypertens. 2012, 2012. [Google Scholar] [CrossRef]
- Sanada, H.; Yatabe, J.; Midorikawa, S.; Hashimoto, S.; Watanabe, T.; Moore, J.H.; Ritchie, M.D.; Williams, S.M.; Pezzullo, J.C.; Sasaki, M.; et al. Single-nucleotide polymorphisms for diagnosis of salt-sensitive hypertension. Clin. Chem. 2006, 52, 352–360. [Google Scholar]
- Speirs, H.J.L.; Katyk, K.; Kumar, N.N.; Benjafield, A.V.; Wang, W.Y.S.; Morris, B.J. Association of G-protein-coupled receptor kinase 4 haplotypes, but not HSD3B1 or PTP1B polymorphisms, with essential hypertension. J. Hypertens. 2004, 22, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Lu, Y.; Wang, X.; Treiber, F.A.; Harshfield, G.A.; Snieder, H.; Dong, Y. The G protein-coupled receptor kinase 4 gene affects blood pressure in young normotensive twins. Am. J. Hypertens. 2006, 19, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Rayner, B.; Ramesar, R.; Steyn, K.; Levitt, N.; Lombard, C.; Charlton, K. G-protein-coupled receptor kinase 4 (GRK-4) polymorphisms predict blood pressure response to dietary modification in black patients with mild to moderate hypertension. J. Hum. Hypertens. 2011, 26, 334–239. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, V.; O’Connor, D.T.; Brophy, V.H.; Schork, N.J.; Richard, E.; Salem, R.M.; Nievergelt, C.M.; Bakris, G.L.; Middleton, J.P.; Norris, K.C.; et al. G-Protein-coupled receptor kinase 4 polymorphisms and blood pressure response to metoprolol among african americans: Sex-specificity and interactions. Am. J. Hypertens. 2009, 22, 332–338. [Google Scholar]
- Vandell, A.G.; Lobmeyer, M.T.; Gawronski, B.E.; Langaee, T.Y.; Gong, Y.; Gums, J.G.; Beitelshees, A.L.; Turner, S.T.; Chapman, A.B.; Cooper-Dehoff, R.M.; et al. G protein receptor kinase 4 polymorphisms: β-Blocker Pharmacogenetics and treatment-related outcomes in Hypertension. Hypertension 2012, 60, 957–964. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rayner, B.; Ramesar, R. The Importance of G Protein-Coupled Receptor Kinase 4 (GRK4) in Pathogenesis of Salt Sensitivity, Salt Sensitive Hypertension and Response to Antihypertensive Treatment. Int. J. Mol. Sci. 2015, 16, 5741-5749. https://doi.org/10.3390/ijms16035741
Rayner B, Ramesar R. The Importance of G Protein-Coupled Receptor Kinase 4 (GRK4) in Pathogenesis of Salt Sensitivity, Salt Sensitive Hypertension and Response to Antihypertensive Treatment. International Journal of Molecular Sciences. 2015; 16(3):5741-5749. https://doi.org/10.3390/ijms16035741
Chicago/Turabian StyleRayner, Brian, and Raj Ramesar. 2015. "The Importance of G Protein-Coupled Receptor Kinase 4 (GRK4) in Pathogenesis of Salt Sensitivity, Salt Sensitive Hypertension and Response to Antihypertensive Treatment" International Journal of Molecular Sciences 16, no. 3: 5741-5749. https://doi.org/10.3390/ijms16035741
APA StyleRayner, B., & Ramesar, R. (2015). The Importance of G Protein-Coupled Receptor Kinase 4 (GRK4) in Pathogenesis of Salt Sensitivity, Salt Sensitive Hypertension and Response to Antihypertensive Treatment. International Journal of Molecular Sciences, 16(3), 5741-5749. https://doi.org/10.3390/ijms16035741