
Skeletal Muscle Mitochondrial Energetic Efficiency and Aging


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Age-Related Changes in Mitochondria
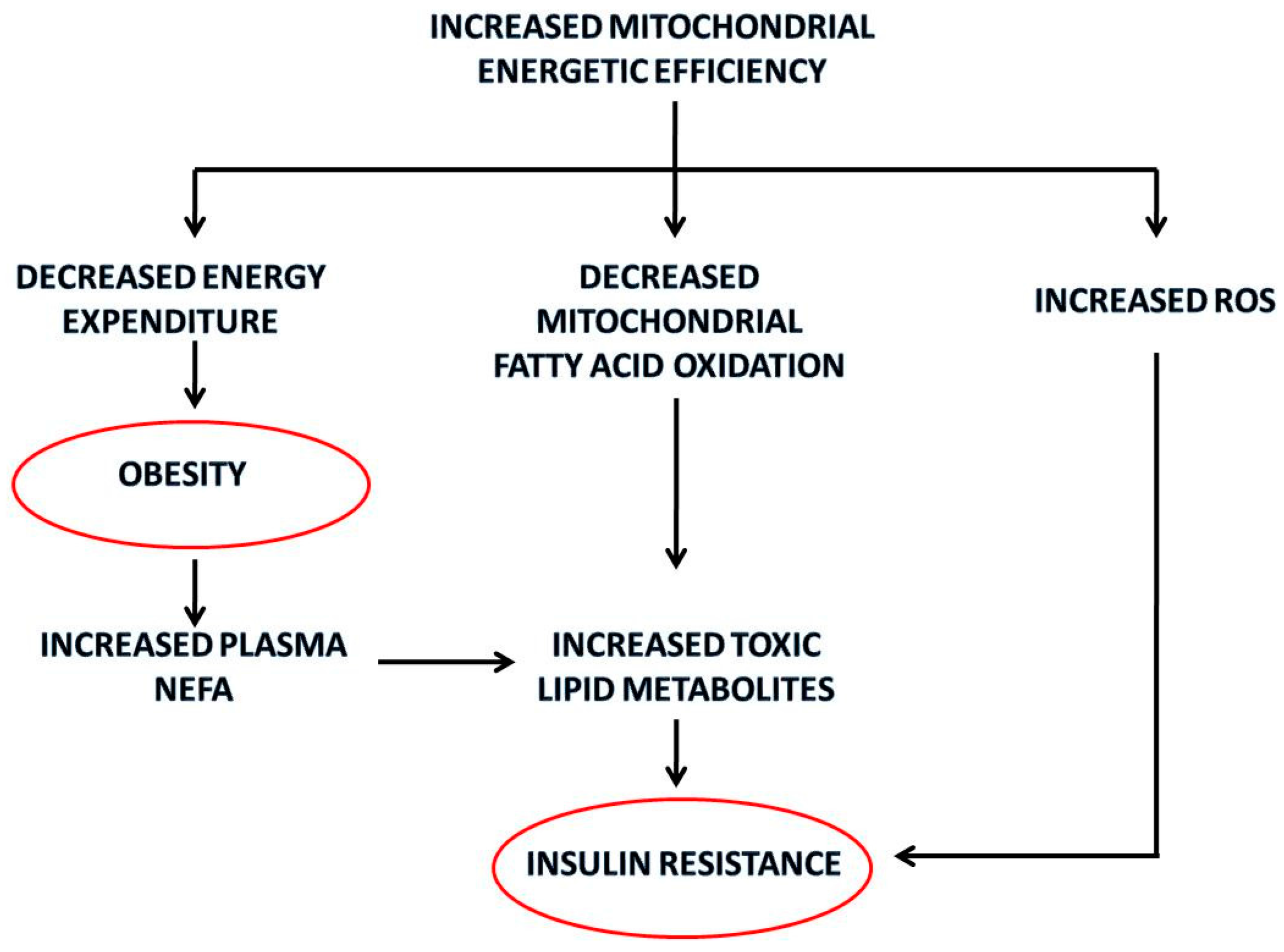
3. Mitochondrial Energetic Efficiency during Aging


4. Oxidative Stress in Aging Mitochondria
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Figueiredo, P.A.; Mota, M.P.; Appell, H.J.; Duarte, J.A. The role of mitochondria in aging of skeletal muscle. Biogerontology 2008, 9, 67–84. [Google Scholar] [CrossRef]
- Kwong, L.K.; Sohal, R.S. Age-related changes in activities of mitochondrial electron transport complexes in various tissues of the mouse. Arch. Biochem. Biophys. 2000, 373, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, J.; Landi, F.; Topinkova, E.; Michel, J.P. Understanding sarcopenia as a geriatric syndrome. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Marzetti, E.; Calvani, R.; Cesari, M.; Buford, T.W.; Lorenzi, M.; Behnke, B.J.; Leeuwenburgh, C. Mitochondrial dysfunction and sarcopenia of aging: From signaling pathways to clinical trials. Int. J. Biochem. Cell Biol. 2013, 45, 2288–2301. [Google Scholar] [CrossRef] [PubMed]
- Shimokata, H.; Tobin, J.; Muller, D.C.; Elahi, D.; Coon, P.J.; Andres, R. Studies in the distribution of body fat: 1. Effects of age, sex and obesity. J. Gerontol. 1989, 44, M66–M73. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G.M.; Reaven, E.P. Age, glucose intolerance, and non-insulin dependent diabetes mellitus. J. Am. Geriatr. Soc. 1985, 286–290. [Google Scholar] [CrossRef]
- Barzilai, N.; Rossetti, L. Relationship between changes in body composition and insulin responsiveness in models of the aging rat. Am. J. Physiol. 1995, 269, E591–E597. [Google Scholar] [PubMed]
- Larkin, L.M.; Reynolds, T.H.; Supiano, M.A.; Kahn, B.B.; Halter, J.B. Effect of aging and obesity on insulin responsiveness and glut-4 glucose transporter content in skeletal muscle of Fisher 344 × Brown Norway rats. J. Gerontol. 2001, 56, B486–B492. [Google Scholar] [CrossRef]
- Iossa, S.; Mollica, M.P.; Lionetti, L.; Crescenzo, R.; Botta, M.; Barletta, A.; Liverini, G. Acetyl-l-carnitine supplementation differently influences nutrient partitioning, serum leptin concentration and skeletal muscle mitochondrial respiration in young and old rats. J. Nutr. 2002, 132, 636–642. [Google Scholar]
- Lenaz, G. Role of mitochondria in oxidative stress and ageing. Biochim. Biophyis. Acta 1998, 1366, 53–67. [Google Scholar] [CrossRef]
- Lenaz, G.; D’Aurelio, M.; Merlo Pich, M.; Genova, M.L.; Ventura, B.; Bovina, C.; Formiggini, G.; Parenti Castelli, G. Mitochondrial bioenergetics in aging. Biochim. Biophys. Acta 2000, 1459, 397–404. [Google Scholar]
- Shigenaga, M.K.; Hagen, T.M.; Ames, B.N. Oxidative damage and mitochondrial decay in aging. Proc. Natl. Acad. Sci. USA 1994, 91, 10771–10778. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Befroy, D.; Dufour, S.; Dziura, J.; Arijan, C.; Rothman, D.L.; DiPietro, L.; Cline, G.W.; Shulman, G.I. Mitochondrial disfunction in the elderly: Possible role in insulin resistance. Science 2003, 300, 1140–1142. [Google Scholar] [CrossRef]
- Bruce, C.R.; Anderson, M.J.; Carey, A.L.; Newman, D.G.; Bonen, A.; Kriketos, A.D.; Cooney, G.J.; Hawley, J.A. Muscle oxidative capacity is a better predictor of insulin sensitivity than lipid status. J. Clin. Endocrinol. Metab. 2003, 88, 5444–5451. [Google Scholar] [CrossRef]
- Drew, B.; Leeuwenburgh, C. Ageing and subcellular distribution of mitochondria: Role of mitochondrial DNA deletions and energy production. Acta Physiol. Scand. 2004, 182, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Mollica, M.P.; Lionetti, L.; Crescenzo, R.; D’Andrea, E.; Ferraro, M.; Liverini, G.; Iossa, S. Heterogeneous bioenergetic behaviour of subsarcolemmal and intermyofibrillar mitochondria in fed and fasted rats. Cell. Mol. Life Sci. 2006, 63, 358–366. [Google Scholar] [CrossRef]
- Figueiredo, P.A.; Powers, S.K.; Ferreira, R.M.; Appell, H.J.; Duarte, J.A. Aging impairs skeletal muscle mitochondrial bioenergetic function. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 21–33. [Google Scholar] [CrossRef]
- Trounce, I.; Byrne, E.; Marzuki, S. Decline in skeletal muscle mitochondrial respiratory chain function: Possible factor in ageing. Lancet 1989, 1, 637–639. [Google Scholar] [CrossRef] [PubMed]
- Conley, K.E.; Jubrias, S.A.; Esselman, P.C. Oxidative capacity and aging in human muscle. J. Physiol. 2000, 526, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Gouspillou, G.; Bourdel-Marchasson, I.; Rouland, R.; Calmettes, G.; Biran, M.; Deschodt-Arsac, V.; Miraux, S.; Thiaudiere, E.; Pasdois, P.; Detaille, D.; et al. Mitochondrial energetics is impaired in vivo in aged skeletal muscle. Aging Cell 2014, 39–48. [Google Scholar] [CrossRef]
- Gouspillou, G.; Sgarioto, N.; Kapchinsky, S.; Purves-Smith, F.; Norris, B.; Pion, C.H.; Barbat-Artigas, S.; Lemieux, F.; Taivassalo, T.; Morais, J.A.; et al. Increased sensitivity to mitochondrial permeability transition and myonuclear translocation of endonuclease G in atrophied muscle of physically active older humans. FASEB J. 2014, 28, 1621–1633. [Google Scholar] [CrossRef]
- Gouspillou, G.; Bourdel-Marchasson, I.; Rouland, R.; Calmettes, G.; Franconi, J.M.; Deschodt-Arsac, V.; Diolez, P. Alteration of mitochondrial oxidative phosphorylation in aged skeletal muscle involves modification of adenine nucleotide translocator. Biochim. Biophys. Acta 2010, 1797, 143–151. [Google Scholar]
- Picard, M.; Ritchie, D.; Thomas, M.M.; Wright, K.J.; Hepple, R.T. Alterations in intrinsic mitochondrial function with aging are fiber type-specific and do not explain differential atrophy between muscles. Aging Cell 2011, 10, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, U.F.; Krustrup, P.; Kjaer, M.; Rasmussen, H.N. Experimental evidence against the mitochondrial theory of aging. A study on isolated human skeletal muscle mitochondria. Exp. Gerontol. 2003, 38, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, U.F.; Krustrup, P.; Kjaer, M.; Rasmussen, H.N. Human skeletal muscle mitochondrial metabolism in youth and senescence: No signs of functional changes of ATP formation and mitochondrial capacity. Pflug. Arch. 2003, 446, 270–278. [Google Scholar]
- Desai, V.G.; Weindruch, R.; Hart, R.W.; Feuers, R.J. Influences of age and dietary restriction on gastrocnemius electron transport system activities in mice. Arch. Biochem. Biophys. 1996, 333, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hagen, J.L.; Krause, D.J.; Baker, D.J.; Fu, M.; Tarnopolsky, M.A.; Hepple, R.T. Skeletal muscle aging in F344BN F1-hybrid rats: I. Mitochondrial dysfunction contributes to the age-associated reduction in VO2max. J. Gerontol. 2004, 59, 1099–1110. [Google Scholar] [CrossRef]
- Lombardi, A.; Silvestri, E.; Cioffi, F.; Senese, R.; Lanni, A.; Goglia, F.; de Lange, P.; Moreno, M. Defining the trascriptomic and proteomic profiles of rat ageing skeletal muscle by the use of a cDNA array, 2D- and Blue native-PAGE approach. J. Proteomics 2009, 72, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Barrientos, A.; Casademont, J.; Rotig, A.; Miro, O.; Urbano-Marquez, A.; Rustin, P.; Cardellach, F. Absence of relationship between the level of electron transport chain activities and aging in human skeletal muscle. Biochem. Biophys. Res. Commun. 1996, 229, 536–539. [Google Scholar] [CrossRef]
- Lyons, C.N.; Mathieu-Costello, O.; Moyes, C.D. Regulation of skeletal muscle mitochondrial content during aging. J. Gerontol. 2006, 61, 3–13. [Google Scholar] [CrossRef]
- Kerner, J.; Turkaly, P.J.; Minkler, P.E.; Hoppel, C.L. Aging skeletal muscle mitochondria in the rat: Decreased uncoupling protein-3 content. Am. J. Physiol. 2001, 281, E1054–E1062. [Google Scholar]
- Chabi, B.; Ljubicic, V.; Menzies, K.J.; Huang, J.H.; Saleem, A.; Hood, D.A. Mitochondrial function and apoptotic susceptibility in aging skeletal muscle. Aging Cell 2008, 7, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Costello, O.; Ju, Y.; Trejo-Morales, M.; Cui, L. Greater capillary-fiber interface per fiber mitochondrial volume in skeletal muscles of old rats. J. Appl. Physiol. 2005, 99, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Callahan, D.M.; Bedrin, N.G.; Subramanian, M.; Berking, J.; Ades, P.A.; Toth, M.J.; Miller, M.S. Age-related structural alterations in human skeletal muscle fibers and mitochondria are sex specific: Relationship to single-fiber function. J. Appl. Physiol. 2014, 116, 1582–1592. [Google Scholar] [CrossRef] [PubMed]
- Konopka, R.; Suer, M.K.; Wolff, C.A.; Harber, M.P. Markers of human skeletal muscle mitochondrial biogenesis and quality control: Effects of age and aerobic exercise training. J. Gerontol. 2014, 69, 371–378. [Google Scholar] [CrossRef]
- Betik, C.; Thomas, M.M.; Wright, K.J.; Riel, C.D.; Hepple, R.T. Exercise training from late middle age until senescence does not attenuate the declines in skeletal muscle aerobic function. Am. J. Physiol. 2009, 297, R744–R755. [Google Scholar]
- Ljubicic, V.; Hood, D.A. Diminished contraction-induced intracellular signaling towards mitochondrial biogenesis in aged skeletal muscle. Aging Cell 2009, 8, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Ljubicic, V.; Joseph, A.M.; Adhihetty, P.J.; Huang, J.H.; Saleem, A.; Uguccioni, G.; Hood, D.A. Molecular basis for an attenuated mitochondrial adaptive plasticity in aged skeletal muscle. Aging 2009, 1, 818–830. [Google Scholar] [PubMed]
- Farrar, R.P.; Martin, T.P.; Ardies, C.M. The interaction of aging and endurance exercise upon the mitochondrial function of skeletal muscle. J. Gerontol. 1981, 36, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Drew, B.; Phaneuf, S.; Dirks, A.; Selman, C.; Gredilla, R.; Lezza, A.; Barja, G.; Leeuwenburgh, C. Effects of aging and caloric restriction on mitochondrial energy production in gastrocnemius muscle and heart. Am. J. Physiol. 2003, 284, R474–R480. [Google Scholar]
- Nielsen, J.; Suetaa, C.; Hvid, L.G.; Schroder, H.D.; Aagaard, P.; Orteblad, N. Subcellular localization-dependent decrements in skeletal muscle glycogen and mitochondria content following short-term disuse in young and old men. Am. J. Physiol. 2010, 299, E1053–E1060. [Google Scholar]
- Holloszy, J.O. Skeletal muscle “mitochondrial deficiency” does not mediate insulin resistance. Am. J. Clin. Nutr. 2009, 89, 463S–466S. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B. Intrinsic and extrinsic uncoupling of oxidative phosphorylation. Biochim. Biophys. Acta 2003, 1604, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Stucki, J.W. The optimal efficiency and the economic degrees of coupling of oxidative phosphorylation. Eur. J. Biochem. 1980, 109, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, D.F.S.; Brown, G.C. Cellular energy utilisation and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997, 77, 731–758. [Google Scholar] [PubMed]
- Skulachev, V.P. Fatty acid circuit as a physiological mechanism of uncoupling of oxidative phosphorylation. FEBS Lett. 1991, 294, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Jezek, P.; Engstova, H.; Zackova, M.; Vercesi, A.E.; Costa, A.D.T.; Arruda, P.; Garlid, K.D. Fatty acid cycling mechanism and mitochondrial uncoupling proteins. Biochim. Biophys. Acta 1998, 1365, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Soboll, S.; Grundel, S.; Schwabe, U.; Scholtz, R. Influence of fatty acids on energy metabolism. 2. Kinetics of changes in metabolic rates and changes in subcellular adenine nucleotide contents and pH gradients following addition of octanoate and oleate in perfused rat liver. Eur. J. Biochem. 1984, 141, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Amara, C.E.; Shankland, E.G.; Jubrias, S.A.; Marcinek, D.J.; Kushmerick, M.J.; Conley, K.E. Mild mitochondrial uncoupling impacts cellular aging in human muscles in vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Fannin, S.W.; Lesnefsky, E.J.; Slabe, T.J.; Hassan, M.O.; Hoppel, C.L. Aging selectively decreases oxidative capacity in rat heart interfibrillar mitochondria. Arch. Biochem. Biophys. 1999, 372, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Iossa, S.; Mollica, M.P.; Lionetti, L.; Crescenzo, R.; Tasso, R.; Liverini, G. A possible link between skeletal muscle mitochondrial efficiency and age-induced insulin resistance. Diabetes 2004, 53, 2861–2866. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Bianco, F.; Mazzoli, A.; Giacco, A.; Liverini, G.; Iossa, S. Alterations in proton leak, oxidative status and uncoupling protein 3 content in skeletal muscle subsarcolemmal and intermyofibrillar mitochondria in old rats. BMC Geriatr. 2014, 14, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koltai, E.; Hart, N.; Taylor, A.W.; Goto, S.; Ngo, J.K.; Davies, K.J.A.; Radak, Z. Age-associated declines in mitochondrial biogenesis and protein quality control factors are minimized by exercise training. Am. J. Physiol. 2012, 303, R127–R134. [Google Scholar]
- Hafner, R.P.; Brown, G.C.; Brand, M.D. Analysis of the control of respiration rate, phosphorylation rate, proton leak rate and protonmotive force in isolated mitochondria using the “top-down” approach of metabolic control theory. Eur. J. Biochem. 1990, 188, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Chien, L.F.; Ainscow, E.K.; Rolfe, D.F.S.; Porter, R.K. The causes and functions of mitochondrial proton leak. Biochim. Biophys. Acta 1994, 1187, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Cairns, C.B.; Walther, J.; Harken, A.H.; Banerjee, A. Mitochondrial oxidative phosphorylation thermodynamic efficiencies reflect physiological organ roles. Am. J. Physiol. 1998, 274, R1376–R1383. [Google Scholar] [PubMed]
- Santanasto, A.J.; Glynn, N.W.; Jubrias, S.A.; Conley, K.E.; Boudreau, R.M.; Amati, F.; Mackey, D.C.; Simonsick, E.M.; Strotmeyer, E.S.; Coen, P.M.; et al. Skeletal muscle mitochondrial function and fatigability in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2015. [Google Scholar] [CrossRef]
- Stannard, S.R.; Johnson, N.A. Insulin resistance and elevated triglyceride in muscle: More important for survival than thrifty genes? J. Physiol. 2003, 554, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, S.S.; Skulachev, V.P.; Starkov, A.A. High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. FEBS Lett. 1997, 416, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Mailloux, R.J.; Harper, M.E. Uncoupling proteins and the control of mitochondrial reactive oxygen species production. Free Radic. Biol. Med. 2011, 51, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D. Uncoupling to survive? The role of mitochondrial inefficiency in ageing. Exp. Gerontol. 2000, 35, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, A.; Busiello, R.A.; Napolitano, L.; Cioffi, F.; Moreno, M.; de Lange, P.; Silvestri, E.; Lanni, A.; Goglia, F. UCP3 translocates lipid hydroperoxide and mediates lipid hydroperoxide-dependent mitochondrial uncoupling. J. Biol. Chem. 2010, 285, 16599–16605. [Google Scholar] [CrossRef] [PubMed]
- Goglia, F.; Skulachev, V.P. A function for novel uncoupling proteins: Antioxidant defence of mitochondrial matrix by translocating fatty acid peroxides from the inner to the outer membrane leaflet. FASEB J. 2003, 17, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Divakaruni, A.S.; Brand, M.D. The regulation and physiology of mitochondrial proton leak. Physiology 2011, 26, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Lal, S.B.; Ramsey, J.J.; Monemdjou, S.; Weindruch, R.; Harper, M.E. Effects of caloric restriction on skeletal muscle mitochondrial proton leak in aging rats. J. Gerontol. 2001, 56, B116–B122. [Google Scholar] [CrossRef]
- Marcinek, D.J.; Schenkman, K.A.; Ciesielski, W.A.; Lee, D.; Conley, K.E. Reduced mitochondrial coupling in vivo alters cellular energetic in aged mouse skeletal muscle. J. Physiol. 2005, 569, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Hood, D. Plasticity in skeletal, cardiac, and smooth muscle: Contractile activity-induced mitochondrial biogenesis in skeletal muscle. J. Appl. Physiol. 2001, 90, 1137–1157. [Google Scholar] [PubMed]
- Beyer, R.E.; Starnes, J.W.; Edington, D.W.; Lipton, R.J.; Compton, R.T.; Kwasman, M.A. Exercise-induced reversal of age-related declines of oxidative reactions, mitochondrial yield, and flavins in skeletal muscle of the rat. Mech. Ageing Dev. 1984, 24, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, L.; Ramsey, J.J.; Hagopian, K.; Weindruch, R.; Harper, M.E. Long-term caloric restriction increases UCP3 content but decreases proton leak and reactive oxygen species production in rat skeletal muscle mitochondria. Am. J. Physiol. 2005, 289, E429–E438. [Google Scholar]
- Barazzoni, R.; Nair, K.S. Changes in uncoupling protein-2 and -3 expression in aging rat skeletal muscle, liver, and heart. Am. J. Physiol. 2001, 280, E413–E419. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crescenzo, R.; Bianco, F.; Mazzoli, A.; Giacco, A.; Liverini, G.; Iossa, S. Skeletal Muscle Mitochondrial Energetic Efficiency and Aging. Int. J. Mol. Sci. 2015, 16, 10674-10685. https://doi.org/10.3390/ijms160510674
Crescenzo R, Bianco F, Mazzoli A, Giacco A, Liverini G, Iossa S. Skeletal Muscle Mitochondrial Energetic Efficiency and Aging. International Journal of Molecular Sciences. 2015; 16(5):10674-10685. https://doi.org/10.3390/ijms160510674
Chicago/Turabian StyleCrescenzo, Raffaella, Francesca Bianco, Arianna Mazzoli, Antonia Giacco, Giovanna Liverini, and Susanna Iossa. 2015. "Skeletal Muscle Mitochondrial Energetic Efficiency and Aging" International Journal of Molecular Sciences 16, no. 5: 10674-10685. https://doi.org/10.3390/ijms160510674
APA StyleCrescenzo, R., Bianco, F., Mazzoli, A., Giacco, A., Liverini, G., & Iossa, S. (2015). Skeletal Muscle Mitochondrial Energetic Efficiency and Aging. International Journal of Molecular Sciences, 16(5), 10674-10685. https://doi.org/10.3390/ijms160510674