
β-Hydroxybutyric Sodium Salt Inhibition of Growth Hormone and Prolactin Secretion via the cAMP/PKA/CREB and AMPK Signaling Pathways in Dairy Cow Anterior Pituitary Cells

Abstract
:1. Introduction
2. Results
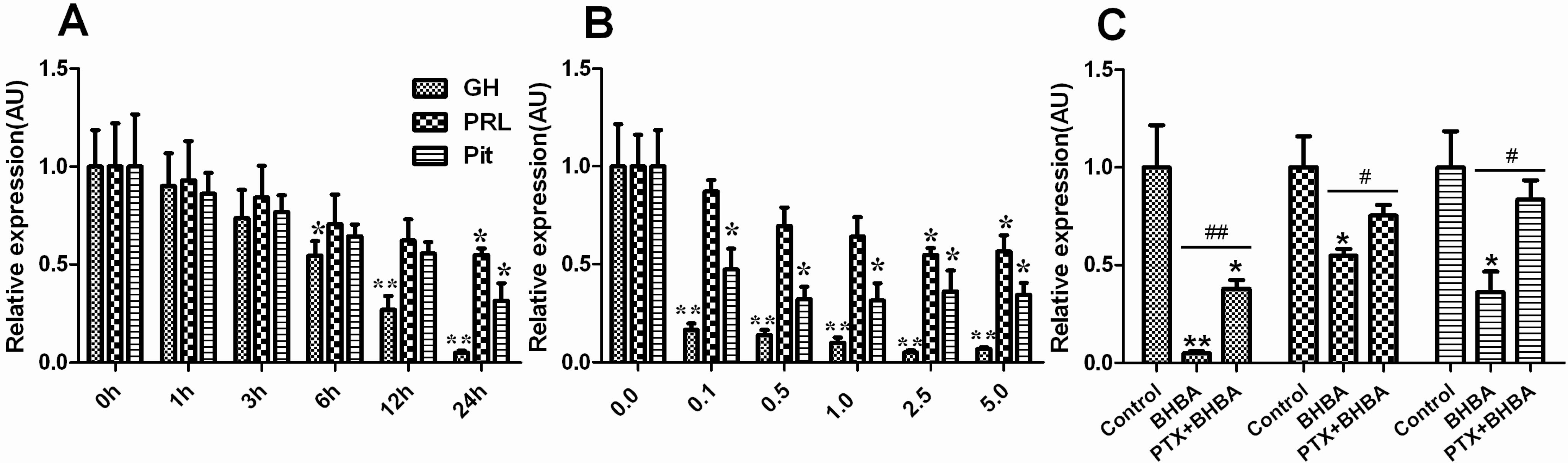
2.1. Effect of BHBA on mRNA Levels of GH, PRL and Pit-1 in DCAPCs

2.2. Effect of BHBA on GH and PRL Secretion in DCAPCs

2.3. Effect of BHBA on Intracellular cAMP Concentration

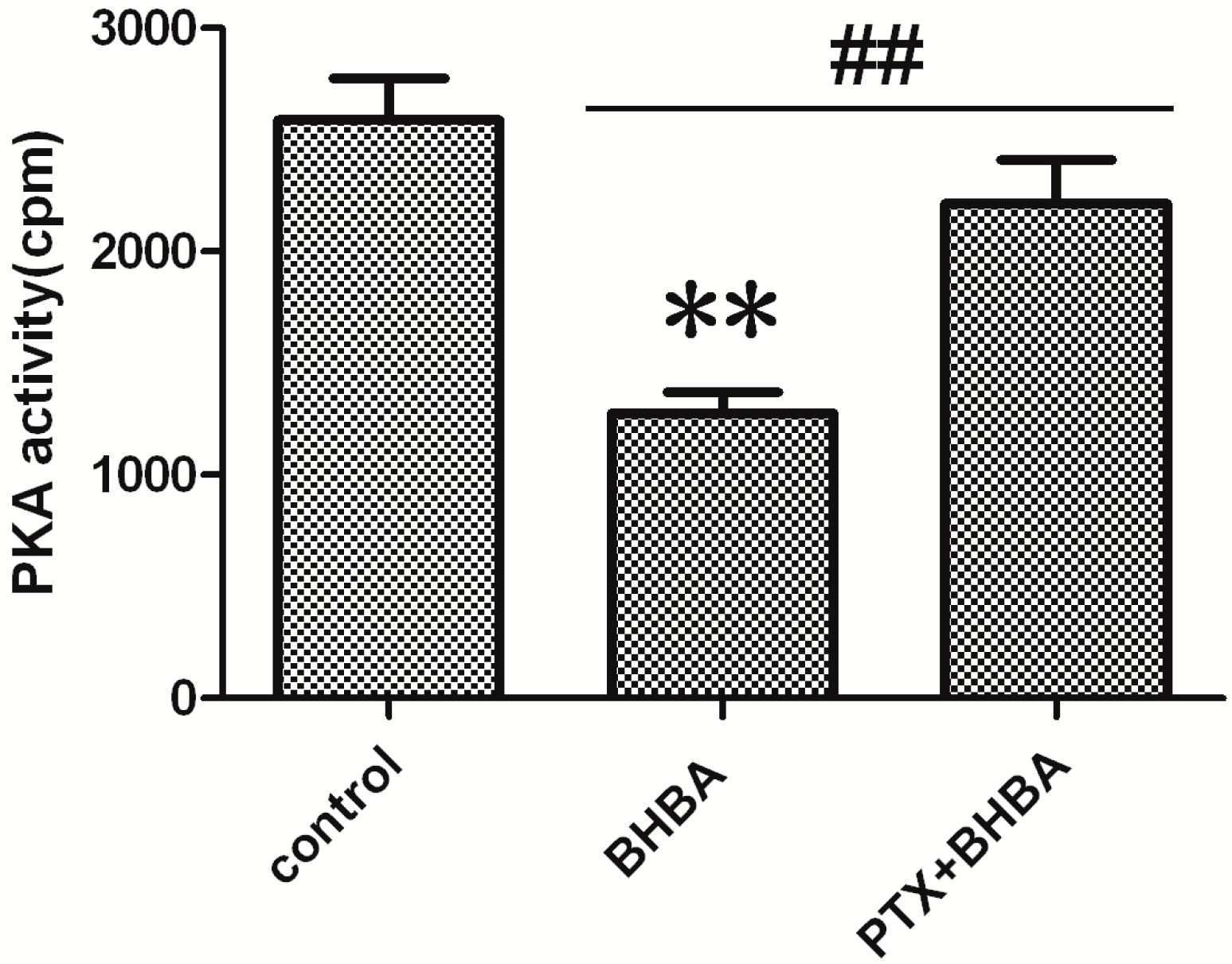
2.4. Effect of BHBA on PKA Activity

2.5. Effect of BHBA on CREB Phosphorylation

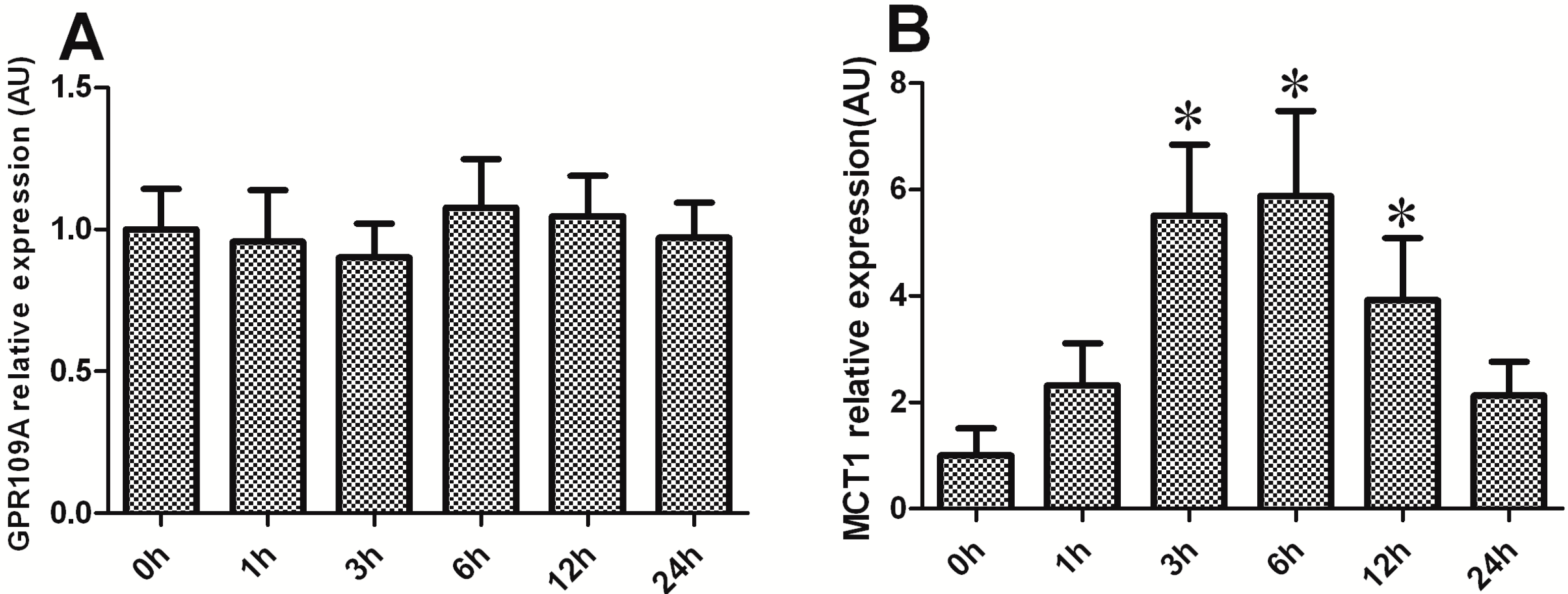
2.6. Effect of BHBA on the mRNA Levels of GPR109A and MCT1 in DCAPCs

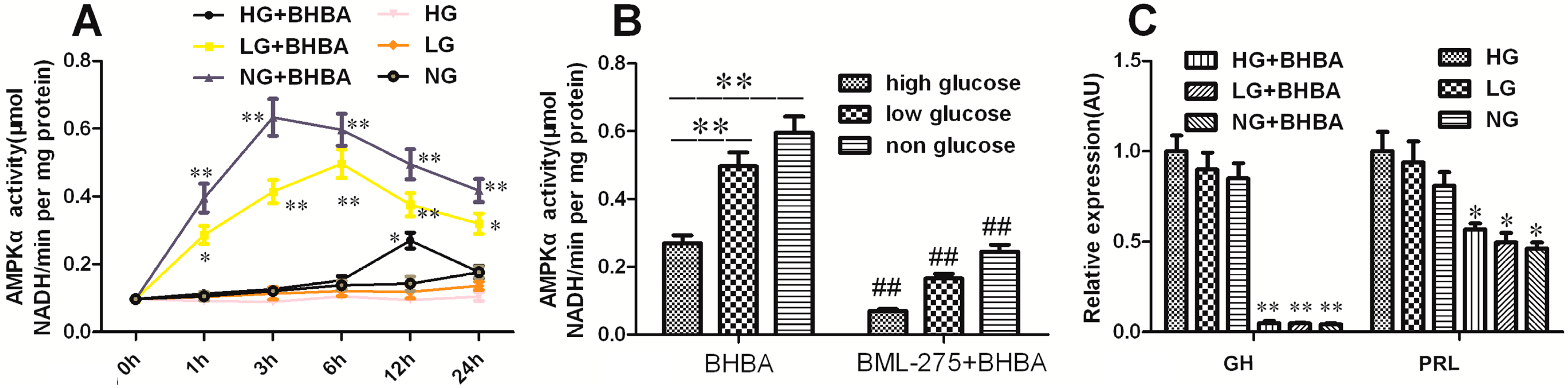
2.7. The Role of AMPK-α in BHBA-Regulated GH and PRL Transcription and Secretion


3. Discussion

4. Materials and Methods
4.1. Isolation and Culture of DCAPCs
4.2. RNA Extraction, Reverse Transcriptase, and Quantitative Real-Time PCR Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences | Length (bp) |
|---|---|---|
| GAPDH | (F) 5'-TGCCCAGAATATCATCCC-3' | 134 |
| (R) 5'-AGGTCAGATCCACAACAG-3' | ||
| GH | (F) 5'-AGATCCTCAAGCAGACCTA-3' | 121 |
| (R) 5'-AGGTACGTCTCCGTCTTA-3' | ||
| PRL | (F) 5'-TATGAAAGGAGCCCCAGATG-3' | 137 |
| (R) 5'-CACACAGGGTAGGGCTCAGT-3' | ||
| Pit-1 | (F) 5'-TTCTGCAACTCTGCCTCTGA-3' | 148 |
| (R) 5'-CCATAGGTCGATGACTGGT-3' | ||
| GPR109A | (F) 5'-ACATCACCCTCAGCTTCACC-3' | 146 |
| (R) 5'-GCGGTTGTTATCCGACTCAT-3' | ||
| MCT1 | (F) 5'-GGAGTCATTGGAGGTCTTGG-3' | 137 |
| (R) 5'-GCCAGGGTAGAGAGGAACAC-3' |
4.3. Measurement of GH and PRL Concentration in the Supernatant
4.4. Measurement of Intracellular cAMP Concentration
4.5. Measurement of PKA Activity
4.6. Western Blotting Analysis of Phosphorylated CREB
4.7. Measurement of AMPK-α Activity
4.8. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shi, X.; Li, D.; Deng, Q.; Li, Y.; Sun, G.; Yuan, X.; Song, Y.; Wang, Z.; Li, X.; Li, X.; et al. NEFAs activate the oxidative stress-mediated NF-κB signaling pathway to induce inflammatory response in calf hepatocytes. J. Steroid Biochem. Mol. Biol. 2015, 14, 103–112. [Google Scholar] [CrossRef]
- Ooi, G.T.; Tawadros, N.; Escalona, R.M. Pituitary cell lines and their endocrine applications. Mol. Cell Biol. 2004, 228, 1–21. [Google Scholar]
- Meier, S.; Gore, P.J.; Barnett, C.M.; Cursons, R.T.; Phipps, D.E.; Watkins, K.A.; Verkerk, G.A. Metabolic adaptations associated with irreversible glucose loss are different to those observed during under-nutrition. Domest. Anim. Endocrinol. 2008, 34, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Laeger, T.; Metges, C.C.; Kuhla, B. Role of beta-hydroxybutyric acid in the central regulation of energy balance. Appetite 2010, 54, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Taggart, A.K.; Kero, J.; Gan, X.; Cai, T.Q.; Cheng, K.; Ippolito, M.; Ren, N.; Kaplan, R.; Wu, K.; Wu, T.J.; et al. (d)-β-Hydroxybutyrate inhibits adipocyte lipolysis via the nicotinic acid receptor PUMA-G. J. Biol. Chem. 2005, 280, 26649–26652. [Google Scholar] [CrossRef]
- Titgemeyer, E.C.; Mamedova, L.K.; Spivey, K.S.; Farney, J.K.; Bradford, B.J. An unusual distribution of the niacin receptor in cattle. J. Dairy Sci. 2011, 94, 2177–2187. [Google Scholar] [CrossRef]
- Wang, J.F.; Fu, S.P.; Li, S.N.; Hu, Z.M.; Xue, W.J.; Li, Z.Q.; Huang, B.X.; Lv, Q.K.; Liu, J.X.; Wang, W. Short-chain fatty acids inhibit growth hormone and prolactin gene transcription via cAMP/PKA/CREB signaling pathway in dairy cow anterior pituitary cells. Int. J. Mol. Sci. 2013, 14, 21474–21488. [Google Scholar] [CrossRef] [PubMed]
- Laeger, T.; Pöhland, R.; Metges, C.C.; Kuhla, B. The ketone body β-hydroxybutyric acid influences agouti-related peptide expression via AMP-activated protein kinase in hypothalamic GT1–7 cells. J. Endocrinol. 2012, 213, 193–203. [Google Scholar] [CrossRef]
- Pelletier, A.; Coderre, L. Ketone bodies alter dinitrophenol-induced glucose uptake through AMPK inhibition and oxidative stress generation in adult cardiomyocytes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1325–E1332. [Google Scholar] [CrossRef] [PubMed]
- Tulipano, G.; Giovannini, M.; Spinello, M.; Sibilia, V.; Giustina, A.; Cocchi, D. AMP-activated protein kinase regulates normal rat somatotroph cell function and growth of rat pituitary adenomatous cells. Pituitary 2011, 14, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Capuco, A.V.; Wood, D.L.; Baldwin, R.; McLeod, K.; Paape, M.J. Mammary cell number, proliferation, and apoptosis during a bovine lactation: Relation to milk production and effect of Bst. J. Dairy Sci. 2001, 84, 2177–2187. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, B.; Baracos, V.E.; Kennelly, J.J. Effects of bovine somatotropin on beta-casein mRNA levels in mammary tissue of lactating cows. J. Dairy Sci. 2005, 88, 2806–2812. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, S.; Sakamoto, K.; Komatsu, T.; Hagino, A.; Katoh, K.; Obara, Y. Growth hormone and lactogenic hormones can reduce the leptin mRNA expression in bovine mammary epithelial cells. Domest. Anim. Endocrinol. 2006, 31, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev. 2000, 4, 1523–1631. [Google Scholar]
- Trott, J.F.; Schennink, A.; Petrie, W.K.; Manjarin, R.; VanKlompenberg, M.K.; Hovey, R.C. Triennial Lactation Symposium: Prolactin: The multifaceted potentiator of mammary growth and function. J. Anim. Sci. 2012, 95, 1674–1686. [Google Scholar] [CrossRef]
- Plaisance, E.P.; Lukasova, M.; Offermanns, S.; Zhang, Y.; Cao, G.; Judd, R.L. Niacin stimulates adiponectin secretion through the GPR109A receptor. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E549–E558. [Google Scholar] [CrossRef] [PubMed]
- Tunaru, S.; Kero, J.; Schaub, A.; Wufka, C.; Blaukat, A.; Pfeffer, K.; Offermanns, S. Puma-G and HM74 are receptors for nicotinic acid and mediate its anti-lipolytic effect. Nat. Med. 2003, 9, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Kamohara, M.; Takasaki, J.; Matsumoto, S.; Saito, T.; Ohishi, T.; Hiyama, H.; Matsuo, A.; Matsushime, H.; Furuichi, K. Molecular identification of nicotinic acid receptor. Biochem. Biophys. Res. Commun. 2003, 303, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Schmidt, R.J.; Foxworthy, P.; Emkey, R.; Oler, J.K.; Larg, T.H.; Wang, H.; Su, E.W.; Mosior, M.K.; Eacho, P.I.; et al. Niacin mediates lipolysis in adipose tissue through its G-protein coupled receptor HM74A. Biochem. Biophys. Res. Commun. 2005, 334, 729–732. [Google Scholar] [CrossRef]
- Nelson, C.; Albert, V.R.; Elsholtz, H.P.; Lu, L.I.; Rosenfeld, M.G. Activation of cell-specific expression of rat growth hormone and prolactin genes by a common transcription factor. Science 1988, 239, 1400–1405. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Pacheco, A.; Palomino, T.; Aranda, A. Negative regulation of expression of the pituitary-specific transcription factor GHF-1/Pit-1 by thyroid hormones through interference with promoter enhancer elements. Mol. Cell. Biol. 1995, 15, 6322–6330. [Google Scholar] [PubMed]
- Melmed, S. The Pituitary, 3rd ed.; Academic Press: London, UK, 2011; pp. 83–166. [Google Scholar]
- Chai, J.T.; Digby, J.E.; Choudhury, R.P. GPR109A and vascular inflammation. Curr. Atheroscler. Rep. 2013, 15, 325. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of GPR109A, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef]
- Kostylina, G.; Simon, D.; Fey, M.F.; Yousefi, S.; Simon, H.U. Neutrophil apoptosis mediated by nicotinic acid receptors (GPR109A). Cell Death Differ. 2008, 15, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Dowden, D.R.; Jacobson, D.R. Inhibition of appetite in dairy cattle by certain intermediate metabolities. Nature 1960, 188, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Zarrin, M.; de Matteis, L.; Vernay, M.C.; Wellnitz, O.; van Dorland, H.A.; Bruckmaier, R.M. Long-term elevation of β-hydroxybutyrate in dairy cows through infusion: Effects on feed intake, milk production, and metabolism. J. Dairy Sci. 2013, 96, 2960–2972. [Google Scholar] [CrossRef] [PubMed]
- Gwinn, D.M.; Shackelford, D.B.; Egan, D.F.; Mihaylova, M.M.; Mery, A.; Vasquez, D.S.; Turk, B.E.; Shaw, R.J. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell 2008, 30, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Hong, Y.H.; Shen, X.Q.; Frankowski, C.; Camp, H.S.; Leff, T. Regulation of transcription by AMP-activated protein kinase: Phosphorylation of p300 blocks its interaction with nuclear receptors. J. Biol. Chem. 2001, 276, 38341–38344. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; Yang, W.; Xiao, C.; Fu, S.; Deng, Q.; Ding, H.; Wang, Z.; Liu, G.; Li, X. SREBP-1c overexpression induces triglycerides accumulation through increasing lipid synthesis and decreasing lipid oxidation and VLDL assembly in bovine hepatocytes. J. Steroid Biochem. Mol. Biol. 2014, 143, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.E.; Hashimoto, Y.; Zanger, K.; Wondisford, F.; Radovick, S. CREB-independent regulation by CBP is a novel mechanism of human growth hormone gene expression. J. Clin. Investig. 1999, 104, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Lundblad, J.R.; Kwok, R.P.S.; Laurance, M.E.; Harter, M.L.; Goodman, R.H. Adenoviral E1A-associated p300 as a functional homologue of the transcriptional co-activator CBP. Nature 1995, 374, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Ouyang, H.; Zhu, T.; Lindvall, C.; Wang, Y.; Zhang, X.; Yang, Q.; Bennett, C.; Harada, Y.; Stankunas, K.; et al. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005, 1, 15–25. [Google Scholar] [CrossRef]
- Hung, C.M.; Garcia-Haro, L.; Sparks, C.A.; Guertin, D.A. mTOR-dependent cell survival mechanisms. Cold Spring Harb. Perspect. Biol. 2012, 4, a008771. [Google Scholar] [PubMed]
- Wang, J.F.; Fu, S.P.; Li, S.N.; Yang, Z.Q.; Xue, W.J.; Li, Z.Q.; Wang, W.; Liu, J.X. Establishment and characterization of dairy cow growth hormone secreting anterior pituitary cell model. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Bruckener, K.E.; el Baya, A.; Galla, H.J.; Schmidt, M.A. Permeabilization in a cerebral endothelial barrier model by pertussis toxin involves the PKC effector pathway and is abolished by elevated levels of cAMP. J. Cell Sci. 2003, 116, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Mangmool, S.; Kurose, H. Gi/o protein-dependent and -independent actions of pertussis toxin (PTX). Toxins 2011, 3, 884–899. [Google Scholar] [CrossRef] [PubMed]
- Laporta, J.; Driver, A.; Khatib, H. Short communication: Expression and alternative splicing of POU1F1 pathway genes in preimplantation bovine embryos. J. Dairy Sci. 2011, 94, 4220–4223. [Google Scholar] [CrossRef] [PubMed]
- Kirat, D.; Kato, S. Monocarboxylate transporter 1 (MCT1) mediates transport of short-chain fatty acids in bovine caecum. Exp. Physiol. 2006, 91, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Chen, H.; Lei, L.; Liu, J.; Guan, Y.; Liu, Z.; Zhang, L.; Yang, W.; Zhao, C.; et al. Non-esterified fatty acids activate the AMP-activated protein kinase signaling pathway to regulate lipid metabolism in bovine hepatocytes. Cell Biochem. Biophys. 2013, 67, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.-P.; Wang, W.; Liu, B.-R.; Yang, H.-M.; Ji, H.; Yang, Z.-Q.; Guo, B.; Liu, J.-X.; Wang, J.-F. β-Hydroxybutyric Sodium Salt Inhibition of Growth Hormone and Prolactin Secretion via the cAMP/PKA/CREB and AMPK Signaling Pathways in Dairy Cow Anterior Pituitary Cells. Int. J. Mol. Sci. 2015, 16, 4265-4280. https://doi.org/10.3390/ijms16024265
Fu S-P, Wang W, Liu B-R, Yang H-M, Ji H, Yang Z-Q, Guo B, Liu J-X, Wang J-F. β-Hydroxybutyric Sodium Salt Inhibition of Growth Hormone and Prolactin Secretion via the cAMP/PKA/CREB and AMPK Signaling Pathways in Dairy Cow Anterior Pituitary Cells. International Journal of Molecular Sciences. 2015; 16(2):4265-4280. https://doi.org/10.3390/ijms16024265
Chicago/Turabian StyleFu, Shou-Peng, Wei Wang, Bing-Run Liu, Huan-Min Yang, Hong Ji, Zhan-Qing Yang, Bin Guo, Ju-Xiong Liu, and Jian-Fa Wang. 2015. "β-Hydroxybutyric Sodium Salt Inhibition of Growth Hormone and Prolactin Secretion via the cAMP/PKA/CREB and AMPK Signaling Pathways in Dairy Cow Anterior Pituitary Cells" International Journal of Molecular Sciences 16, no. 2: 4265-4280. https://doi.org/10.3390/ijms16024265
APA StyleFu, S. -P., Wang, W., Liu, B. -R., Yang, H. -M., Ji, H., Yang, Z. -Q., Guo, B., Liu, J. -X., & Wang, J. -F. (2015). β-Hydroxybutyric Sodium Salt Inhibition of Growth Hormone and Prolactin Secretion via the cAMP/PKA/CREB and AMPK Signaling Pathways in Dairy Cow Anterior Pituitary Cells. International Journal of Molecular Sciences, 16(2), 4265-4280. https://doi.org/10.3390/ijms16024265