
The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on Alcohol-Induced Liver Injury

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
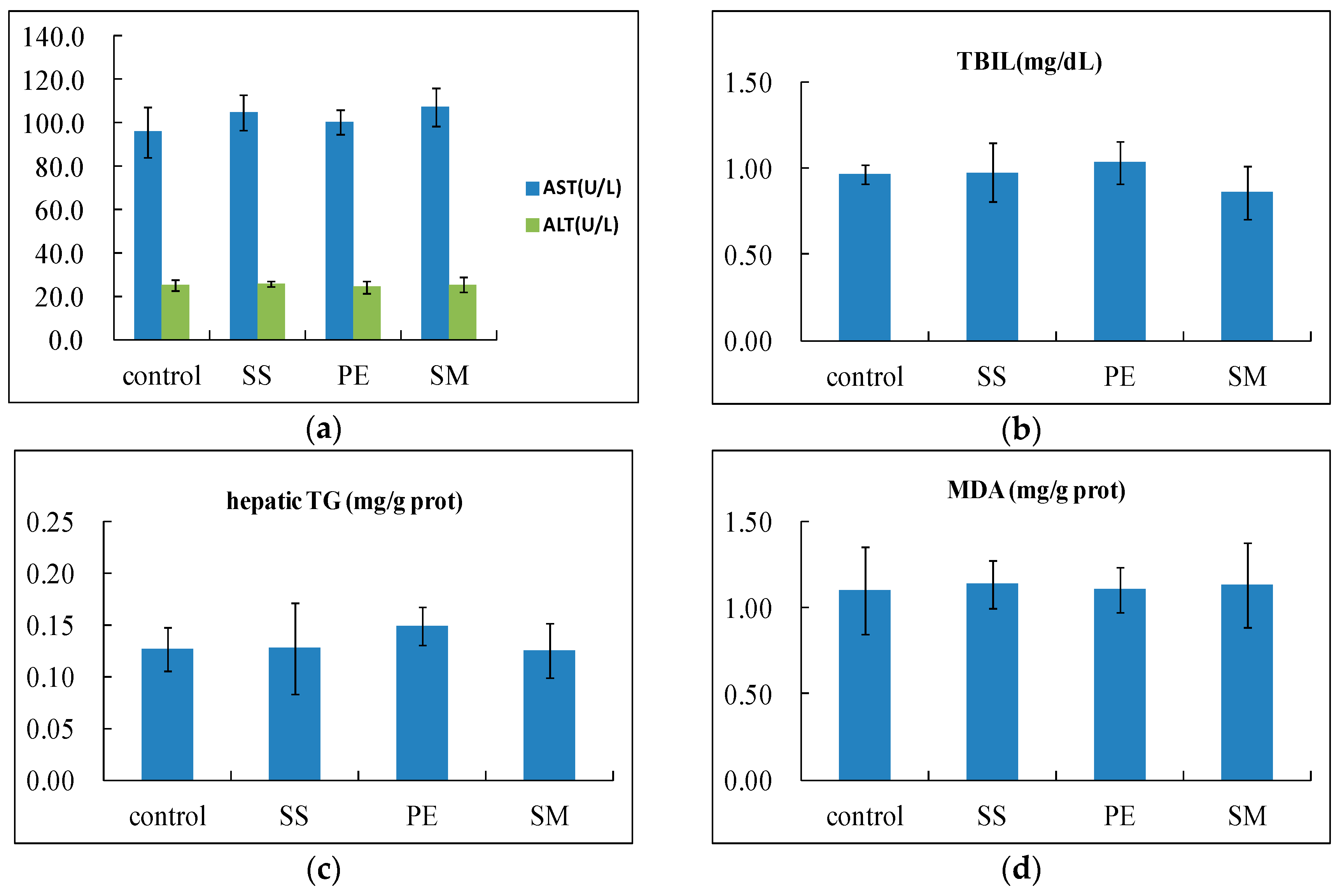{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
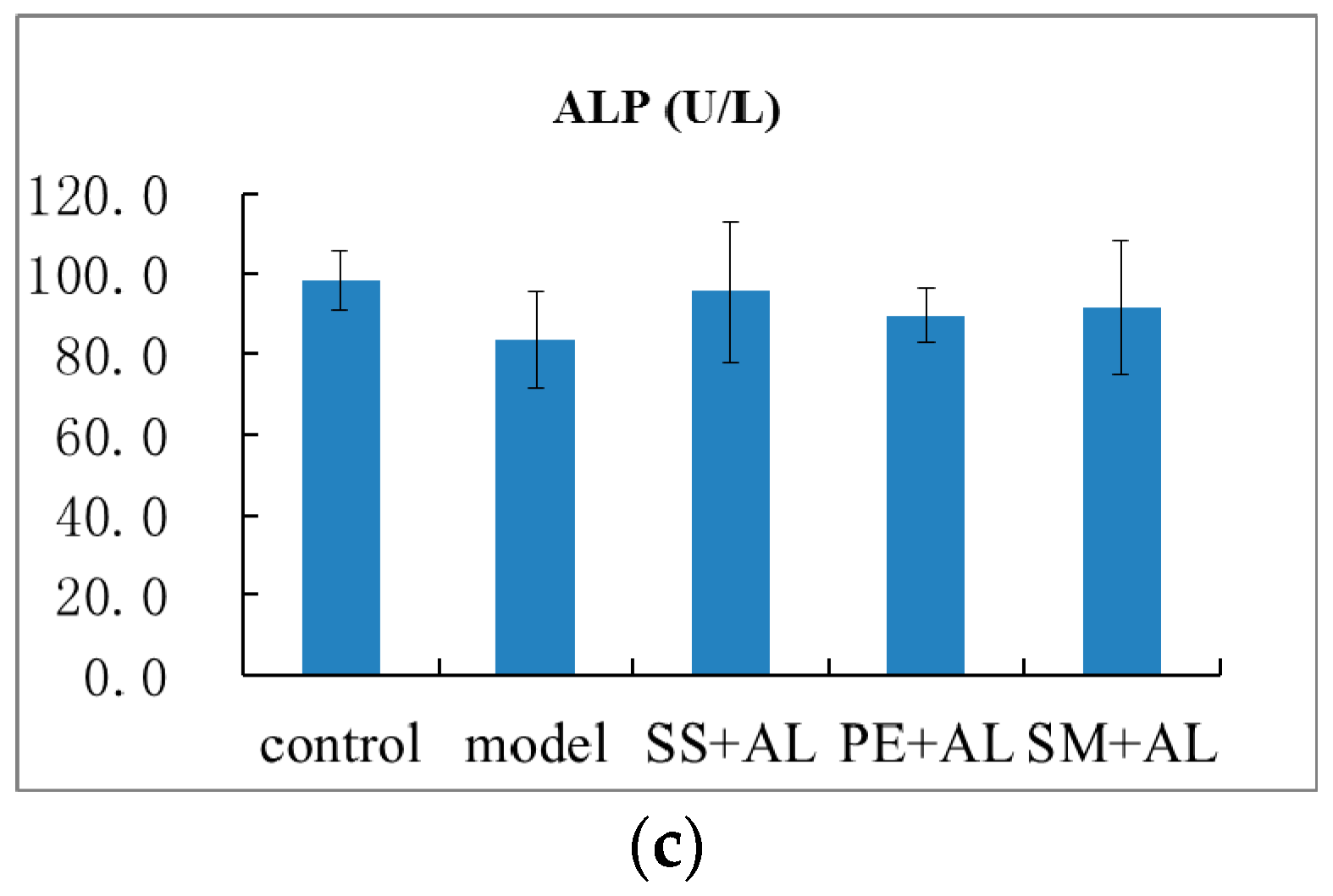
2.1. The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on the Levels of Aspartate Transaminase (AST), Alanine Transaminase (ALT) and Alkaline Phosphatase (ALP) in Serum
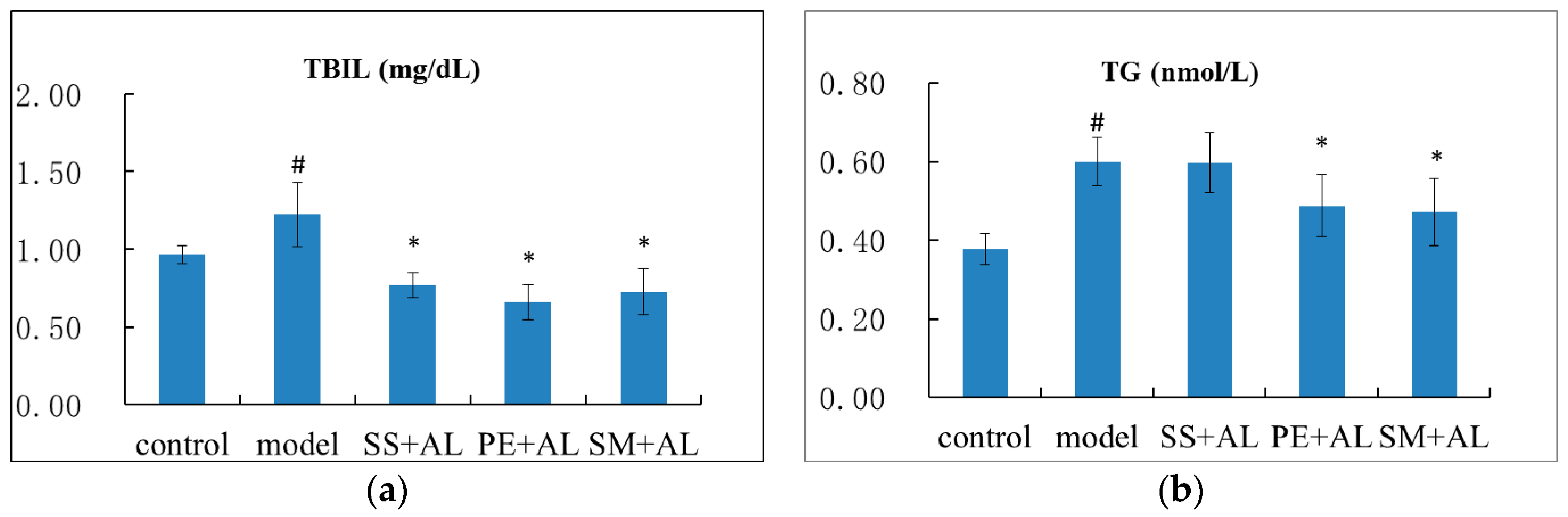
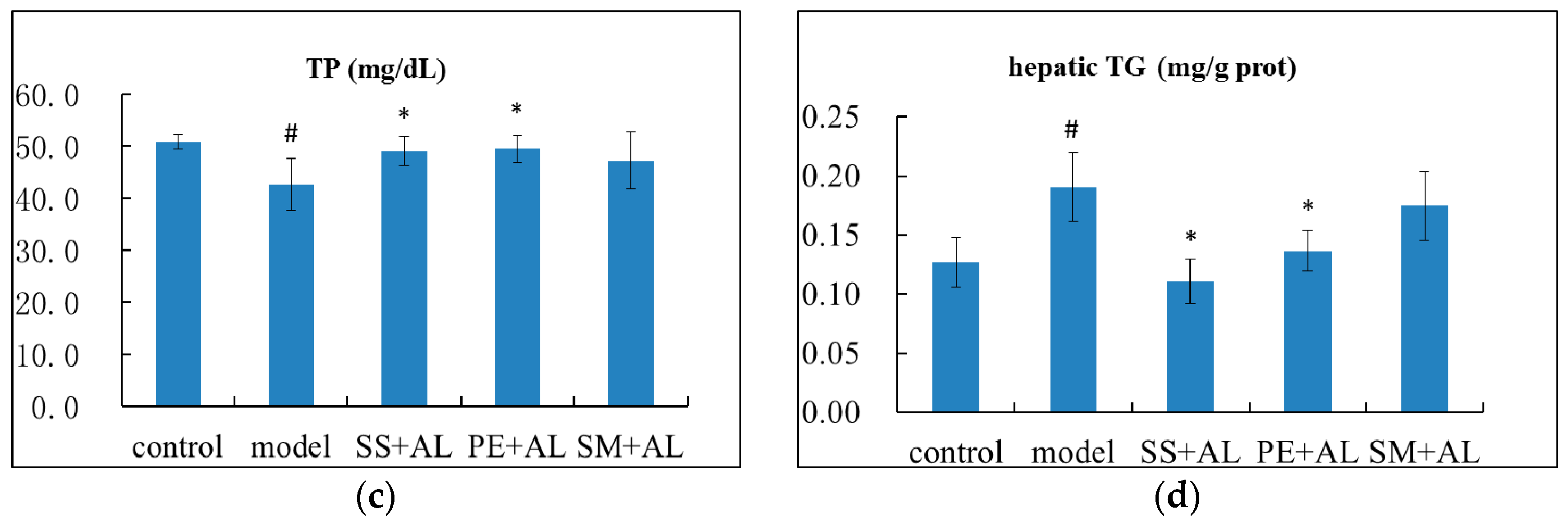
2.2. The Effects of Syzygium samarangense, Passiflora edulis, and Solanum muricatum on the Levels of Serum Triglyceride (TG), Total Protein (TP), Total Bilirubin (TBIL), and Hepatic TG
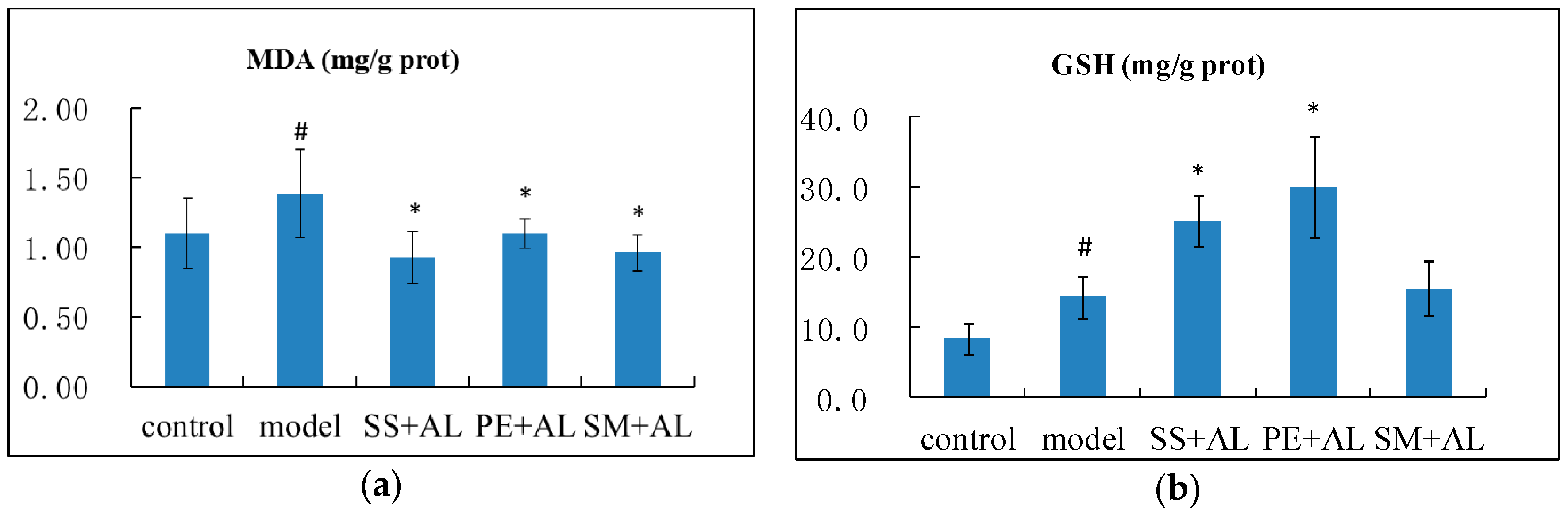
2.3. The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on the Contents of Malondialdehyde (MDA) and Glutathione (GSH) in the Liver
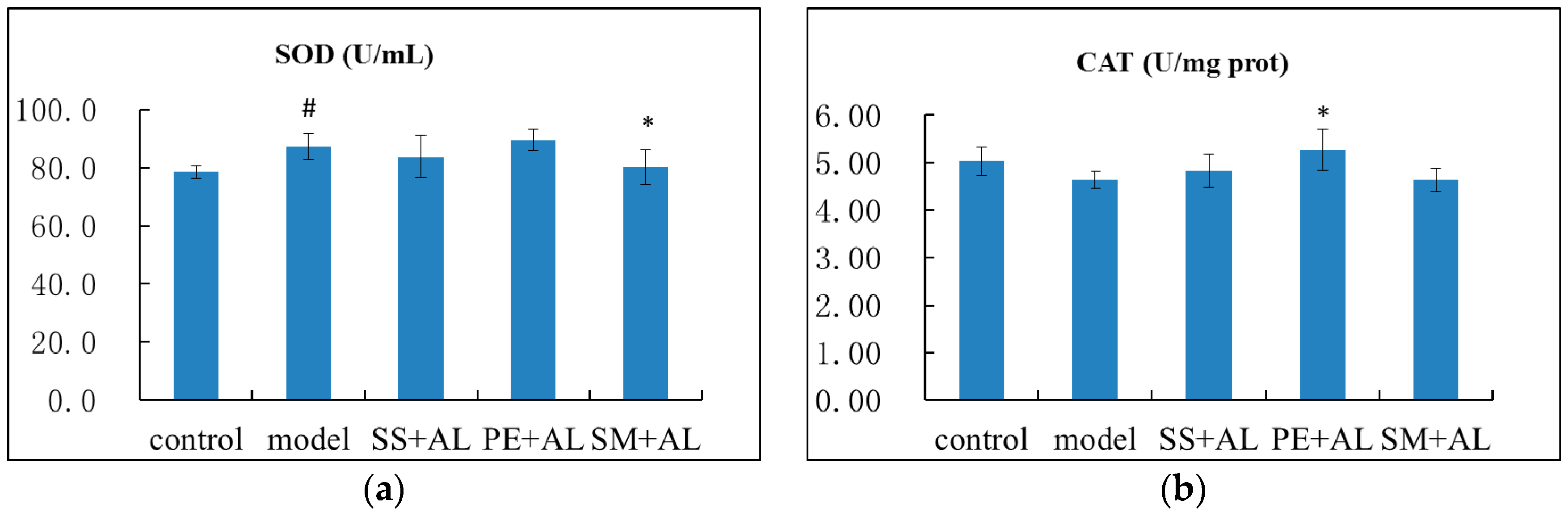
2.4. The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on the Levels of Hepatic Superoxide Dismutase (SOD) and Catalase (CAT)
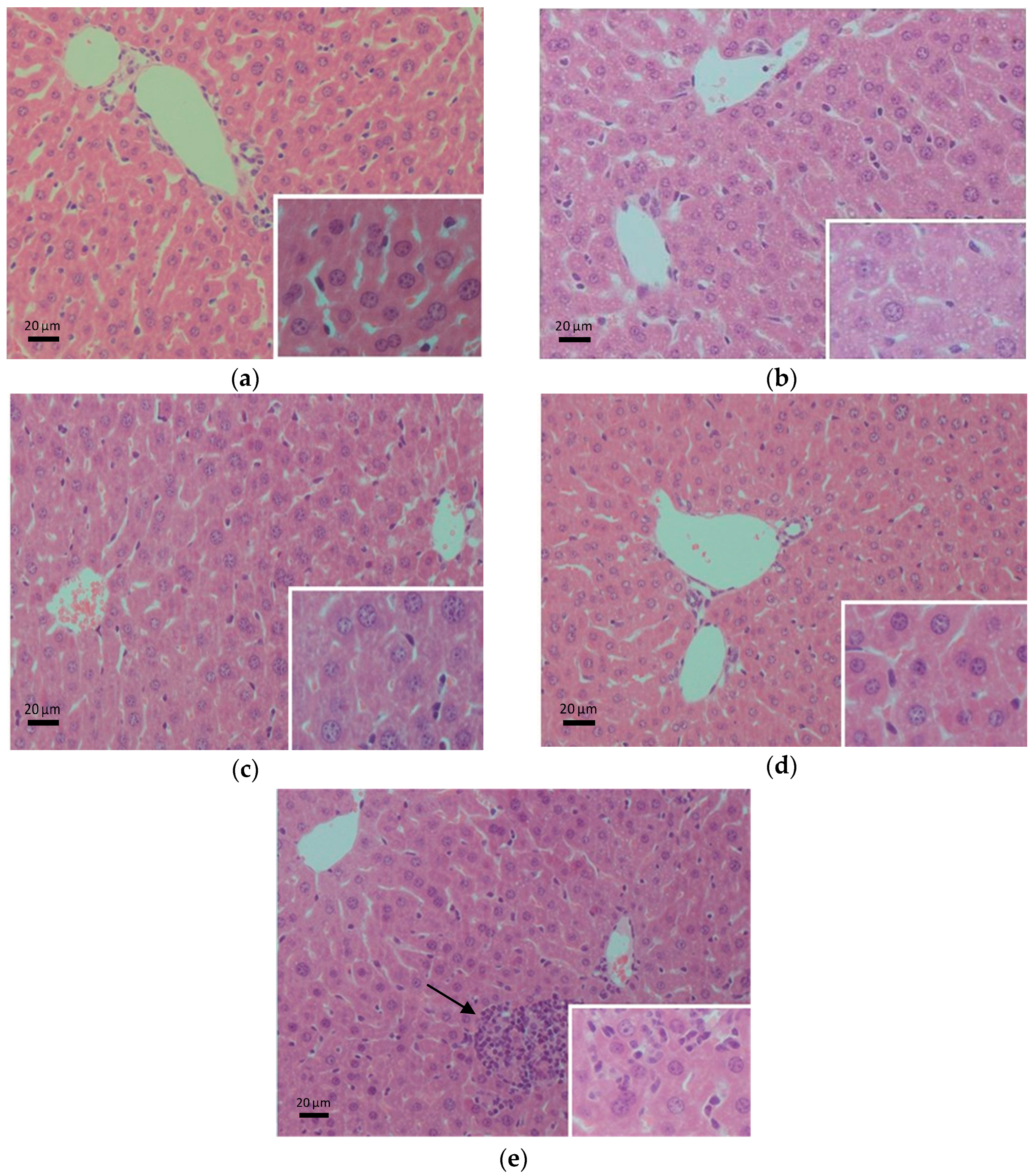
2.5. Histopathological Evaluation
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Fruit Materials
3.3. Animal Study
3.4. Measurement of Hepatic Injury in the Serum
3.5. Measurement of MDA, SOD, GSH, CAT, and TG in the Liver
3.6. Liver Histopathological Evaluation
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sung, C.; Kim, S.; Oh, C.; Yang, S.; Han, B.; Mo, E. Taraxerone enhances alcohol oxidation via increases of alcohol dehyderogenase (ADH) and acetaldehyde dehydrogenase (ALDH) activities and gene expressions. Food Chem. Toxicol. 2012, 50, 2508–2514. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, S.; Zhou, T.; Zhang, P.; Li, H.B. Alcoholic beverage consumption and chronic diseases. Int. J. Environ. Res. Public Health 2016. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Cai, S.; Guo, J.; Zhu, Y.; Li, M.; Yu, Y.; Zhang, S.; Chen, K. Alcohol drinking and all cancer mortality: A meta-analysis. Ann. Oncol. 2013, 24, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Ramchandani, V.A.; Bosron, W.F.; Li, T.K. Research advances in ethanol metabolism. Pathol. Biol. 2001, 49, 676–682. [Google Scholar] [CrossRef]
- Bourogaa, E.; Nciri, R.; Mezghani-Jarraya, R.; Racaud-Sultan, C.; Damak, M.; El Feki, A. Antioxidant activity and hepatoprotective potential of Hammada scoparia against ethanol-induced liver injury in rats. J. Physiol. Biochem. 2013, 69, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Hasumura, Y.; Joly, J.G.; Lieber, C.S. Microsomal ethanol-oxidizing system (MEOS): Purification and properties of a rat liver system free of catalase and alcohol dehydrogenase. Biochem. Biophys. Res. Commun. 1972, 49, 1187–1193. [Google Scholar] [CrossRef]
- Teschke, R.; Gellert, J. Hepatic microsomal ethanol-oxidizing system (MEOS): Metabolic aspects and clinical implications. Alcohol Clin. Exp. Res. 1986, 10, 20S–32S. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Hasumura, Y.; Lieber, C.S. Hepatic microsomal alcohol-oxidizing system: Affinity for methanol, ethanol, propanol, and butanol. J. Biol. Chem. 1975, 250, 7397–7404. [Google Scholar] [PubMed]
- Caro, A.A.; Cederbaum, A.I. Oxidative stress, toxicology, and pharmacology of CYP2E1. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Moreno, F.; Petrides, A.S. Hepatic microsomal ethanol oxidizing system (MEOS): Respective roles of ethanol and carbohydrates for the enhanced activity after chronic alcohol consumption. Biochem. Pharmacol. 1981, 30, 1745–1751. [Google Scholar] [CrossRef]
- Lieber, C.S. Cytochrome P-4502E1: Its physiological and pathological role. Physiol. Rev. 1997, 77, 517–544. [Google Scholar] [PubMed]
- Dey, A. Cytochrome P450 2E1: Its clinical aspects and a brief perspective on the current research scenario. Subcell. Biochem. 2013, 67, 1–104. [Google Scholar] [PubMed]
- Grasselli, E.; Compalati, A.D.; Voci, A.; Vecchione, G.; Ragazzoni, M.; Gallo, G.; Borro, P.; Sumberaz, A.; Testino, G.; Vergani, L. Altered oxidative stress/antioxidant status in blood of alcoholic subjects is associated with alcoholic liver disease. Drug Alcohol Depend. 2014, 143, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.D.; Padmavathi, P.; Varadacharyulu, N.C. Emblica officinalis protects against alcohol-induced liver mitochondrial dysfunction in rats. J. Med. Food 2009, 12, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, Y.; Zhang, Y.J.; Zhou, Y.; Li, S.; Li, H.B. Natural products for the prevention and treatment of hangover and alcohol use disorder. Molecules 2016. [Google Scholar] [CrossRef] [PubMed]
- Maninang, J.S.; Lizada, M.C.C.; Gemma, H. Inhibition of aldehyde dehydrogenase enzyme by Durian (Durio zibethinus Murray) fruit extract. Food Chem. 2009, 117, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Wang, F.; Zhou, Y.; Li, Y.; Zhou, T.; Zheng, J.; Zhang, J.J.; Li, S.; Xu, D.P.; Li, H.B. Effects of 20 selected fruits on ethanol metabolism: Potential health benefits and harmful impacts. Int. J. Environ. Res. Public Health 2016. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.M.; Yusof, H.M.; Long, K.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Koh, S.P.; Abdullah, M.P.; Alitheen, N.B. Antioxidant and hepatoprotective effect of aqueous extract of germinated and fermented mung bean on ethanol-mediated liver damage. BioMed Res. Int. 2013. [Google Scholar] [CrossRef]
- Wang, J.M.; Zhang, Y.Y.; Zhang, Y.S.; Cui, Y.; Liu, J.; Zhang, B.F. Protective effect of Lysimachia christinae against acute alcohol-induced liver injury in mice. Biosci. Trends 2012, 6, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Dahiru, D.; Obidoa, O. Evaluation of the antioxidant effects of Ziziphus mauritiana Lam. leaf extracts against chronic ethanol-induced hepatotoxicity in rat liver. Afr. J. Tradit. Complement. 2008, 5, 39–45. [Google Scholar] [CrossRef]
- Yi, J.; Xia, W.; Wu, J.; Yuan, L.; Wu, J.; Tu, D.; Fang, J.; Tan, Z. Betulinic acid prevents alcohol-induced liver damage by improving the antioxidant system in mice. J. Vet. Sci. 2014, 15, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Giriwono, P.E.; Hashimoto, T.; Ohsaki, Y.; Shirakawa, H.; Hokazono, H.; Komai, M. Extract of fermented barley attenuates chronic alcohol induced liver damage by increasing antioxidative activities. Food Res. Int. 2010, 43, 118–124. [Google Scholar] [CrossRef]
- Huang, C.H.; Chang, Y.Y.; Liu, C.W.; Kang, W.Y.; Lin, Y.L.; Chang, H.C.; Chen, Y.C. Fruiting body of Niuchangchih (Antrodia camphorata) protects livers against chronic alcohol consumption damage. J. Agric. Food Chem. 2010, 58, 3859–3866. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Du, B.; Wang, Y.; Gao, H.; Cao, W.; Zheng, J.; Feng, F. Antioxidant properties of jujube honey and its protective effects against chronic alcohol-induced liver damage in mice. Food Funct. 2014, 5, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Masalkar, P.D.; Abhang, S.A. Oxidative stress and antioxidant status in patients with alcoholic liver disease. Clin. Chim. Acta 2005, 355, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ye, Y.; Kang, J.; Yao, X.; Zhang, Y.; Jiang, W.; Gao, M.; Dai, Y.; Xin, Y.; Wang, Q.; et al. l-Theanine prevents alcoholic liver injury through enhancing the antioxidant capability of hepatocytes. Food Chem. Toxicol. 2012, 50, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, K.; Swaminathan, K.; Kumar, S.M.; Chatterjee, S.; Clemens, D.L.; Dey, A. Elevated glutathione level does not protect against chronic alcohol mediated apoptosis in recombinant human hepatoma cell line VL-17A over-expressing alcohol metabolizing enzymes—Alcohol dehydrogenase and Cytochrome P450 2E1. Toxicol. in Vitro 2011, 25, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Ignatowicz, E.; Wozniak, A.; Kulza, M.; Senczuk-Przybylowska, M.; Cimino, F.; Piekoszewski, W.; Chuchracki, M.; Florek, E. Exposure to alcohol and tobacco smoke causes oxidative stress in rats. Pharmacol. Rep. 2013, 65, 906–913. [Google Scholar] [CrossRef]
- Das, S.K.; Dhanya, L.; Varadhan, S.; Mukherjee, S.; Vasudevan, D.M. Effects of chronic ethanol consumption in blood: A time dependent study on rat. Indian J. Clin. Biochem. 2009, 24, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.J.; Koh, E.J.; Kim, C.S.; Zee, O.P.; Kwak, J.H.; Jeong, W.J.; Kim, J.H.; Lee, S.M. Agrimonia eupatoria protects against chronic ethanol-induced liver injury in rats. Food Chem. Toxicol. 2012, 50, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xie, G.X.; Zhang, W.L.; Zhong, W.; Sun, X.H.; Tan, X.B.; Sun, X.G.; Jia, W.; Zhou, Z.X. Dietary nicotinic acid supplementation ameliorates chronic alcohol-induced fatty liver in rats. Alcohol Clin. Exp. Res. 2014, 38, 1982–1992. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, N.; Yamashina, S.; Kono, H.; Schemmer, P.; Rivera, C.A.; Enomoto, A.; Nishiura, T.; Nishimura, T.; Brenner, D.A.; Thurman, R.G. Development of a new, simple rat model of early alcohol-induced liver injury based on sensitization of Kupffer cells. Hepatology 1999, 29, 1680–1689. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Ye, Q.; Wang, H.Y.; Li, Y.C.; Xia, X.H.; Yao, W.R.; Qian, H. Aloin protects against chronic alcoholic liver injury via attenuating lipid accumulation, oxidative stress and inflammation in mice. Arch. Pharm. Res. 2014, 37, 1624–1633. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.E.; Dong, W.G.; Wang, B.Y.; Tong, Q.Y.; Li, Z.Y. Curcumin attenuates chronic ethanol-induced liver injury by inhibition of oxidative stress via mitogen-activated protein kinase/nuclear factor E2-related factor 2 pathway in mice. Pharmacogn. Mag. 2015, 11, 707–715. [Google Scholar] [PubMed]
- Xiang, J.L.; Zhu, W.X.; Li, Z.X.; Ling, S.B. Effect of juice and fermented vinegar from Hovenia dulcis peduncles on chronically alcohol-induced liver damage in mice. Food Funct. 2012, 3, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Shati, A.A.; Elsaid, F.G. Effects of water extracts of thyme (Thymus vulgaris) and ginger (Zingiber officinale Roscoe) on alcohol abuse. Food Chem. Toxicol. 2009, 47, 1945–1949. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.N.; Mu, T.H.; Liu, X.L.; Zhang, M.; Chen, J.W. Purple sweet potato (Ipomoea batatas L.) anthocyanins: Preventive effect on acute and subacute alcoholic liver damage and dealcoholic effect. J. Agric. Food Chem. 2014, 62, 2364–2373. [Google Scholar] [CrossRef] [PubMed]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Bertola, A.; Mathews, S.; Ki, S.H.; Wang, H.; Gao, B. Mouse model of chronic and binge ethanol feeding (the NIAAA model). Nat. Protoc. 2013, 8, 627–637. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.-J.; Zhou, T.; Wang, F.; Zhou, Y.; Li, Y.; Zhang, J.-J.; Zheng, J.; Xu, D.-P.; Li, H.-B. The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on Alcohol-Induced Liver Injury. Int. J. Mol. Sci. 2016, 17, 1616. https://doi.org/10.3390/ijms17101616
Zhang Y-J, Zhou T, Wang F, Zhou Y, Li Y, Zhang J-J, Zheng J, Xu D-P, Li H-B. The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on Alcohol-Induced Liver Injury. International Journal of Molecular Sciences. 2016; 17(10):1616. https://doi.org/10.3390/ijms17101616
Chicago/Turabian StyleZhang, Yu-Jie, Tong Zhou, Fang Wang, Yue Zhou, Ya Li, Jiao-Jiao Zhang, Jie Zheng, Dong-Ping Xu, and Hua-Bin Li. 2016. "The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on Alcohol-Induced Liver Injury" International Journal of Molecular Sciences 17, no. 10: 1616. https://doi.org/10.3390/ijms17101616
APA StyleZhang, Y. -J., Zhou, T., Wang, F., Zhou, Y., Li, Y., Zhang, J. -J., Zheng, J., Xu, D. -P., & Li, H. -B. (2016). The Effects of Syzygium samarangense, Passiflora edulis and Solanum muricatum on Alcohol-Induced Liver Injury. International Journal of Molecular Sciences, 17(10), 1616. https://doi.org/10.3390/ijms17101616