
Chromate Reductase YieF from Escherichia coli Enhances Hexavalent Chromium Resistance of Human HepG2 Cells


Abstract
:1. Introduction
2. Results and Discussions
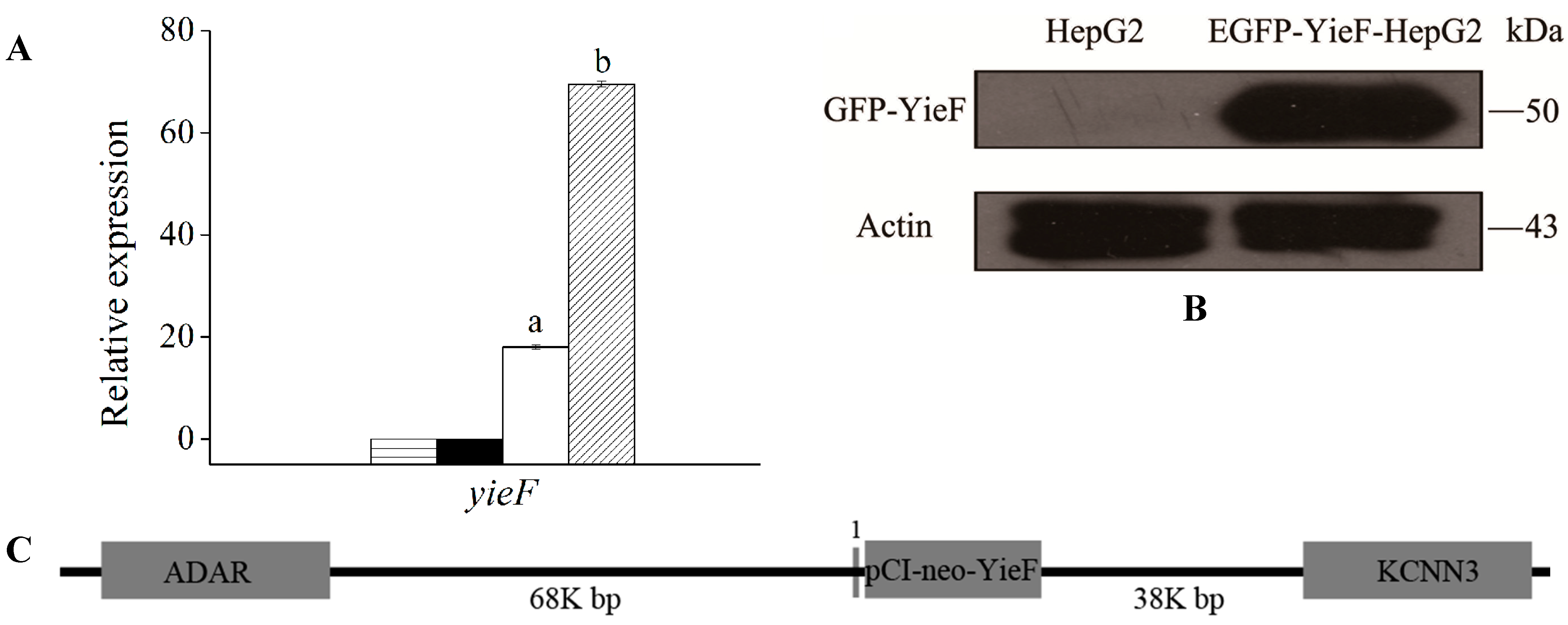
2.1. Expression of YieF under Cr(VI) Stress

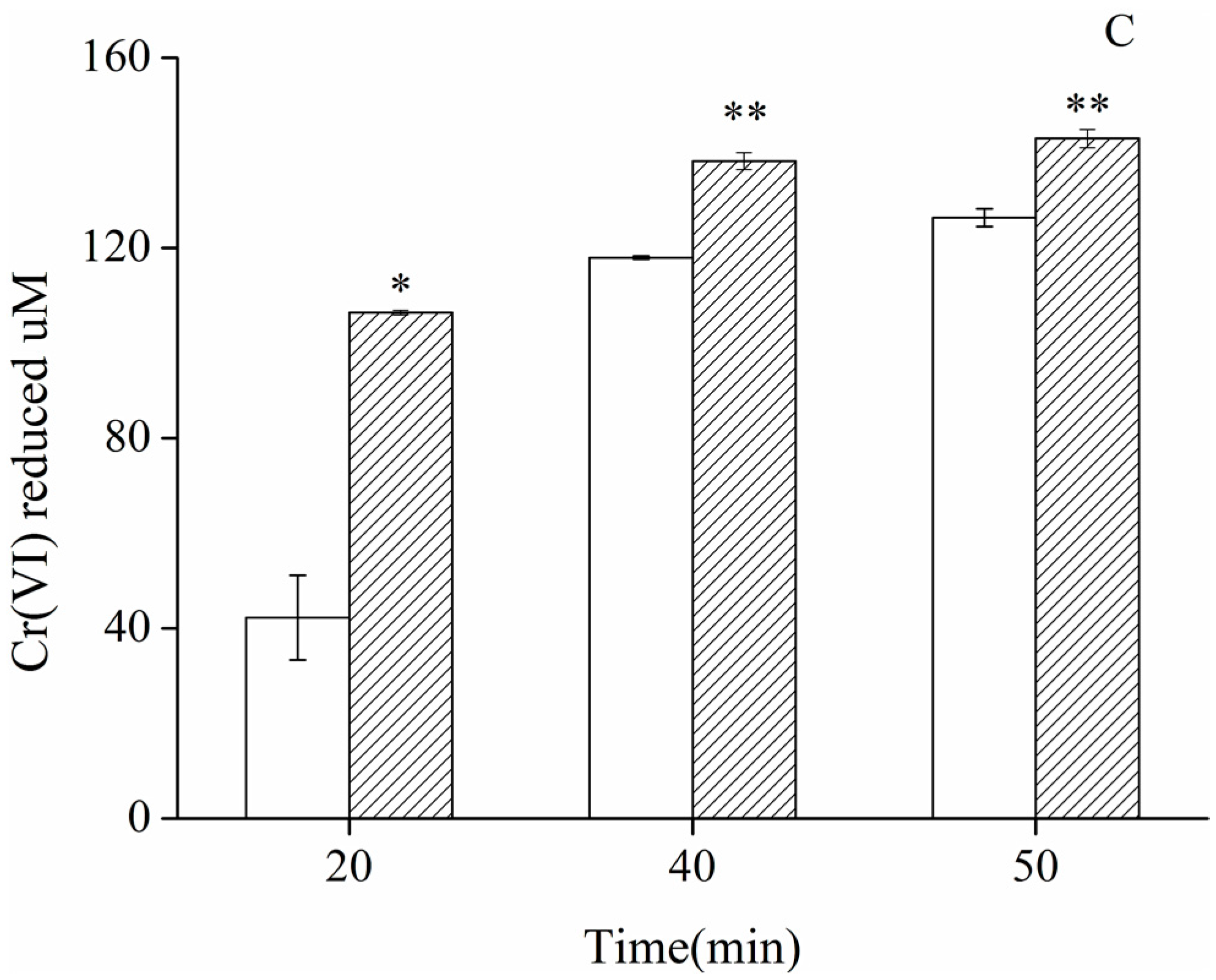
2.2. Enhanced Cr(VI)-Reducing Ability of Cultured HepG2-YieF Cells and Crude Cell Extracts


2.3. Expression Profiles of Glutathione Synthetase and Glutathione Reductase under Cr(VI) Treatment
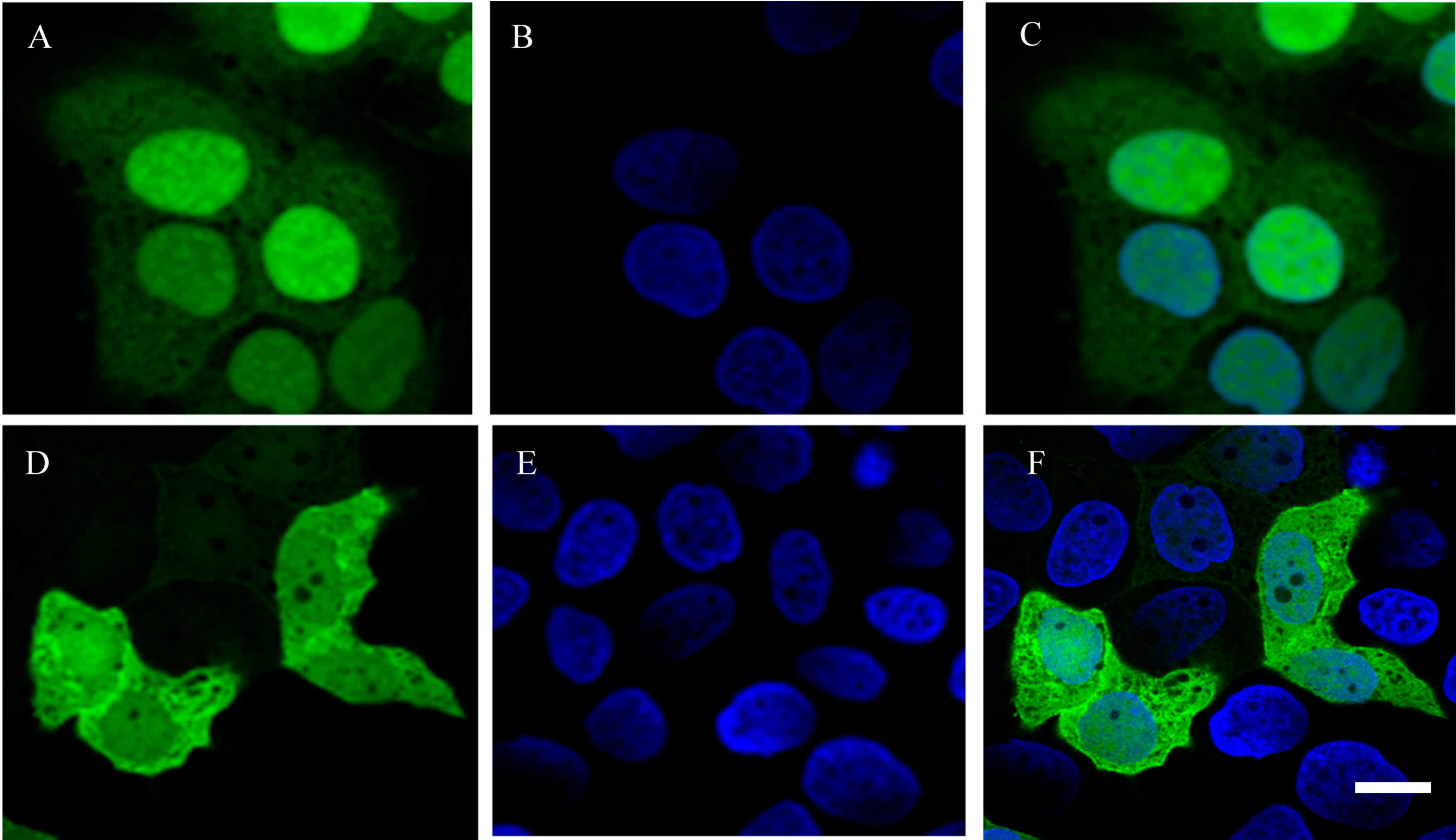
2.4. Localization of Recombinant EGFP-YieF Fusion Protein


3. Experimental Section
3.1. Plasmid Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences |
|---|---|
| pCI-neo-YieF | 5ʹ-GAATTCTCTAGAATGTCTGAAAAATTGCAGGT-3ʹ/ 5ʹ-GAATTCCCCGGGTCAGATCTTAACTCGCTGAATAA-3ʹ |
| pEGFP-N1-YieF | 5ʹ-GAATTCCTCGAGATGTCTGAAAAATTGCAGGT-3ʹ/ 5ʹ-GAATTCCTGCAGGATCTTAACTCGCTGAATAAACT-3ʹ |
| yieF | 5ʹ-AGCTCATTTAATGGCATGG-3ʹ/ 5ʹ-ATCAAGGGAATGTCGGCAA-3ʹ |
| glutathione synthetase | 5ʹ-TGGTCCAGTGCATTTCAGAG-3ʹ/ 5ʹ-TTGGTTCGAAGTAGATGCCC-3ʹ |
| glutathione reductase | 5ʹ-TTACTGCAGTTCCCGGTAGG-3ʹ/ 5ʹ-CTCAGGTCCTTGGTATTCGG-3ʹ |
| β-actin | 5ʹ-ACATCCGCAAAGACCTGTATG-3ʹ/ 5ʹ-GCCAG AG-CAGTGATCTCCTT-3ʹ |
| SP * | 5ʹ-TGACGGTTCACTAAACGAGCTCTGC-3ʹ/ 5ʹ-ATCGCAGTTGTTACGACATTTTGGA-3ʹ/ 5ʹ-ATCCCCGTGAGTCAAACCGCTATC-3ʹ |
3.2. Cell Culture, DNA Transfection and Cell Selection
3.3. Western Blotting
3.4. Genome Walking
3.5. Chromate Reduction of Cell Culture and Crude Cell Extracts
3.6. Expression Profiles of YieF, Glutathione Synthetase and Glutathione Reductase under Cr(VI) Treatment
3.7. Laser Scanning Confocal Microscopy
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bryant, H.E.; Ying, S.; Helleday, T. Homologous recombination is involved in repair of chromium-induced DNA damage in mammalian cells. Mutat. Res. 2006, 599, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wuana, R.A.; Okieimen, F.E. Heavy metals in contaminated soils: A review of sources, chemistry, risks and best available strategies for remediation. ISRN Ecol. 2011, 2011. [Google Scholar] [CrossRef]
- Di Bona, K.R.; Love, S.; Rhodes, N.R.; McAdory, D.; Sinha, S.H.; Kern, N.; Kent, J.; Strickland, J.; Wilson, A.; Beaird, J.; et al. Chromium is not an essential trace element for mammals: Effects of a “low-chromium” diet. J. Biol. Inorg. Chem. 2011, 16, 381–390. [Google Scholar]
- Kimbrough, D.E.; Cohen, Y.; Winer, A.M.; Creelman, L.; Mabuni, C. A critical assessment of chromium in the environment. Crit. Rev. Environ. Sci. Technol. 1999, 29, 1–46. [Google Scholar] [CrossRef]
- Costa, M.; Klein, C.B. Toxicity and carcinogenicity of chromium compounds in humans. CRC Crit. Rev. Toxicol. 2006, 6, 155–163. [Google Scholar] [CrossRef]
- Leonard, A.; Lauwerys, R. Carcinogenicity and mutagenicity of chromium. Mutat. Res. 1980, 76, 227–239. [Google Scholar] [CrossRef]
- Megharaj, M.; Avudainayagam, S.; Naidu, R. Toxicity of hexavalent chromium and its reduction by bacteria isolated from soil contaminated with tannery waste. Curr. Microbiol. 2003, 47, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Straif, K.; Benbrahim-Tallaa, L.; Baan, R.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Bouvard, V.; Guha, N.; Freeman, C.; Galichet, L. A review of human carcinogens—Part C: Metals, arsenic, dusts, and fibres. Lancet Oncol. 2009, 10, 453–454. [Google Scholar] [CrossRef]
- Huamain, C.; Chunrong, Z.; Cong, T.; Yongguan, Z. Heavy metal pollution in soils in China: Status and countermeasures. Ambio 1999, 28, 130–134. [Google Scholar]
- Nriagu, J.O.; Nieboer, E. Chromium in the Natural and Human Environments; John Wiley & Sons: Hoboken, NJ, USA, 1988. [Google Scholar]
- Loza-Tavera, H.; Torres-Guzmän, J.C.; Moreno-Sänchez, R. Interactions of chromium with microorganisms and plants. FEMS Microbiol. Rev. 2001, 25. [Google Scholar] [CrossRef]
- Itoh, M.; Nakamura, M.; Suzuki, T.; Kawai, K.; Horitsu, H.; Takamizawa, K. Mechanism of chromium (VI) toxicity in Escherichia coli: Is hydrogen peroxide essential in Cr(VI) toxicity. J. Biochem. 1995, 117, 780–786. [Google Scholar] [PubMed]
- Bagchi, D.; Bagchi, M.; Stohs, S.J. Chromium(VI)-induced oxidative stress, apoptotic cell death and modulation of p53 tumor suppressor gene. In Molecular Mechanisms of Metal Toxicity and Carcinogenesis; Springer: New York City, NY, USA, 2001; pp. 149–158. [Google Scholar]
- Mikalsen, A.; Alexander, J.; Wallin, H.; Ingelman-Sundsberg, M.; Anderson, R. Reductive metabolism and protein binding of chromium(VI) by P450 protein enzymes. Carcinogenesis 1991, 12, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Levis, A.; Majone, F. Cytotoxic and clastogenic effects of soluble chromium compounds on mammalian cell cultures. Br. J. Cancer 1979, 40, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Tebo, B.M.; Obraztsova, A.Y. Sulfate-reducing bacterium grows with Cr(VI), U(VI), Mn(IV), and Fe(III) as electron acceptors. FEMS Microbiol. Lett. 1998, 162, 193–198. [Google Scholar]
- Shen, H.; Wang, Y. Characterization of enzymatic reduction of hexavalent chromium by Escherichia coli ATCC 33456. Appl. Environ. Microbiol. 1993, 59, 3771–3777. [Google Scholar] [PubMed]
- Zheng, Z.; Li, Y.; Zhang, X.; Liu, P.; Ren, J.; Wu, G.; Zhang, Y.; Chen, Y.; Li, X. A Bacillus subtilis strain can reduce hexavalent chromium to trivalent and an nfrA gene is involved. Int. Biodeterior. Biodegrad. 2015, 97, 90–96. [Google Scholar] [CrossRef]
- Barak, Y.; Thorne, S.H.; Ackerley, D.F.; Lynch, S.V.; Contag, C.H.; Matin, A. New enzyme for reductive cancer chemotherapy, YieF, and its improvement by directed evolution. Mol. Cancer Ther. 2006, 5, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, D.; Gonzalez, C.; Keyhan, M.; Blake, R.; Matin, A. Mechanism of chromate reduction by the Escherichia coli protein, NfsA, and the role of different chromate reductases in minimizing oxidative stress during chromate reduction. Environ. Microbiol. 2004, 6, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.H.; Moreno-Sánchez, R.; Cervantes, C. Chromate efflux by means of the ChrA chromate resistance protein from Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 7398–7400. [Google Scholar] [PubMed]
- Moch, C.; Schrogel, O.; Allmansberger, R. Transcription of the nfrA-ywcH operon from Bacillus subtilis is specifically induced in response to heat. J. Bacteriol. 2000, 182, 4384–4393. [Google Scholar] [CrossRef] [PubMed]
- Zhitkovich, A. Importance of chromium-DNA adducts in mutagenicity and toxicity of chromium(VI). Chem. Res. Toxicol. 2005, 18, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Dayan, A.; Paine, A. Mechanisms of chromium toxicity, carcinogenicity and allergenicity: Review of the literature from 1985 to 2000. Hum. Exp. Toxicol. 2001, 20, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.-E.; Kim, I.-G.; Kim, W.-S.; Suh, S.-C.; Kim, B.-D.; Rhim, S.-L. Expression of chromium(VI) reductase gene of heavy metal reducing bacteria in tobacco plants. J. Plant Biotechnol. 2001, 3, 13–17. [Google Scholar]
- Hawkes, H.J.; Karlenius, T.C.; Tonissen, K.F. Regulation of the human thioredoxin gene promoter and its key substrates: A study of functional and putative regulatory elements. Biochim. Biophys. Acta 2014, 1840, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Zhitkovich, A.; Voitkun, V.; Costa, M. Glutathione and free amino acids form stable complexes with DNA following exposure of intact mammalian cells to chromate. Carcinogenesis 1995, 16, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Xing, M.-L.; Shen, Y.; Zhu, X.; Xu, L.-H. Oral administration of Cr(VI) induced oxidative stress, DNA damage and apoptotic cell death in mice. Toxicology 2006, 228, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Pettine, M.; Capri, S. Digestion treatments and risks of Cr(III)–Cr(VI) interconversions during Cr(VI) determination in soils and sediments—A review. Anal. Chim. Acta 2005, 540, 231–238. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wu, G.; Zhang, Y.; Wu, D.; Li, X.; Liu, P. Chromate Reductase YieF from Escherichia coli Enhances Hexavalent Chromium Resistance of Human HepG2 Cells. Int. J. Mol. Sci. 2015, 16, 11892-11902. https://doi.org/10.3390/ijms160611892
Liu X, Wu G, Zhang Y, Wu D, Li X, Liu P. Chromate Reductase YieF from Escherichia coli Enhances Hexavalent Chromium Resistance of Human HepG2 Cells. International Journal of Molecular Sciences. 2015; 16(6):11892-11902. https://doi.org/10.3390/ijms160611892
Chicago/Turabian StyleLiu, Xuan, Gaofeng Wu, Yanli Zhang, Dan Wu, Xiangkai Li, and Pu Liu. 2015. "Chromate Reductase YieF from Escherichia coli Enhances Hexavalent Chromium Resistance of Human HepG2 Cells" International Journal of Molecular Sciences 16, no. 6: 11892-11902. https://doi.org/10.3390/ijms160611892
APA StyleLiu, X., Wu, G., Zhang, Y., Wu, D., Li, X., & Liu, P. (2015). Chromate Reductase YieF from Escherichia coli Enhances Hexavalent Chromium Resistance of Human HepG2 Cells. International Journal of Molecular Sciences, 16(6), 11892-11902. https://doi.org/10.3390/ijms160611892