
4,4'-Diisothiocyanostilbene-2,2'-disulfonic Acid (DIDS) Ameliorates Ischemia-Hypoxia-Induced White Matter Damage in Neonatal Rats through Inhibition of the Voltage-Gated Chloride Channel ClC-2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
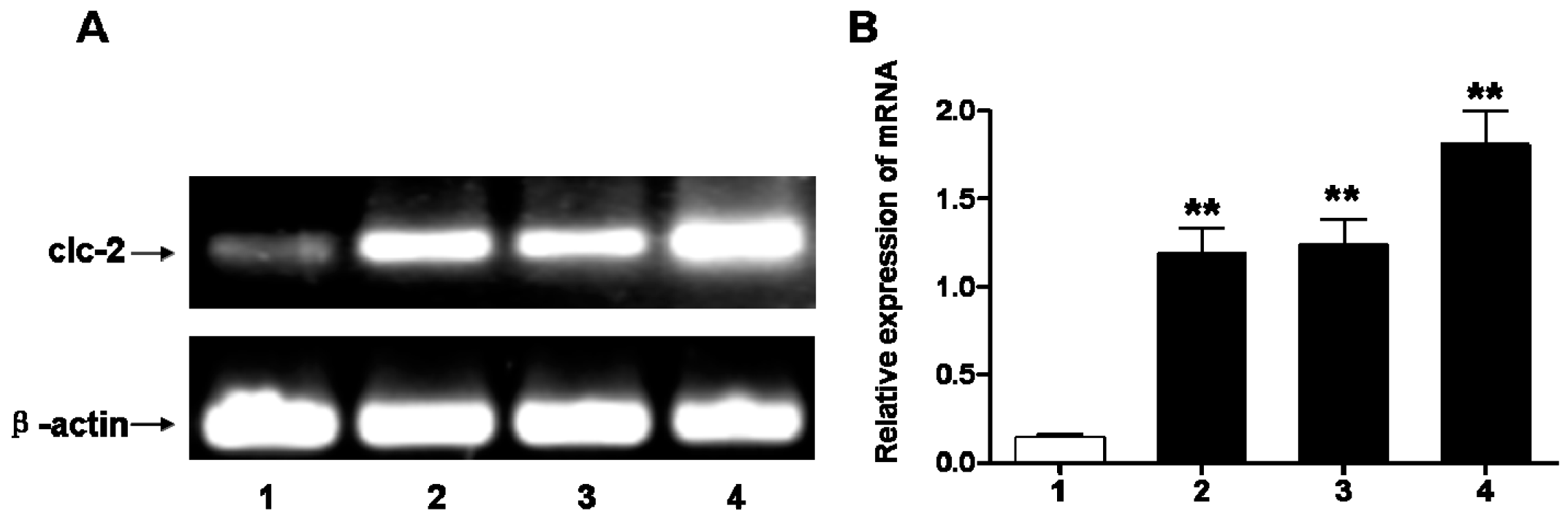
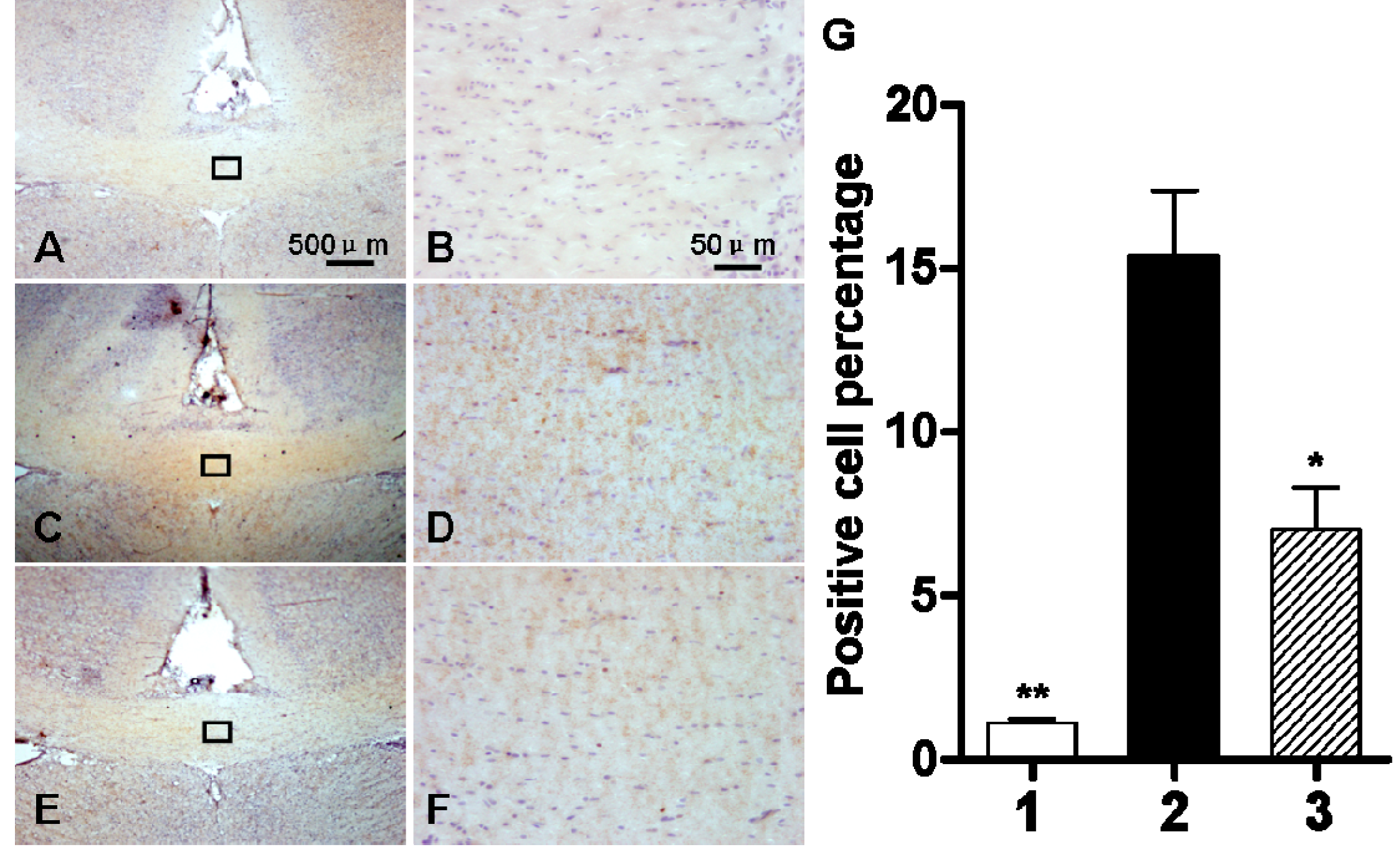
2.1. Chronic Ischemia and Hypoxia Induced ClC-2 Expression in the Cerebral White Matter



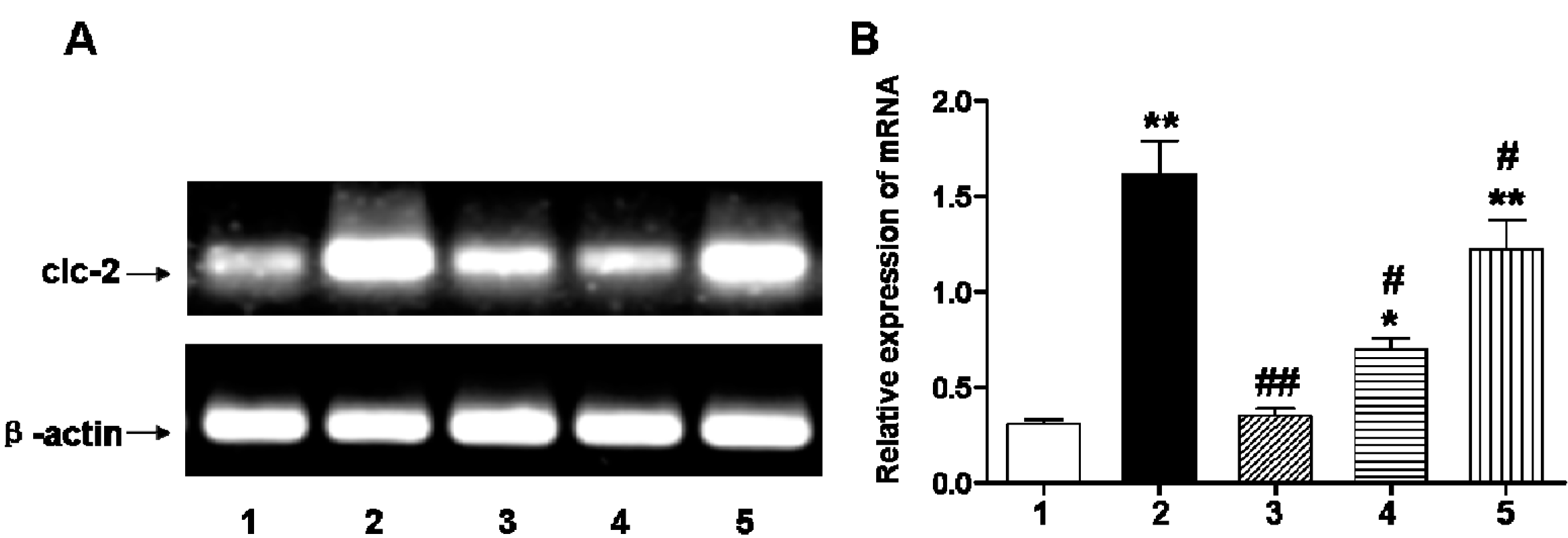
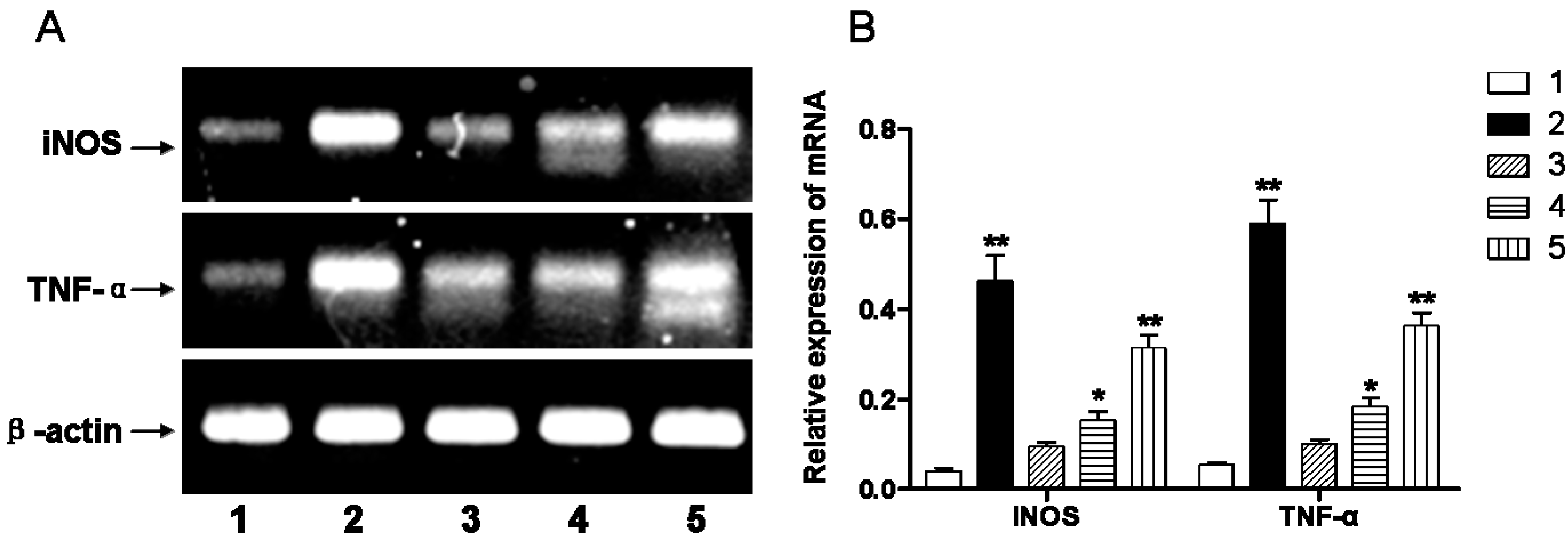
2.2. Early DIDS Administration during Hypoxic-Ischemic Injury Can Reduce the Concentration of Reactive Oxygen Species (ROS) and Inflammation


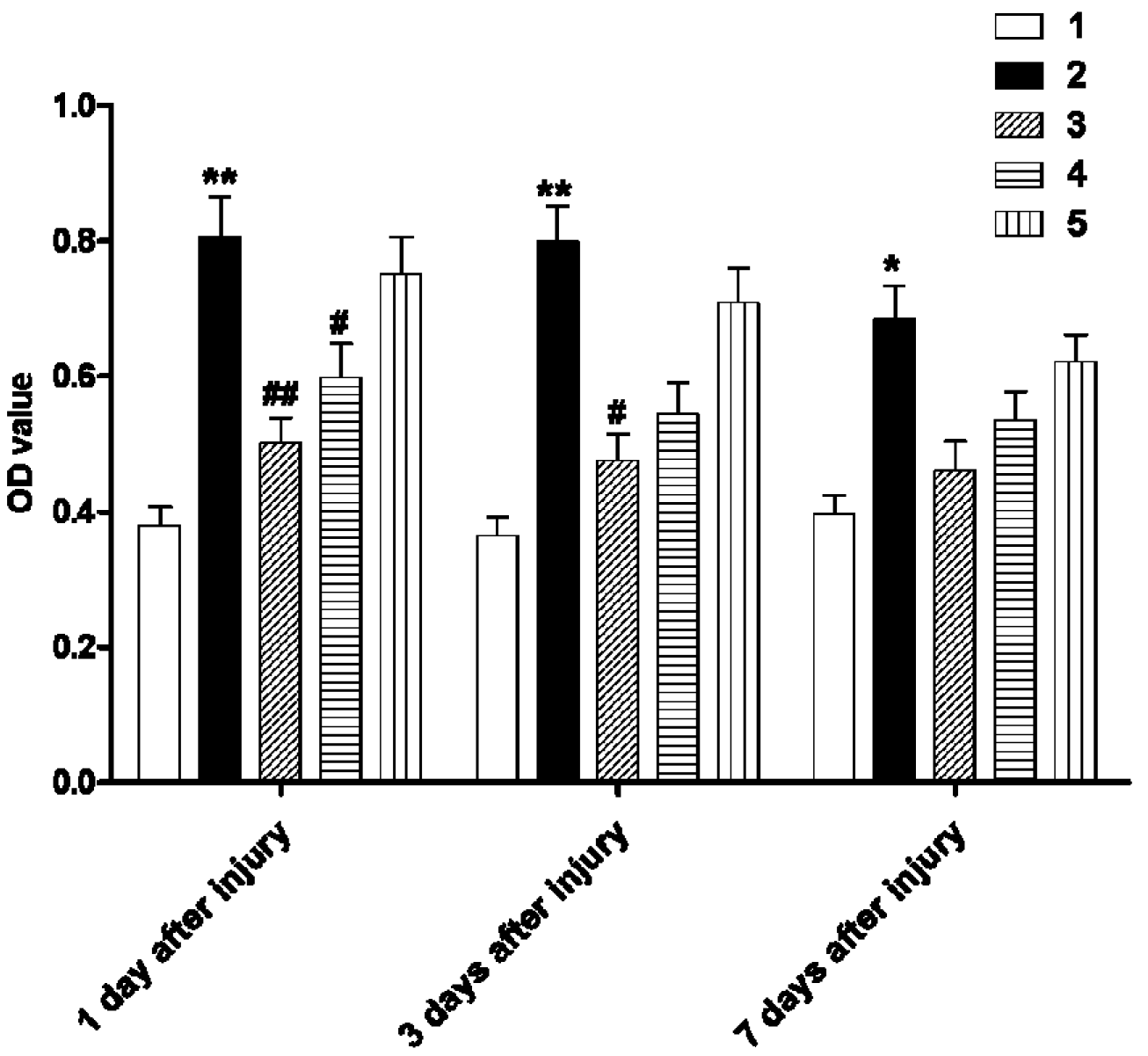
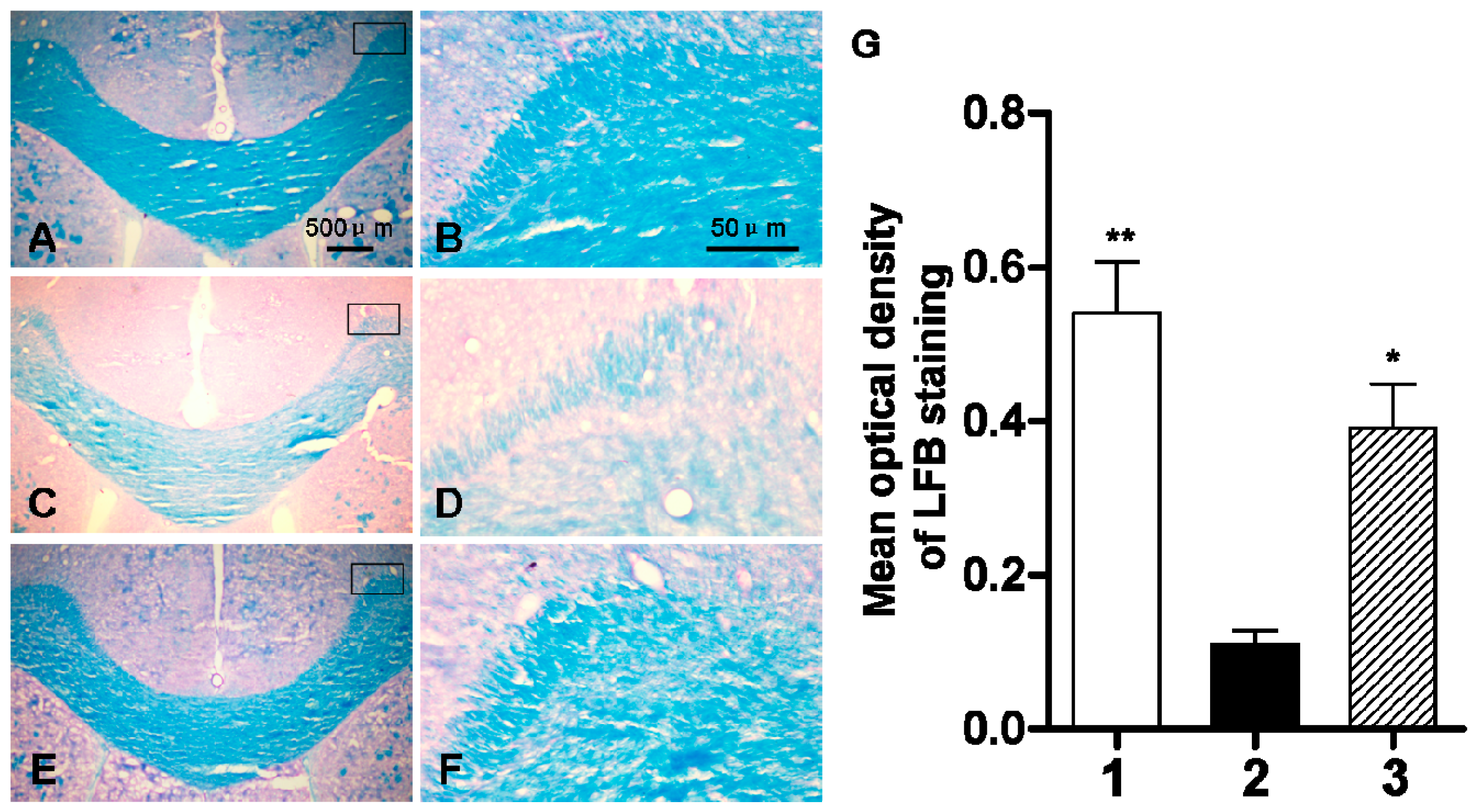
2.3. Early Administration of DIDS Reduced Demyelination


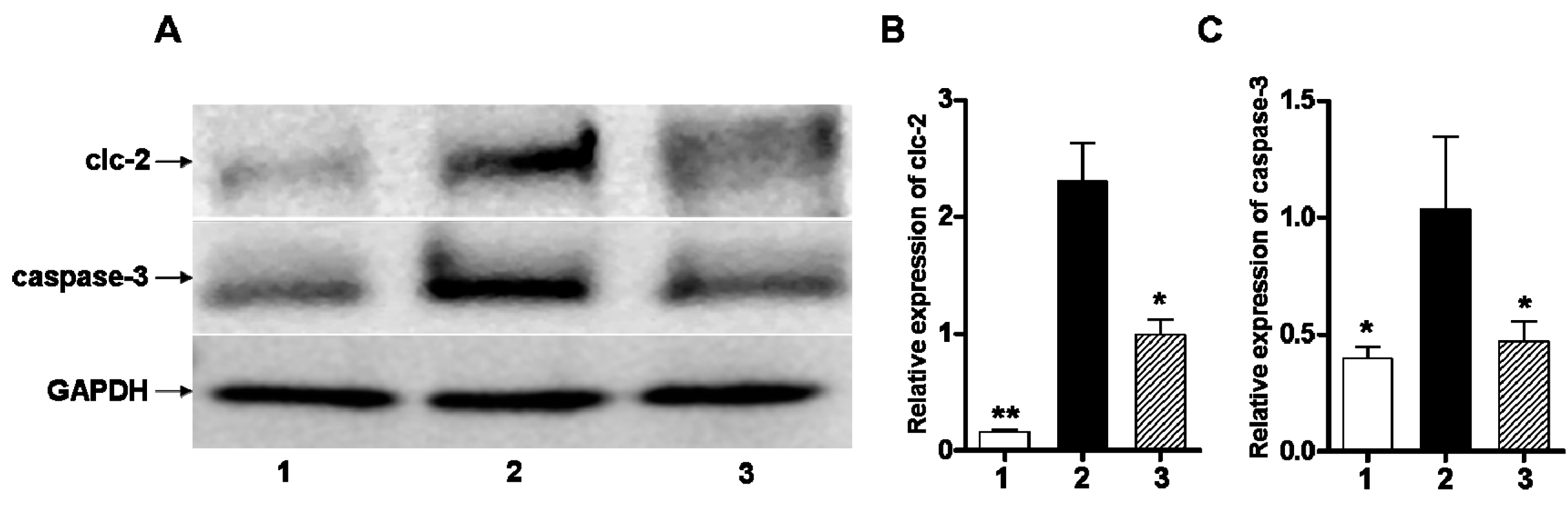
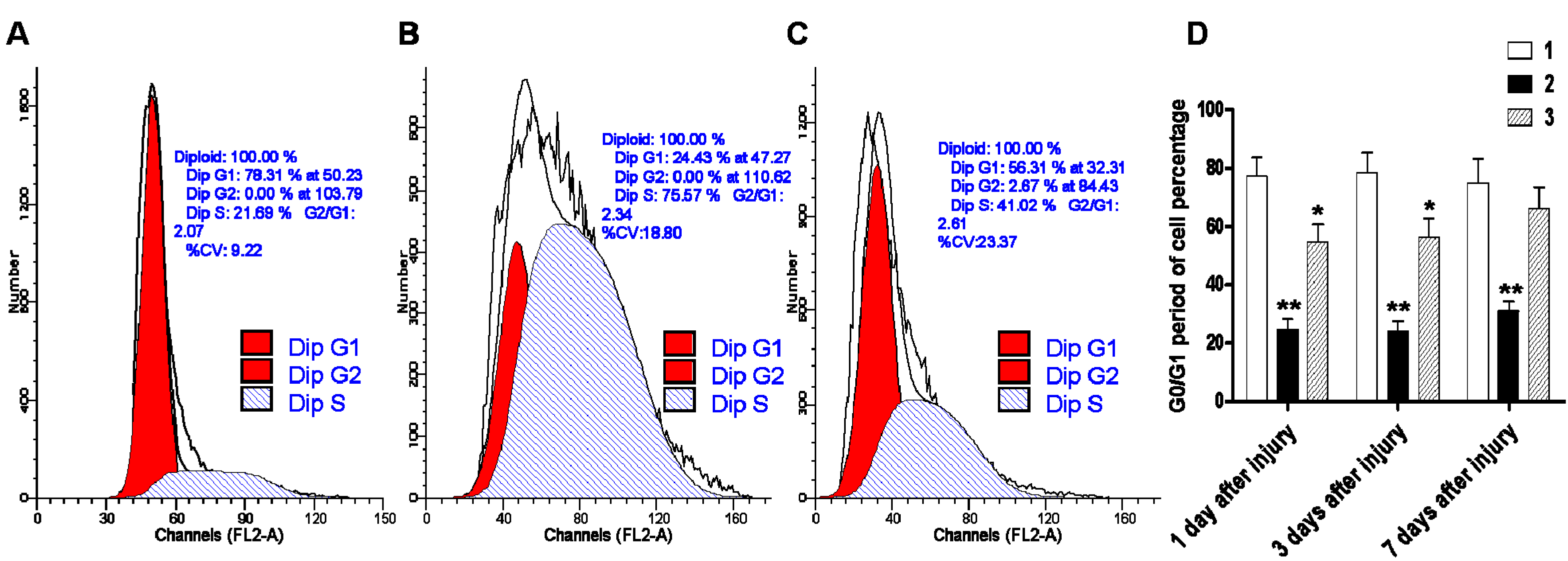
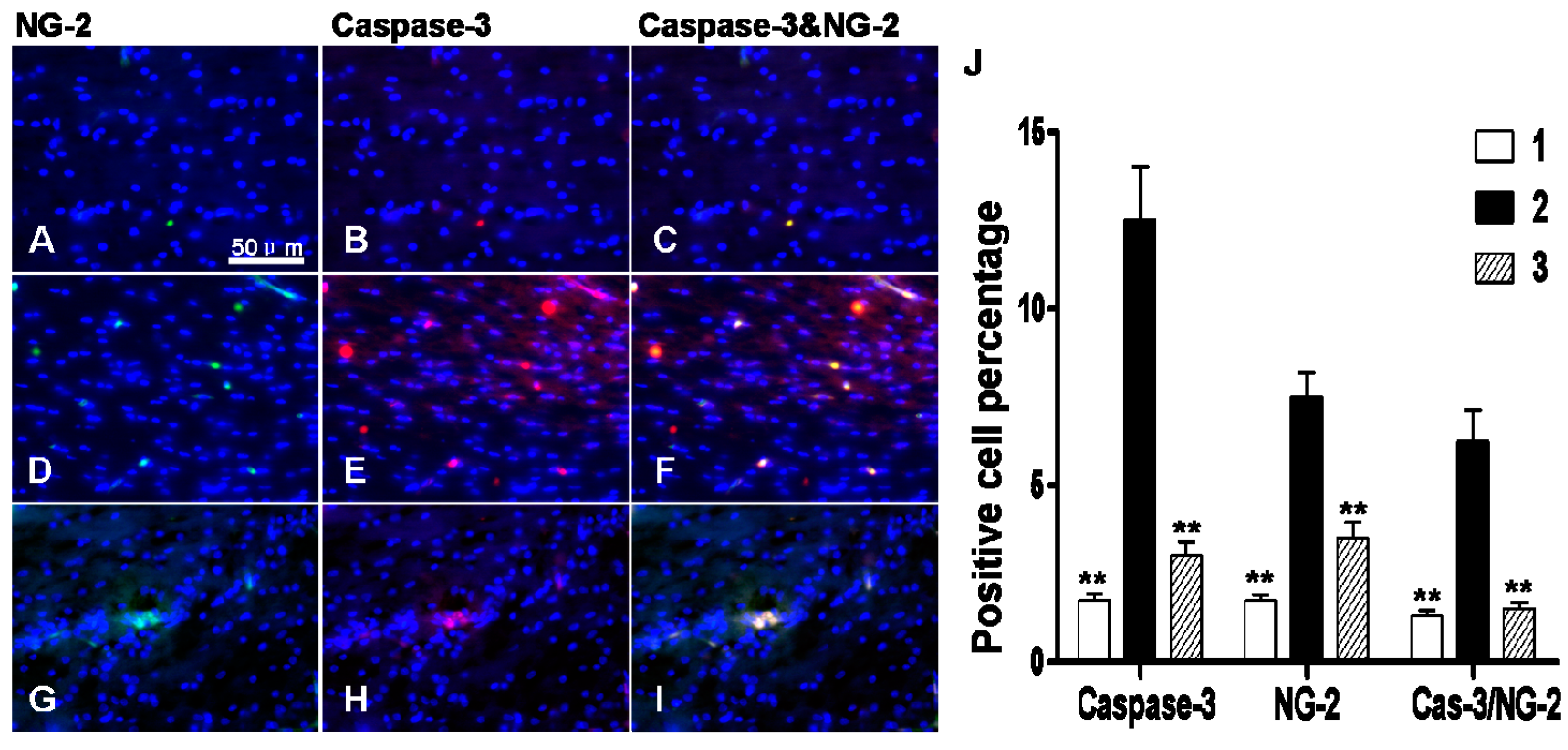
2.4. Early Administration of DIDS Minimized the Apoptotic Ratio after Hypoxic-Ischemic Injury


3. Discussion
4. Experimental Section
4.1. Animal Model
4.2. Experimental Animal Group
4.3. Tissue Harvesting
4.4. RT-PCR
4.5. Western Blot Analysis
4.6. Immunohistochemistry Staining
4.7. Active Oxygen Concentration Detection
4.8. Flow Cytometry
4.9. Luxol Fast Blue (LFB) Staining
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Back, S.A.; Luo, N.L.; Borenstein, N.S.; Levine, J.M.; Volpe, J.J.; Kinney, H.C. Late oligodendrocyte progenitors coincide with the developmental window of vulnerability for human perinatal white matter injury. J. Neurosci. 2001, 21, 1302–1312. [Google Scholar]
- Boylan, G.B.; Young, K.; Panerai, R.B.; Rennie, J.M.; Evans, D.H. Dynamic cerebral autoregulation in sick newborn infants. Pediatr. Res. 2000, 48, 12–17. [Google Scholar] [CrossRef]
- Folkerth, R.D.; Keefe, R.J.; Haynes, R.L.; Trachtenberg, F.L.; Volpe, J.J.; Kinney, H.C. Interferon-γ expression in periventricular leukomalacia in the human brain. Brain Pathol. 2004, 14, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Zierler, S.; Frei, E.; Grissmer, S.; Kerschbaum, H.H. Chloride influx provokes lamellipodium formation in microglial cells. Cell. Physiol. Biochem. 2008, 21, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Ducharme, G.; Newell, E.W.; Pinto, C.; Schlichter, L.C. Small-conductance Cl− channels contribute to volume regulation and phagocytosis in microglia. Eur. J. Neurosci. 2007, 26, 2119–2130. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, K.; Maeta, H.; Yamamoto, A.; Oe, M.; Kosaka, H. Amelioration of myocardial global ischemia/reperfusion injury with volume-regulatory chloride channel inhibitors in vivo. Transplantation 2002, 73, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Malekova, L.; Tomaskova, J.; Novakova, M.; Stefanik, P.; Kopacek, J.; Lakatos, B.; Pastorekova, S.; Krizanova, O.; Breier, A.; Ondrias, K. Inhibitory effect of DIDS, NPPB, and phloretin on intracellular chloride channels. Pflug. Arch. 2007, 455, 349–357. [Google Scholar] [CrossRef]
- Okada, Y.; Maeno, E. Apoptosis, cell volume regulation and volume-regulatory chloride channels. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 377–383. [Google Scholar] [CrossRef]
- Feldhaus, B.; Dietzel, I.D.; Heumann, R.; Berger, R. Effects of interferon-gamma and tumor necrosis factor-alpha on survival and differentiation of oligodendrocyte progenitors. J. Soc. Gynecol. Investig. 2004, 11, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Zhou, D.; Yao, H.; Haddad, G.G. Role of transporters and ion channels in neuronal injury under hypoxia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R451–R457. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Celma, J.; Szydelko, A.; Dutzler, R. Functional characterization of a ClC transporter by solid-supported membrane electrophysiology. J. Gen. Physiol. 2013, 141, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Fern, R.; Moller, T. Rapid ischemic cell death in immature oligodendrocytes: A fatal glutamate release feedback loop. J. Neurosci. 2000, 20, 34–42. [Google Scholar] [PubMed]
- Pamenter, M.E.; Ali, S.S.; Tang, Q.; Finley, J.C.; Gu, X.Q.; Dugan, L.L.; Haddad, G.G. An in vitro ischemic penumbral mimic perfusate increases NADPH oxidase-mediated superoxide production in cultured hippocampal neurons. Brain Res. 2012, 1452, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Milton, R.H.; Abeti, R.; Averaimo, S.; DeBiasi, S.; Vitellaro, L.; Jiang, L.; Curmi, P.M.; Breit, S.N.; Duchen, M.R.; Mazzanti, M. CLIC1 function is required for β-amyloid-induced generation of reactive oxygen species by microglia. J. Neurosci. 2008, 28, 11488–11499. [Google Scholar] [CrossRef] [PubMed]
- Druzhyna, N.M.; Musiyenko, S.I.; Wilson, G.L.; LeDoux, S.P. Cytokines induce nitric oxide-mediated mtDNA damage and apoptosis in oligodendrocytes. Protective role of targeting 8-oxoguanine glycosylase to mitochondria. J. Biol. Chem. 2005, 280, 21673–21679. [Google Scholar]
- Zhao, W.; Zhu, F.; Shen, W.; Fu, A.; Zheng, L.; Yan, Z.; Zhao, L.; Fu, G. Protective effects of DIDS against ethanol-induced gastric mucosal injury in rats. Acta Biochim. Biophys. Sin. 2009, 41, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Hong, J.S. Microglia and inflammation-mediated neurodegeneration: multiple triggers with a common mechanism. Prog. Neurobiol. 2005, 76, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, K.; Strobaek, D.; Christophersen, P.; Ronn, L.C. Chloride channel blockers inhibit iNOS expression and NO production in IFNγ-stimulated microglial BV2 cells. Brain Res. 2009, 1281, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.H.; Cao, Y.N.; Liu, H.T.; Zhang, W.W.; Liu, Y.; Shi, T.W.; Jia, G.L.; Wang, X.M. DIDS attenuates staurosporine-induced cardiomyocyte apoptosis by PI3K/Akt signaling pathway: Activation of eNOS/NO and inhibition of Bax translocation. Cell. Physiol. Biochem. 2008, 22, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Shimizu, T.; Maeno, E.; Tanabe, S.; Wang, X.; Takahashi, N. Volume-sensitive chloride channels involved in apoptotic volume decrease and cell death. J. Membr. Biol. 2006, 209, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Yu, S.; Binek, J.; Chalimoniuk, M.; Zhang, X.; Lo, S.C.; Hannink, M.; Wu, J.; Fritsche, K.; Donato, R.; et al. Distinct signaling pathways for induction of type II NOS by IFNγ and LPS in BV-2 microglial cells. Neurochem. Int. 2005, 47, 298–307. [Google Scholar]
- Pamenter, M.E.; Ryu, J.; Hua, S.T.; Perkins, G.A.; Mendiola, V.L.; Gu, X.Q.; Ellisman, M.H.; Haddad, G.G. DIDS prevents ischemic membrane degradation in cultured hippocampal neurons by inhibiting matrix metalloproteinase release. PLoS ONE 2012, 7, e43995. [Google Scholar] [CrossRef] [PubMed]
- Levine, S. Anoxic-ischemic encephalopathy in rats. Am. J. Pathol. 1960, 36, 1–17. [Google Scholar] [PubMed]
- Cai, Z.; Pang, Y.; Xiao, F.; Rhodes, P.G. Chronic ischemia preferentially causes white matter injury in the neonatal rat brain. Brain Res. 2001, 898, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Uehara, H.; Yoshioka, H.; Kawase, S.; Nagai, H.; Ohmae, T.; Hasegawa, K.; Sawada, T. A new model of white matter injury in neonatal rats with bilateral carotid artery occlusion. Brain Res. 1999, 837, 213–220. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, B.; Quan, H.; Ma, T.; Tian, Y.; Cai, Q.; Li, H. 4,4'-Diisothiocyanostilbene-2,2'-disulfonic Acid (DIDS) Ameliorates Ischemia-Hypoxia-Induced White Matter Damage in Neonatal Rats through Inhibition of the Voltage-Gated Chloride Channel ClC-2. Int. J. Mol. Sci. 2015, 16, 10457-10469. https://doi.org/10.3390/ijms160510457
Zhao B, Quan H, Ma T, Tian Y, Cai Q, Li H. 4,4'-Diisothiocyanostilbene-2,2'-disulfonic Acid (DIDS) Ameliorates Ischemia-Hypoxia-Induced White Matter Damage in Neonatal Rats through Inhibition of the Voltage-Gated Chloride Channel ClC-2. International Journal of Molecular Sciences. 2015; 16(5):10457-10469. https://doi.org/10.3390/ijms160510457
Chicago/Turabian StyleZhao, Baixiong, Hongyu Quan, Teng Ma, Yanping Tian, Qiyan Cai, and Hongli Li. 2015. "4,4'-Diisothiocyanostilbene-2,2'-disulfonic Acid (DIDS) Ameliorates Ischemia-Hypoxia-Induced White Matter Damage in Neonatal Rats through Inhibition of the Voltage-Gated Chloride Channel ClC-2" International Journal of Molecular Sciences 16, no. 5: 10457-10469. https://doi.org/10.3390/ijms160510457
APA StyleZhao, B., Quan, H., Ma, T., Tian, Y., Cai, Q., & Li, H. (2015). 4,4'-Diisothiocyanostilbene-2,2'-disulfonic Acid (DIDS) Ameliorates Ischemia-Hypoxia-Induced White Matter Damage in Neonatal Rats through Inhibition of the Voltage-Gated Chloride Channel ClC-2. International Journal of Molecular Sciences, 16(5), 10457-10469. https://doi.org/10.3390/ijms160510457