
The Role of Alternative Splicing in the Control of Immune Homeostasis and Cellular Differentiation


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Alternative Splicing of Pre-mRNA

3. HnRNP Proteins and Their Role in Pre-mRNA Alternative Splicing
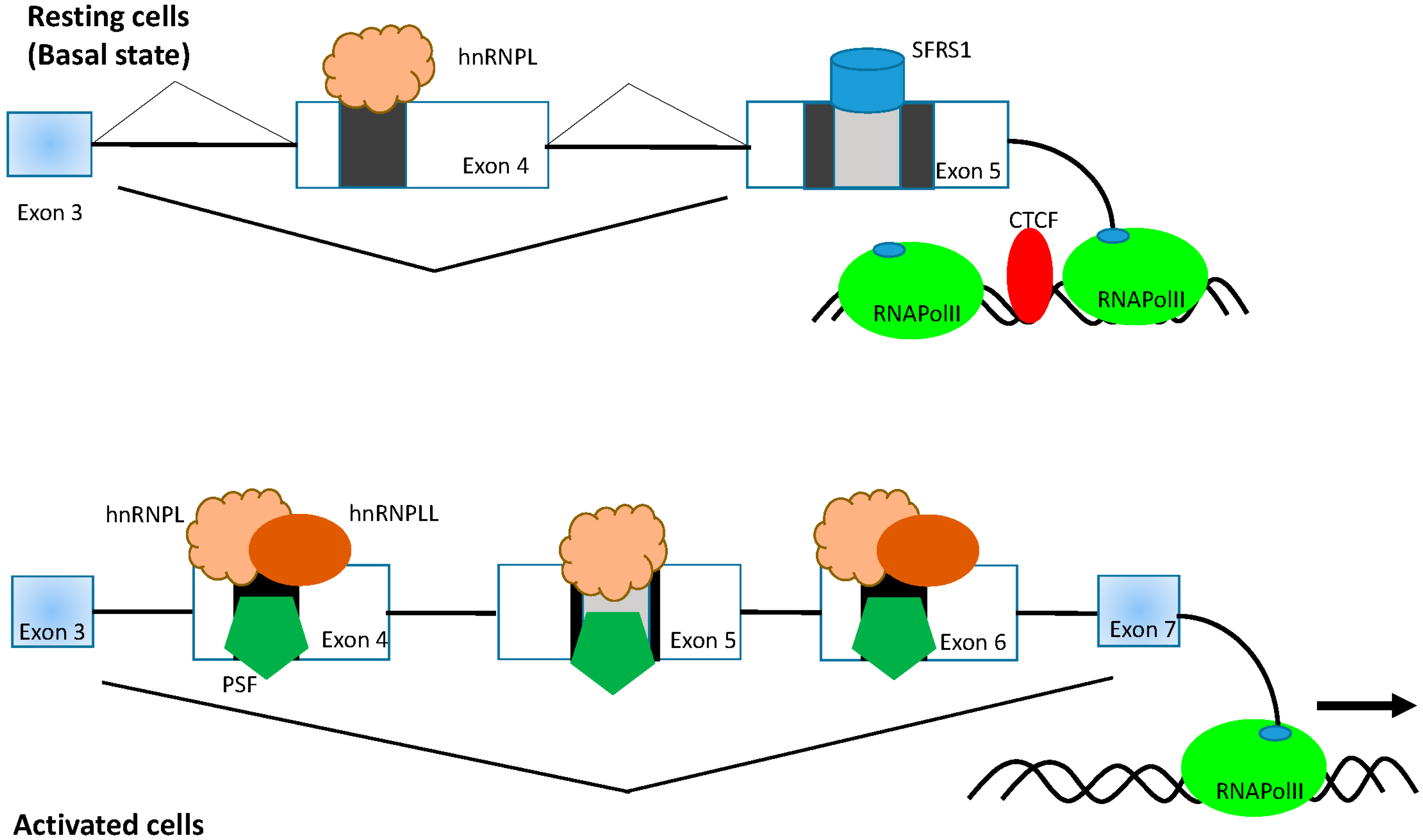
3.1. HnRNPL
3.2. HnRNPLL

3.3. Polypyrimidine-Tract-Binding Protein (PTB)-Associated Splicing Factor (PSF)
3.4. Serine/Arginine-Rich Splicing Factor 1 (SRSF1)
3.5. The Src-Associated Protein in Mitosis of 68 kDa (Sam68)
3.6. CCCTC Binding Factor (CTCF)
3.7. T cell Restricted Intracellular Antigen 1 (TIA-1)
4. Immunity versus Tolerance
5. Control of Apoptotic Effector Proteins by Alternative Splicing
6. Alternative Splicing and Control of T Cell Activation
7. Control of T Cell Homeostasis
8. Alternative Splicing in B Cells and the Role of hnRNPLL in B Cell Development and Function
9. Future Directions
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar]
- Palm, N.W.; Medzhitov, R. Pattern recognition receptors and control of adaptive immunity. Immunol. Rev. 2009, 227, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Akira, S. Innate immunity to pathogens: Diversity in receptors for microbial recognition. Immunol. Rev. 2009, 227, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed]
- Modrek, B.; Resch, A.; Grasso, C.; Lee, C. Genome-wide detection of alternative splicing in expressed sequences of human genes. Nucleic Acids Res. 2001, 29, 2850–2859. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Manley, J.L. Regulation of pre-mRNA splicing in metazoa. Curr. Opin. Genet. Dev. 1997, 7, 205. [Google Scholar] [CrossRef]
- Cooper, T.A.; Wan, L.; Dreyfuss, G. RNA and disease. Cell 2009, 136, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, N.; Tonegawa, S. Evidence for somatic rearrangement of immunoglobulin genes coding for variable and constant regions. Proc. Natl. Acad. Sci. USA 1976, 73, 3628–3632. [Google Scholar] [CrossRef] [PubMed]
- Tonegawa, S.; Steinberg, C.; Dube, S.; Bernardini, A. Evidence for somatic generation of antibody diversity. Proc. Natl. Acad. Sci. USA 1974, 71, 4027–4031. [Google Scholar] [CrossRef] [PubMed]
- Strasser, A. The role of BH3-only proteins in the immune system. Nat. Rev. Immunol. 2005, 5, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J.; Proudfoot, N.J. Pre-mRNA processing reaches back to transcription and ahead to translation. Cell 2009, 136, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lin, R.J. Spliceosome activation by PRP2 ATPase prior to the first transesterification reaction of pre-mRNA splicing. Mol. Cell. Biol. 1996, 16, 6810–6819. [Google Scholar] [CrossRef] [PubMed]
- Podar, M.; Perlman, P.S.; Padgett, R.A. Stereochemical selectivity of group II intron splicing, reverse splicing, and hydrolysis reactions. Mol. Cell. Biol. 1995, 15, 4466–4478. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.C.; Will, C.L.; Lührmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Caputi, M.; Casari, G.; Guenzi, S.; Tagliabue, R.; Sidoli, A.; Melo, C.A.; Baralle, F.E. A novel bipartite splicing enhancer modulates the differential processing of the human fibronectin EDA exon. Nucleic Acids Res. 1994, 22, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Maniatis, T. Positive control of pre-mRNA splicing in vitro. Science 1992, 256, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Mayeda, A.; Screaton, G.R.; Chandler, S.D.; Fu, X.-D.; Krainer, A.R. Substrate specificities of SR proteins in constitutive splicing are determined by their RNA recognition motifs and composite pre-mRNA exonic elements. Mol. Cell. Biol. 1999, 19, 1853–1863. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Mayeda, A.; Krainer, A.R. Exon identity established through differential antagonism between exonic splicing silencer-bound hnRNP A1 and enhancer-bound SR proteins. Mol. Cell 2001, 8, 1351–1361. [Google Scholar] [CrossRef]
- Cote, G.J.; Stolow, D.T.; Peleg, S.; Berget, S.M.; Gagel, R.F. Identification of exon sequences and an exon binding protein involved in alternative RNA splicing of calcitonin/CGRP. Nucleic Acids Res. 1992, 20, 2361–2366. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Manley, J.L. Mechanisms of alternative splicing regulation: Insights from molecular and genomics approaches. Nat. Rev. Mol. Cell Biol. 2009, 10, 741–754. [Google Scholar] [CrossRef] [PubMed]
- House, A.E.; Lynch, K.W. Regulation of alternative splicing: More than just the ABCs. J. Biol. Chem. 2008, 283, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Motta-Mena, L.B.; Heyd, F.; Lynch, K.W. Context-dependent regulatory mechanism of the splicing factor hnRNP L. Mol. Cell 2010, 37, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.M.; Lynch, K.W. Control of alternative splicing in immune responses: Many regulators, many predictions, much still to learn. Immunol. Rev. 2013, 253, 216–236. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Jia, X.; de la Cruz, L.; Su, X.-C.; Marzolf, B.; Troisch, P.; Zak, D.; Hamilton, A.; Whittle, B.; Yu, D.; et al. Memory T cell RNA rearrangement programmed by heterogeneous nuclear ribonucleoprotein hnRNPLL. Immunity 2008, 29, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Moehle, E.A.; Ryan, C.J.; Krogan, N.J.; Kress, T.L.; Guthrie, C. The yeast SR-like protein Npl3 links chromatin modification to mRNA processing. PLoS Genet. 2012, 8, e1003101. [Google Scholar] [CrossRef] [PubMed]
- Prasad, J.; Colwill, K.; Pawson, T.; Manley, J.L. The protein kinase Clk/Sty directly modulates SR protein activity: Both hyper- and hypophosphorylation inhibit splicing. Mol. Cell. Biol. 1999, 19, 6991–7000. [Google Scholar] [CrossRef] [PubMed]
- Khodor, Y.L.; Rodriguez, J.; Abruzzi, K.C.; Tang, C.H.; Marr, M.T.; Rosbash, M. Nascent-seq indicates widespread cotranscriptional pre-mRNA splicing in Drosophila. Genes Dev. 2011, 25, 2502–2512. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Dufu, K.; Romney, B.; Feldt, M.; Elenko, M.; Reed, R. Functional coupling of RNAP II transcription to spliceosome assembly. Genes Dev. 2006, 20, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Khodor, Y.L.; Menet, J.S.; Tolan, M.; Rosbash, M. Cotranscriptional splicing efficiency differs dramatically between Drosophila and mouse. RNA 2012, 18, 2174–2186. [Google Scholar] [CrossRef] [PubMed]
- Schwerk, C.; Schulze-Osthoff, K. Regulation of apoptosis by alternative pre-mRNA splicing. Mol. Cell 2005, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mayya, V.; Lundgren, D.H.; Hwang, S.I.; Rezaul, K.; Wu, L.; Eng, J.K.; Rodionov, V.; Han, D.K. Quantitative phosphoproteomic analysis of T cell receptor signaling reveals system-wide modulation of protein-protein interactions. Sci. Signal. 2009, 2, ra46. [Google Scholar] [CrossRef] [PubMed]
- Akusjärvi, G.; Kanopka, A.; Estmer, C.; Mühlemann, O.; Petersen-Mahrt, S.; Öhrmalm, C.; Akusjärvi, G. Regulation of adenovirus alternative RNA splicing by dephosphorylation of SR proteins. Nature 1998, 393, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Ip, J.Y.; Tong, A.; Pan, Q.; Topp, J.D.; Blencowe, B.J.; Lynch, K.W. Global analysis of alternative splicing during T-cell activation. RNA 2007, 13, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Mallory, M.J.; Jackson, J.; Weber, B.; Chi, A.; Heyd, F.; Lynch, K.W. Signal- and development-dependent alternative splicing of LEF1 in T cells is controlled by CELF2. Mol. Cell. Biol. 2011, 31, 2184–2195. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.R. The CD45 tyrosine phosphatase: A positive and negative regulator of immune cell function. Semin. Immunol. 2000, 12, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.N.; Jackson, D.I.; Willis, A.C.; Williams, A.F. Lymphocyte specific heterogeneity in the rat leucocyte common antigen (T200) is due to differences in polypeptide sequences near the NH2-terminus. EMBO J. 1987, 6, 1259–1264. [Google Scholar] [PubMed]
- Beverley, P.C.; Daser, A.; Michie, C.A.; Wallace, D.L. Functional subsets of T cells defined by isoforms of CD45. Biochem. Soc. Trans. 1992, 20, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Merkenschlager, M.; Beverley, P.C. Evidence for differential expression of CD45 isoforms by precursors for memory-dependent and independent cytotoxic responses: Human CD8 memory CTLp selectively express CD45RO (UCHL1). Int. Immunol. 1989, 1, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Sieh, M.; Bolen, J.B.; Weiss, A. CD45 specifically modulates binding of Lck to a phosphopeptide encompassing the negative regulatory tyrosine of Lck. EMBO J. 1993, 12, 315–321. [Google Scholar] [PubMed]
- Xu, Z.; Weiss, A. Negative regulation of CD45 by differential homodimerization of the alternatively spliced isoforms. Nat. Immunol. 2002, 3, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Dawes, R.; Petrova, S.; Beverley, P.C.; Tchilian, E.Z. CD45 regulates apoptosis in peripheral T lymphocytes. Int. Immunol. 2006, 18, 959–966. [Google Scholar] [CrossRef] [PubMed]
- McNeill, L.; Cassady, R.L.; Sarkardei, S.; Cooper, J.C.; Morgan, G.; Alexander, D.R. CD45 isoforms in T cell signalling and development. Immunol. Lett. 2004, 92, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Dupéré-Minier, G.; Desharnais, P.; Bernier, J. Involvement of tyrosine phosphatase CD45 in apoptosis. Apoptosis 2010, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hermiston, M.L.; Xu, Z.; Weiss, A. CD45: A critical regulator of signaling thresholds in immune cells. Annu. Rev. Immunol. 2003, 21, 107–137. [Google Scholar] [CrossRef] [PubMed]
- Cyster, J.G.; Healy, J.I.; Kishihara, K.; Mak, T.W.; Thomas, M.L.; Goodnow, C.C. Regulation of B-lymphocyte negative and positive selection by tyrosine phosphatase CD45. Nature 1996, 381, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Beverley, P.C.; Merkenschlager, M.; Terry, L. Phenotypic diversity of the CD45 antigen and its relationship to function. Immunol. Suppl. 1988, 1, 3–5. [Google Scholar] [PubMed]
- Tong, A.; Nguyen, J.; Lynch, K.W. Differential expression of CD45 isoforms is controlled by the combined activity of basal and inducible splicing-regulatory elements in each of the variable exons. J. Biol. Chem. 2005, 280, 38297–38304. [Google Scholar] [CrossRef] [PubMed]
- Rhee, I.; Veillette, A. Protein tyrosine phosphatases in lymphocyte activation and autoimmunity. Nat. Immunol. 2012, 13, 439. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.N.; Terry, L.; Timms, A.; Beverley, P.C.; Janossy, G. Loss of CD45R and gain of UCHL1 reactivity is a feature of primed T cells. J. Immunol. 1988, 140, 2171–2178. [Google Scholar] [PubMed]
- Zikherman, J.; Weiss, A. Alternative splicing of CD45: The tip of the iceberg. Immunity 2008, 29, 839–841. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.W.; Weiss, A. A model system for activation-induced alternative splicing of CD45 pre-mRNA in T cells implicates protein kinase C and Ras. Mol. Cell. Biol. 2000, 20, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Hou, V.C.; Lersch, R.; Gee, S.L.; Ponthier, J.L.; Lo, A.J.; Wu, M.; Turck, C.W.; Koury, M.; Krainer, A.R.; Mayeda, A.; et al. Decrease in hnRNP A/B expression during erythropoiesis mediates a pre-mRNA splicing switch. EMBO J. 2002, 21, 6195–6204. [Google Scholar] [CrossRef] [PubMed]
- Caceres, J.F.; Stamm, S.; Helfman, D.M.; Krainer, A.R. Regulation of alternative splicing in vivo by overexpression of antagonistic splicing factors. Science 1994, 265, 1706–1709. [Google Scholar] [CrossRef] [PubMed]
- Del Gatto, F.; Breathnach, R. Exon and intron sequences, respectively, repress and activate splicing of a fibroblast growth factor receptor 2 alternative exon. Mol. Cell. Biol. 1995, 15, 4825–4834. [Google Scholar] [CrossRef] [PubMed]
- Hui, J.; Hung, L.H.; Heiner, M.; Schreiner, S.; Neumuller, N.; Reither, G.; Haas, S.A.; Bindereif, A. Intronic CA-repeat and CA-rich elements: A new class of regulators of mammalian alternative splicing. EMBO J. 2005, 24, 1988–1998. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.H.; Heiner, M.; Hui, J.; Schreiner, S.; Benes, V.; Bindereif, A. Diverse roles of hnRNP L in mammalian mRNA processing: A combined microarray and RNAi analysis. RNA 2008, 14, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Rothrock, C.R.; House, A.E.; Lynch, K.W. HnRNP L represses exon splicing via a regulated exonic splicing silencer. EMBO J. 2005, 24, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Topp, J.D.; Jackson, J.; Melton, A.A.; Lynch, K.W. A cell-based screen for splicing regulators identifies hnRNPLL as a distinct signal-induced repressor of CD45 variable exon 4. RNA 2008, 14, 2038–2049. [Google Scholar] [CrossRef] [PubMed]
- Gaudreau, M.C.; Heyd, F.; Bastien, R.; Wilhelm, B.; Moroy, T. Alternative splicing controlled by heterogeneous nuclear ribonucleoprotein L regulates development, proliferation, and migration of thymic pre-T cells. J. Immunol. 2012, 188, 5377–5388. [Google Scholar] [CrossRef] [PubMed]
- Oberdoerffer, S.; Moita, L.F.; Neems, D.; Freitas, R.P.; Hacohen, N.; Rao, A. Regulation of CD45 alternative splicing by heterogeneous ribonucleoprotein, hnRNPLL. Science 2008, 321, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Cho, V.; Mei, Y.; Sanny, A.; Chan, S.; Enders, A.; Bertram, E.M.; Tan, A.; Goodnow, C.C.; Andrews, T.D. The RNA-binding protein hnRNPLL induces a T cell alternative splicing program delineated by differential intron retention in polyadenylated RNA. Genome Biol. 2014, 15, R26. [Google Scholar] [CrossRef] [PubMed]
- Heyd, F.; Lynch, K.W. Phosphorylation-dependent regulation of PSF by GSK3 controls CD45 alternative splicing. Mol. Cell 2010, 40, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Melton, A.A.; Jackson, J.; Wang, J.; Lynch, K.W. Combinatorial control of signal-induced exon repression by hnRNP L and PSF. Mol. Cell. Biol. 2007, 27, 6972–6984. [Google Scholar] [CrossRef] [PubMed]
- Moulton, V.R.; Tsokos, G.C. Alternative splicing factor/splicing factor 2 regulates the expression of the ζ subunit of the human T cell receptor-associated CD3 complex. J. Biol. Chem. 2010, 285, 12490–12496. [Google Scholar] [CrossRef] [PubMed]
- Lukong, K.E.; Richard, S. SAM68, the KH domain-containing superSTAR. Biochim. Biophys. Acta 2003, 1653, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Boise, L.H.; Gonzalez-Garcia, M.; Postema, C.E.; Ding, L.; Lindsten, T.; Turka, L.A.; Mao, X.; Nunez, G.; Thompson, C.B. Bcl-X, a Bcl-2-related gene that functions as a dominant regulator of apoptotic cell death. Cell 1993, 74, 597–608. [Google Scholar] [CrossRef]
- Matter, N.; Herrlich, P.; Konig, H. Signal-dependent regulation of splicing via phosphorylation of Sam68. Nature 2002, 420, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Arch, R.; Wirth, K.; Hofmann, M.; Ponta, H.; Matzku, S.; Herrlich, P.; Zoller, M. Participation in normal immune responses of a metastasis-inducing splice variant of CD44. Science 1992, 257, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.L.; Dougherty, G.J. To metastasize or not? Selection of CD44 splice sites. Nat. Med. 1995, 1, 635–637. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.; Sleeman, J.; Dall, P.; Hekele, A.; Moll, J.; Ponta, H.; Herrlich, P. The CD44 proteins in embryonic development and in cancer. Curr. Top. Microbiol. Immunol. 1996, 213, 249–269. [Google Scholar] [PubMed]
- Sherman, L.; Wainwright, D.; Ponta, H.; Herrlich, P. A splice variant of CD44 expressed in the apical ectodermal ridge presents fibroblast growth factors to limb mesenchyme and is required for limb outgrowth. Genes Dev. 1998, 12, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Weg-Remers, S.; Ponta, H.; Herrlich, P.; Konig, H. Regulation of alternative pre-mRNA splicing by the ERK MAP-kinase pathway. EMBO J. 2001, 20, 4194–4203. [Google Scholar] [CrossRef] [PubMed]
- Batsche, E.; Yaniv, M.; Muchardt, C. The human SWI/SNF subunit Brm is a regulator of alternative splicing. Nat. Struct. Mol. Biol. 2006, 13, 22–29. [Google Scholar] [CrossRef] [PubMed]
- de la Mata, M.; Alonso, C.R.; Kadener, S.; Fededa, J.P.; Blaustein, M.; Pelisch, F.; Cramer, P.; Bentley, D.; Kornblihtt, A.R. A slow RNA polymerase II affects alternative splicing in vivo. Mol. Cell 2003, 12, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.J.; Kane, C.M.; Ares, M., Jr. Perturbation of transcription elongation influences the fidelity of internal exon inclusion in Saccharomyces cerevisiae. RNA 2003, 9, 993–1006. [Google Scholar] [CrossRef] [PubMed]
- Paronetto, M.P.; Achsel, T.; Massiello, A.; Chalfant, C.E.; Sette, C. The RNA-binding protein Sam68 modulates the alternative splicing of Bcl-X. J. Cell Biol. 2007, 176, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Boise, L.H.; Thompson, C.B. Bcl-XL can inhibit apoptosis in cells that have undergone Fas-induced protease activation. Proc. Natl. Acad. Sci. USA 1997, 94, 3759–3764. [Google Scholar] [CrossRef] [PubMed]
- Busa, R.; Sette, C. An emerging role for nuclear RNA-mediated responses to genotoxic stress. RNA Biol. 2010, 7, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Bielli, P.; Busa, R.; Di Stasi, S.M.; Munoz, M.J.; Botti, F.; Kornblihtt, A.R.; Sette, C. The transcription factor FBI-1 inhibits SAM68-mediated Bcl-X alternative splicing and apoptosis. EMBO Rep. 2014, 15, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornblihtt, A.R. Coupling transcription and alternative splicing. Adv. Exp. Med. Biol. 2007, 623, 175–189. [Google Scholar] [PubMed]
- Shukla, S.; Kavak, E.; Gregory, M.; Imashimizu, M.; Shutinoski, B.; Kashlev, M.; Oberdoerffer, P.; Sandberg, R.; Oberdoerffer, S. CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature 2011, 479, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Iseni, F.; Garcin, D.; Nishio, M.; Kedersha, N.; Anderson, P.; Kolakofsky, D. Sendai virus trailer RNA binds TIAR, a cellular protein involved in virus-induced apoptosis. EMBO J. 2002, 21, 5141–5150. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Streuli, M.; Saito, H.; Schlossman, S.F.; Anderson, P. A polyadenylate binding protein localized to the granules of cytolytic lymphocytes induces DNA fragmentation in target cells. Cell 1991, 67, 629–639. [Google Scholar] [CrossRef]
- Piecyk, M.; Wax, S.; Beck, A.R.; Kedersha, N.; Gupta, M.; Maritim, B.; Chen, S.; Gueydan, C.; Kruys, V.; Streuli, M.; Anderson, P. TIA-1 is a translational silencer that selectively regulates the expression of TNF-α. EMBO J. 2000, 19, 4154–4163. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, J.M.; Majos, N.; Bonnal, S.; Martinez, C.; Castelo, R.; Guigo, R.; Bilbao, D.; Valcarcel, J. Regulation of Fas alternative splicing by antagonistic effects of TIA-1 and PTB on exon definition. Mol. Cell 2005, 19, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Cascino, I.; Fiucci, G.; Papoff, G.; Ruberti, G. Three functional soluble forms of the human apoptosis-inducing FAS molecule are produced by alternative splicing. J. Immunol. 1995, 154, 2706–2713. [Google Scholar] [PubMed]
- Cheng, J.; Zhou, T.; Liu, C.; Shapiro, J.P.; Brauer, M.J.; Kiefer, M.C.; Barr, P.J.; Mountz, J.D. Protection from FAS-mediated apoptosis by a soluble form of the FAS molecule. Science 1994, 263, 1759–1762. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, J.M.; Valcarcel, J. FAS-activated serine/threonine kinase (FAST K) synergizes with TIA-1/TIAR proteins to regulate Fas alternative splicing. J. Biol. Chem. 2007, 282, 1539–1543. [Google Scholar] [CrossRef] [PubMed]
- Goodnow, C.C.; Sprent, J.; Fazekas de St Groth, B.; Vinuesa, C.G. Cellular and genetic mechanisms of self tolerance and autoimmunity. Nature 2005, 435, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Hogquist, K.A.; Jameson, S.C.; Bevan, M.J. Strong agonist ligands for the T cell receptor do not mediate positive selection of functional CD8+ T cells. Immunity 1995, 3, 79–86. [Google Scholar] [CrossRef]
- Jameson, S.C.; Bevan, M.J. T cell receptor antagonists and partial agonists. Immunity 1995, 2, 1–11. [Google Scholar] [CrossRef]
- Zikherman, J.; Au-Yeung, B. The role of T cell receptor signaling thresholds in guiding T cell fate decisions. Curr. Opin. Immunol. 2015, 33, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Flavell, R.A. The relationship of inflammation and initiation of autoimmune disease: Role of TNF super family members. Curr. Top. Microbiol. Immunol. 2002, 266, 1–9. [Google Scholar] [PubMed]
- Cascino, I.; Papoff, G.; de Maria, R.; Testi, R.; Ruberti, G. FAS/Apo-1 (CD95) receptor lacking the intracytoplasmic signaling domain protects tumor cells from FAS-mediated apoptosis. J. Immunol. 1996, 156, 13–17. [Google Scholar] [PubMed]
- Hughes, D.P.; Crispe, I.N. A naturally occurring soluble isoform of murine Fas generated by alternative splicing. J. Exp. Med. 1995, 182, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.H.; Zhang, W.J.; Rao, Y.; Wu, J.Y. Regulation of Ich-1 pre-mRNA alternative splicing and apoptosis by mammalian splicing factors. Proc. Natl. Acad. Sci. USA 1998, 95, 9155–9160. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Chen, G.G.; Yun, J.P.; Chun, S.Y.; Zheng, Z.Z.; Ho, R.L.; Chak, E.C.; Xia, N.S.; Lai, P.B. Identification and characterization of BH3 domain protein Bim and its isoforms in human hepatocellular carcinomas. Apoptosis 2007, 12, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Chalfant, C.E.; Ogretmen, B.; Galadari, S.; Kroesen, B.J.; Pettus, B.J.; Hannun, Y.A. FAS activation induces dephosphorylation of SR proteins; dependence on the de novo generation of ceramide and activation of protein phosphatase 1. J. Biol. Chem. 2001, 276, 44848–44855. [Google Scholar] [CrossRef] [PubMed]
- Bouillet, P.; Purton, J.F.; Godfrey, D.I.; Zhang, L.C.; Coultas, L.; Puthalakath, H.; Pellegrini, M.; Cory, S.; Adams, J.M.; Strasser, A. BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. Nature 2002, 415, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-W.; Chandra, D.; Tang, S.-H.; Chopra, D.; Tang, D.G. Identification and characterization of Bimγ, a novel proapoptotic BH3-only splice variant of Bim. Cancer Res. 2002, 62, 2976. [Google Scholar] [PubMed]
- O’Connor, L.; Strasser, A.; O’Reilly, L.A.; Hausmann, G.; Adams, J.M.; Cory, S.; Huang, D.C. Bim: A novel member of the Bcl-2 family that promotes apoptosis. EMBO J. 1998, 17, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Bouillet, P.; Metcalf, D.; Huang, D.C.; Tarlinton, D.M.; Kay, T.W.; Kontgen, F.; Adams, J.M.; Strasser, A. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. Science 1999, 286, 1735–1738. [Google Scholar] [CrossRef] [PubMed]
- Anczukow, O.; Rosenberg, A.Z.; Akerman, M.; Das, S.; Zhan, L.; Karni, R.; Muthuswamy, S.K.; Krainer, A.R. The splicing factor SRSF1 regulates apoptosis and proliferation to promote mammary epithelial cell transformation. Nat. Struct. Mol. Biol. 2012, 19, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Juan, W.C.; Roca, X.; Ong, S.T. Identification of cis-acting elements and splicing factors involved in the regulation of Bim pre-mRNA splicing. PLoS ONE 2014, 9, e95210. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Takeda, T.; Yamamoto, N.; Furuya, K.; Hirose, K.; Kamiya, T.; Adachi, T. Zinc-induced modulation of SRSF6 activity alters Bim splicing to promote generation of the most potent apoptotic isoform BimS. FEBS J. 2013, 280, 3313–3327. [Google Scholar] [CrossRef] [PubMed]
- Chikuma, S.; Imboden, J.B.; Bluestone, J.A. Negative regulation of T cell receptor-lipid raft interaction by cytotoxic T lymphocyte-associated antigen 4. J. Exp. Med. 2003, 197, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, J.J.; Sullivan, T.J.; Allison, J.P. CTLA-4 overexpression inhibits T cell responses through a CD28-B7-dependent mechanism. J. Immunol. 2006, 177, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Walunas, T.L.; Bakker, C.Y.; Bluestone, J.A. CTLA-4 ligation blocks CD28-dependent T cell activation. J. Exp. Med. 1996, 183, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Walunas, T.L.; Lenschow, D.J.; Bakker, C.Y.; Linsley, P.S.; Freeman, G.J.; Green, J.M.; Thompson, C.B.; Bluestone, J.A. CTLA-4 can function as a negative regulator of T cell activation. Immunity 1994, 1, 405–413. [Google Scholar] [CrossRef]
- Butte, M.J.; Lee, S.J.; Jesneck, J.; Keir, M.E.; Haining, W.N.; Sharpe, A.H. CD28 costimulation regulates genome-wide effects on alternative splicing. PLoS ONE 2012, 7, e40032. [Google Scholar] [CrossRef] [PubMed]
- Brunner, M.C.; Chambers, C.A.; Chan, F.K.; Hanke, J.; Winoto, A.; Allison, J.P. CTLA-4-Mediated inhibition of early events of T cell proliferation. J. Immunol. 1999, 162, 5813–5820. [Google Scholar] [PubMed]
- Chambers, C.A.; Sullivan, T.J.; Allison, J.P. Lymphoproliferation in CTLA-4-deficient mice is mediated by costimulation-dependent activation of CD4+ T cells. Immunity 1997, 7, 885–895. [Google Scholar] [CrossRef]
- Krummel, M.F.; Allison, J.P. CTLA-4 engagement inhibits IL-2 accumulation and cell cycle progression upon activation of resting T cells. J. Exp. Med. 1996, 183, 2533–2540. [Google Scholar] [CrossRef] [PubMed]
- Kantarci, O.H.; Hebrink, D.D.; Achenbach, S.J.; Atkinson, E.J.; Waliszewska, A.; Buckle, G.; McMurray, C.T.; de Andrade, M.; Hafler, D.A.; Weinshenker, B.G. CTLA4 is associated with susceptibility to multiple sclerosis. J. Neuroimmunol. 2003, 134, 133–141. [Google Scholar] [CrossRef]
- Nithiyananthan, R.; Heward, J.M.; Allahabadia, A.; Franklyn, J.A.; Gough, S.C. Polymorphism of the CTLA-4 gene is associated with autoimmune hypothyroidism in the United Kingdom. Thyroid 2002, 12, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Howson, J.M.; Esposito, L.; Heward, J.; Snook, H.; Chamberlain, G.; Rainbow, D.B.; Hunter, K.M.; Smith, A.N.; di Genova, G.; et al. Association of the T-cell regulatory gene CTLA4 with susceptibility to autoimmune disease. Nature 2003, 423, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Vijayakrishnan, L.; Slavik, J.M.; Illes, Z.; Greenwald, R.J.; Rainbow, D.; Greve, B.; Peterson, L.B.; Hafler, D.A.; Freeman, G.J.; Sharpe, A.H.; et al. An autoimmune disease-associated CTLA-4 splice variant lacking the B7 binding domain signals negatively in T cells. Immunity 2004, 20, 563–575. [Google Scholar] [CrossRef]
- Surh, C.D.; Sprent, J. Homeostasis of naive and memory T cells. Immunity 2008, 29, 848–862. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.G.; Schmidt, S.; Seth, P.; Oksenberg, J.R.; Hart, J.; Prokop, A.; Caillier, S.J.; Ban, M.; Goris, A.; Barcellos, L.F.; et al. Multiple Sclerosis Genetics, G., Interleukin 7 receptor α chain (IL7R) shows allelic and functional association with multiple sclerosis. Nat. Genet. 2007, 39, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- McKay, F.C.; Swain, L.I.; Schibeci, S.D.; Rubio, J.P.; Kilpatrick, T.J.; Heard, R.N.; Stewart, G.J.; Booth, D.R. CD127 immunophenotyping suggests altered CD4+ T cell regulation in primary progressive multiple sclerosis. J. Autoimmun. 2008, 31, 52–58. [Google Scholar] [CrossRef] [PubMed]
- McKay, F.C.; Swain, L.I.; Schibeci, S.D.; Rubio, J.P.; Kilpatrick, T.J.; Heard, R.N.; Stewart, G.J.; Booth, D.R. Haplotypes of the interleukin 7 receptor α gene are correlated with altered expression in whole blood cells in multiple sclerosis. Genes Immun. 2008, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vudattu, N.K.; Magalhaes, I.; Hoehn, H.; Pan, D.; Maeurer, M.J. Expression analysis and functional activity of interleukin-7 splice variants. Genes Immun. 2009, 10, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Nelms, K.A.; Goodnow, C.C. Genome-wide ENU mutagenesis to reveal immune regulators. Immunity 2001, 15, 409–418. [Google Scholar] [CrossRef]
- Wu, Z.; Yates, A.L.; Hoyne, G.F.; Goodnow, C.C. Consequences of increased CD45RA and RC isoforms for TCR signaling and peripheral T cell deficiency resulting from heterogeneous nuclear ribonucleoprotein L-like mutation. J. Immunol. 2010, 185, 231. [Google Scholar] [CrossRef] [PubMed]
- Yabas, M.; Godfrey, D.I.; Goodnow, C.C.; Hoyne, G.F. Differential requirement for the CD45 splicing regulator hnRNPLL for accumulation of NKT and conventional T cells. PLoS ONE 2011, 6, e26440. [Google Scholar] [CrossRef] [PubMed]
- McNeill, L.; Salmond, R.J.; Cooper, J.C.; Carret, C.K.; Cassady-Cain, R.L.; Roche-Molina, M.; Tandon, P.; Holmes, N.; Alexander, D.R. The differential regulation of Lck kinase phosphorylation sites by CD45 is critical for T cell receptor signaling responses. Immunity 2007, 27, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Nutt, S.L.; Kee, B.L. The transcriptional regulation of B cell lineage commitment. Immunity 2007, 26, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.R.; Mandal, M.; Ochiai, K.; Singh, H. Orchestrating B cell lymphopoiesis through interplay of IL-7 receptor and pre-B cell receptor signalling. Nat. Rev. Immunol. 2014, 14, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Gay, D.; Saunders, T.; Camper, S.; Weigert, M. Receptor editing: An approach by autoreactive B cells to escape tolerance. J. Exp. Med. 1993, 177, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Tiegs, S.L.; Russell, D.M.; Nemazee, D. Receptor editing in self-reactive bone marrow B cells. J. Exp. Med. 1993, 177, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Radic, M.Z.; Erikson, J.; Litwin, S.; Weigert, M. B lymphocytes may escape tolerance by revising their antigen receptors. J. Exp. Med. 1993, 177, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Halverson, R.; Torres, R.M.; Pelanda, R. Receptor editing is the main mechanism of B cell tolerance toward membrane antigens. Nat. Immunol. 2004, 5, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Nemazee, D.A.; Burki, K. Clonal deletion of B lymphocytes in a transgenic mouse bearing anti-MHC class I antibody genes. Nature 1989, 337, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Goodnow, C.C.; Crosbie, J.; Adelstein, S.; Lavoie, T.B.; Smith-Gill, S.J.; Brink, R.A.; Pritchard-Briscoe, H.; Wotherspoon, J.S.; Loblay, R.H.; Raphael, K.; et al. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature 1988, 334, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Loder, F.; Mutschler, B.; Ray, R.J.; Paige, C.J.; Sideras, P.; Torres, R.; Lamers, M.C.; Carsetti, R. B cell development in the spleen takes place in discrete steps and is determined by the quality of B cell receptor-derived signals. J. Exp. Med. 1999, 190, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.W.; Rogers, J.; Hunkapiller, T.; Early, P.; Nottenburg, C.; Weissman, I.; Bazin, H.; Wall, R.; Hood, L.E. Expression of IgD may use both DNA rearrangement and RNA splicing mechanisms. Proc. Natl. Acad. Sci. USA 1981, 78, 1800–1804. [Google Scholar] [CrossRef] [PubMed]
- Maki, R.; Roeder, W.; Traunecker, A.; Sidman, C.; Wabl, M.; Raschke, W.; Tonegawa, S. The role of DNA rearrangement and alternative RNA processing in the expression of immunoglobulin δ genes. Cell 1981, 24, 353–365. [Google Scholar] [CrossRef]
- Ohta, Y.; Flajnik, M. IgD, like IgM, is a primordial immunoglobulin class perpetuated in most jawed vertebrates. Proc. Natl. Acad. Sci. USA 2006, 103, 10723–10728. [Google Scholar] [CrossRef] [PubMed]
- Enders, A.; Short, A.; Miosge, L.A.; Bergmann, H.; Sontani, Y.; Bertram, E.M.; Whittle, B.; Balakishnan, B.; Yoshida, K.; Sjollema, G.; et al. Zinc-finger protein ZFP318 is essential for expression of IgD, the alternatively spliced Igh product made by mature B lymphocytes. Proc. Natl. Acad. Sci. USA 2014, 111, 4513–4518. [Google Scholar] [CrossRef] [PubMed]
- Pioli, P.D.; Debnath, I.; Weis, J.J.; Weis, J.H. Zfp318 regulates IgD expression by abrogating transcription termination within the Ighm/Ighd locus. J. Immunol. 2014, 193, 2546–2553. [Google Scholar] [CrossRef] [PubMed]
- Heng, T.S.; Painter, M.W. The Immunological Genome Project: Networks of gene expression in immune cells. Nat. Immunol. 2008, 9, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Lamson, G.; Koshland, M.E. Changes in J chain and mu chain RNA expression as a function of B cell differentiation. J. Exp. Med. 1984, 160, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Alt, F.W.; Bothwell, A.L.; Knapp, M.; Siden, E.; Mather, E.; Koshland, M.; Baltimore, D. Synthesis of secreted and membrane-bound immunoglobulin mu heavy chains is directed by mRNAs that differ at their 3′ ends. Cell 1980, 20, 293–301. [Google Scholar] [CrossRef]
- Rogers, J.; Early, P.; Carter, C.; Calame, K.; Bond, M.; Hood, L.; Wall, R. Two mRNAs with different 3′ ends encode membrane-bound and secreted forms of immunoglobulin mu chain. Cell 1980, 20, 303–312. [Google Scholar] [CrossRef]
- Early, P.; Rogers, J.; Davis, M.; Calame, K.; Bond, M.; Wall, R.; Hood, L. Two mRNAs can be produced from a single immunoglobulin mu gene by alternative RNA processing pathways. Cell 1980, 20, 313–319. [Google Scholar] [CrossRef]
- Benson, M.J.; Aijo, T.; Chang, X.; Gagnon, J.; Pape, U.J.; Anantharaman, V.; Aravind, L.; Pursiheimo, J.P.; Oberdoerffer, S.; Liu, X.S.; et al. Heterogeneous nuclear ribonucleoprotein L-like (hnRNPLL) and elongation factor, RNA polymerase II, 2 (ELL2) are regulators of mRNA processing in plasma cells. Proc. Natl. Acad. Sci. USA 2012, 109, 16252–16257. [Google Scholar] [CrossRef] [PubMed]
- Martincic, K.; Alkan, S.A.; Cheatle, A.; Borghesi, L.; Milcarek, C. Transcription elongation factor ELL2 directs immunoglobulin secretion in plasma cells by stimulating altered RNA processing. Nat. Immunol. 2009, 10, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Bayles, I.; Szlachta-McGinn, A.; Paul, J.; Boiko, J.; Santos, P.; Liu, J.; Wang, Z.; Borghesi, L.; Milcarek, C. Transcription elongation factor ELL2 drives Ig secretory-specific mRNA production and the unfolded protein response. J. Immunol. 2014, 193, 4663–4674. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Li, B.; Rao, A. RNA-binding protein hnRNPLL regulates mRNA splicing and stability during B-cell to plasma-cell differentiation. Proc. Natl. Acad. Sci. USA 2015, 112, E1888–E1897. [Google Scholar] [CrossRef] [PubMed]
- Hathcock, K.S.; Hirano, H.; Murakami, S.; Hodes, R.J. CD45 expression by B cells. Expression of different CD45 isoforms by subpopulations of activated B cells. J. Immunol. 1992, 149, 2286–2294. [Google Scholar] [PubMed]
- Ogimoto, M.; Katagiri, T.; Hasegawa, K.; Mizuno, K.; Yakura, H. Induction of CD45 isoform switch in murine B cells by antigen receptor stimulation and by phorbol myristate acetate and ionomycin. Cell Immunol. 1993, 151, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Basso, K.; Dalla-Favera, R. BCL6: Master regulator of the germinal center reaction and key oncogene in B cell lymphomagenesis. Adv. Immunol. 2010, 105, 193–210. [Google Scholar] [PubMed]
- Klein, U.; Casola, S.; Cattoretti, G.; Shen, Q.; Lia, M.; Mo, T.; Ludwig, T.; Rajewsky, K.; Dalla-Favera, R. Transcription factor IRF4 controls plasma cell differentiation and class-switch recombination. Nat. Immunol. 2006, 7, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Wilton, S.D.; Fletcher, S. RNA splicing manipulation: Strategies to modify gene expression for a variety of therapeutic outcomes. Curr. Gene Ther. 2011, 11, 259–275. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yabas, M.; Elliott, H.; Hoyne, G.F. The Role of Alternative Splicing in the Control of Immune Homeostasis and Cellular Differentiation. Int. J. Mol. Sci. 2016, 17, 3. https://doi.org/10.3390/ijms17010003
Yabas M, Elliott H, Hoyne GF. The Role of Alternative Splicing in the Control of Immune Homeostasis and Cellular Differentiation. International Journal of Molecular Sciences. 2016; 17(1):3. https://doi.org/10.3390/ijms17010003
Chicago/Turabian StyleYabas, Mehmet, Hannah Elliott, and Gerard F. Hoyne. 2016. "The Role of Alternative Splicing in the Control of Immune Homeostasis and Cellular Differentiation" International Journal of Molecular Sciences 17, no. 1: 3. https://doi.org/10.3390/ijms17010003
APA StyleYabas, M., Elliott, H., & Hoyne, G. F. (2016). The Role of Alternative Splicing in the Control of Immune Homeostasis and Cellular Differentiation. International Journal of Molecular Sciences, 17(1), 3. https://doi.org/10.3390/ijms17010003