
Interactions between Two Different G Protein-Coupled Receptors in Reproductive Hormone-Producing Cells: The Role of PACAP and Its Receptor PAC1R

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) and Its Receptor
3. Hypothalamic Kisspeptin Controls GnRH Neurons
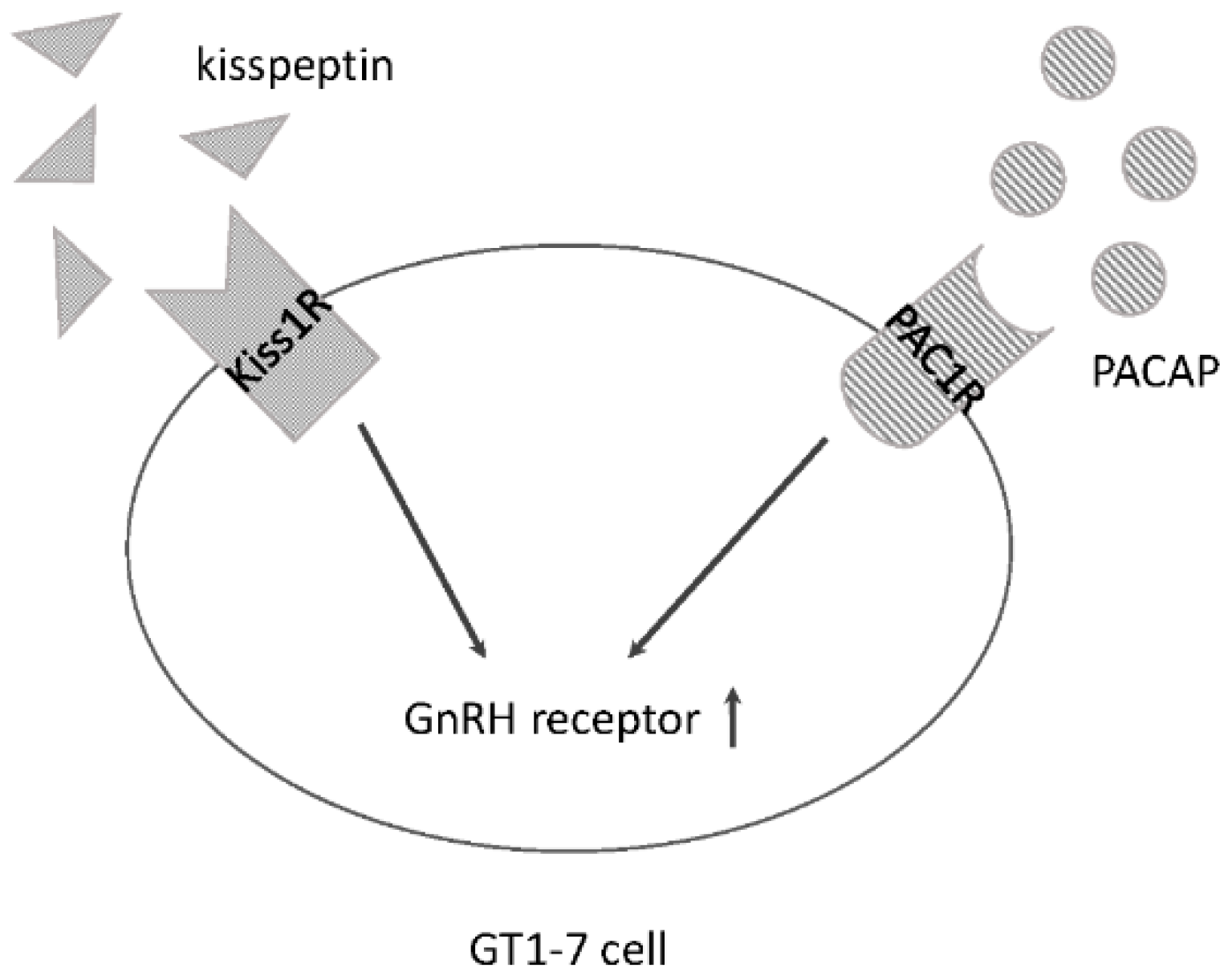
3.1. Kiss1R and PACAP within GnRH Neurons
3.2. Kisspeptin and PACAP Cooperate in GnRH-Producing Neurons
4. Gonadotropin Secretion from Pituitary Gonadotrophs
4.1. GnRHR Signalling in Pituitary Gonadotrophs
4.2. Effect of PACAP on Pituitary Gonadotrophs
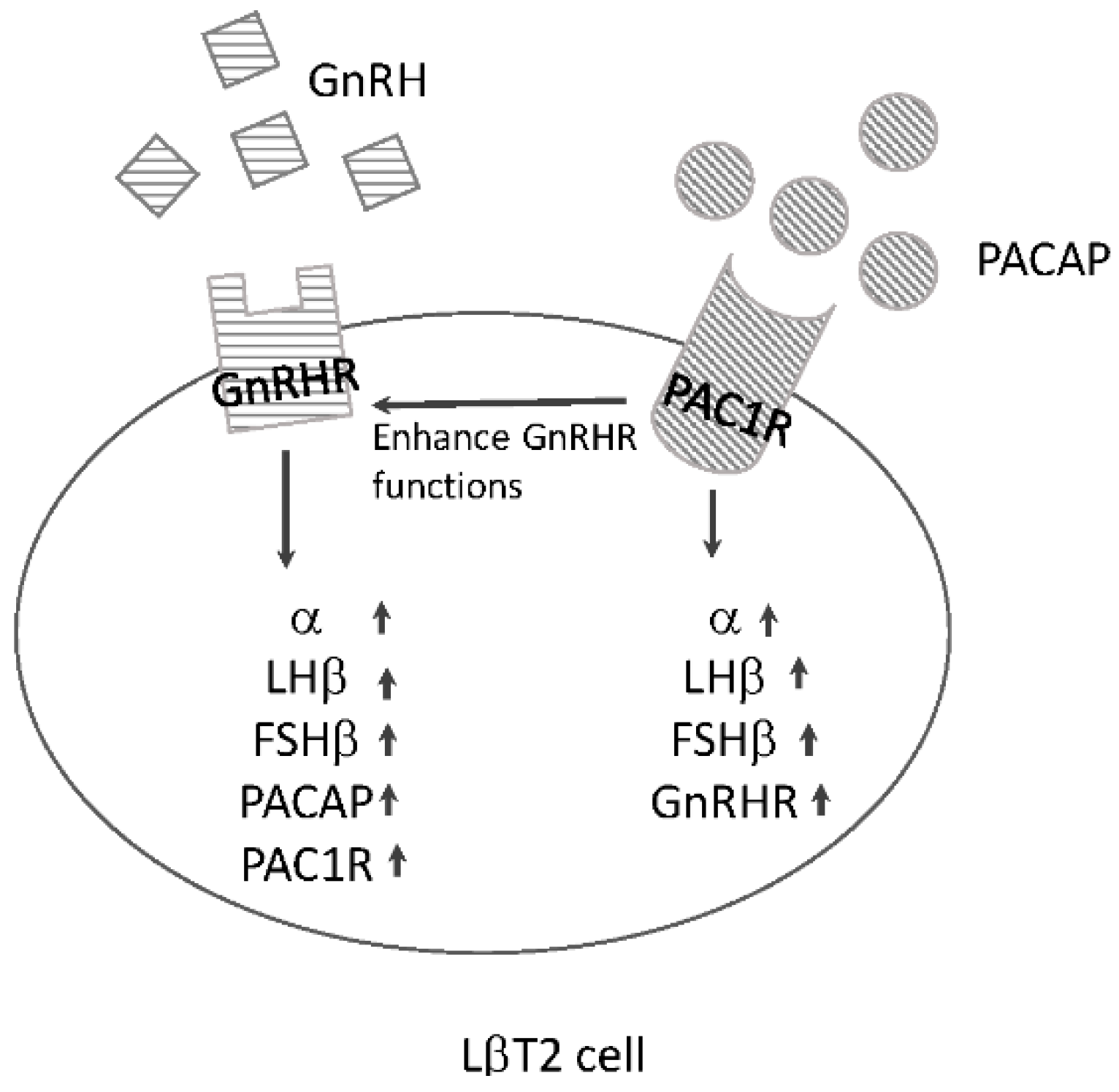
4.3. Interaction between GnRHR and PAC1R in Pituitary Gonadotrophs
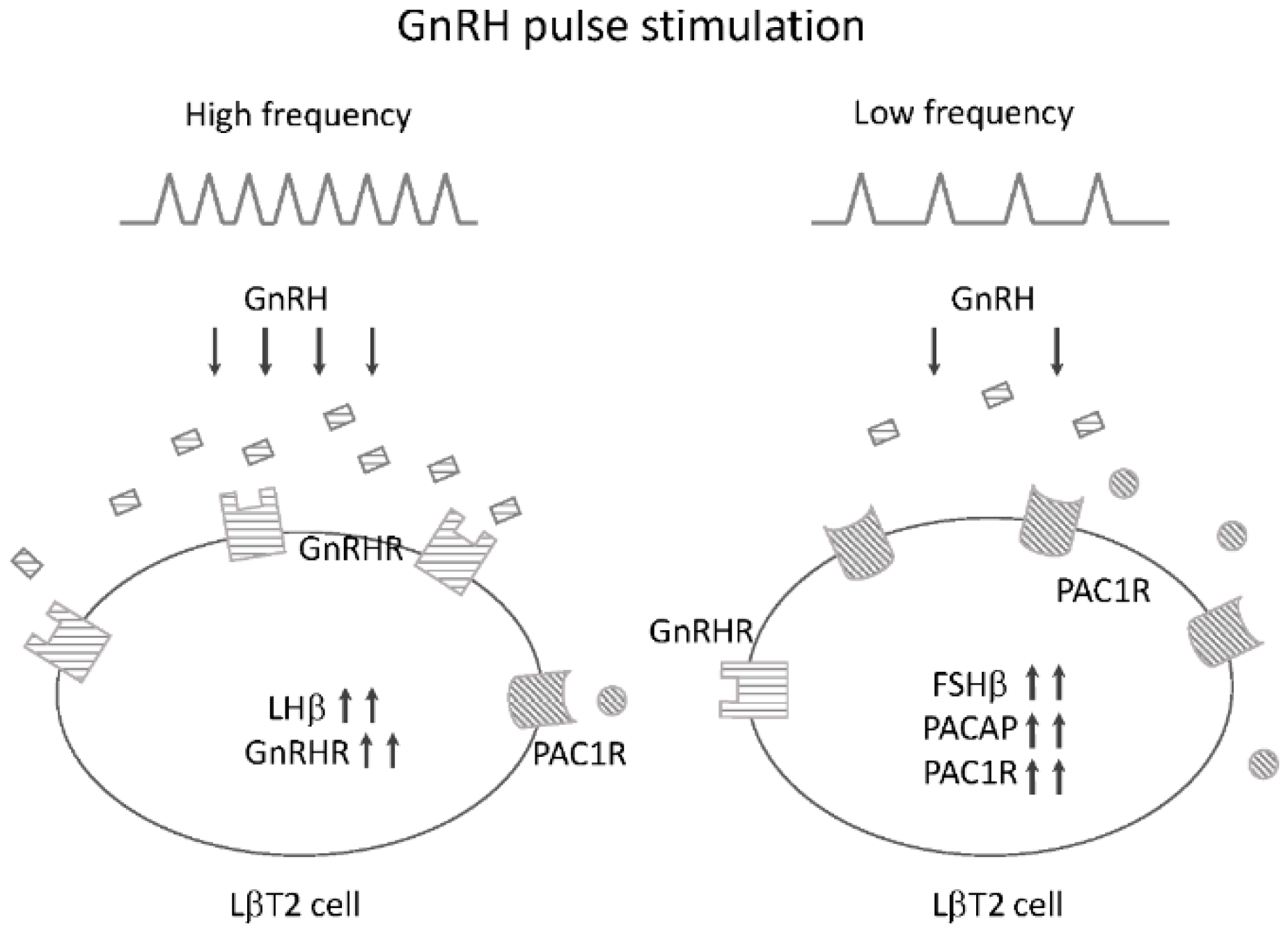
4.4. Potential Roles of PACAP and PAC1R in GnRH Pulse Frequency-Dependent Gonadotropin Subunit Expression
5. Prolactin Synthesis and Secretion from Pituitary Lactotrophs
5.1. Thyrotropin-Releasing Hormone (TRH) as a Prolactin-Releasing Factor
5.2. Effect of PACAP on Prolactin-Producing Cells
5.3. Interaction between the TRH Receptor and PAC1R in Somatolactotrophic GH3 Cells
6. Conclusions
Conflicts of Interest
References
- Gharib, S.D.; Wierman, M.E.; Shupnik, M.A.; Chin, W.W. Molecular biology of the pituitary gonadotropins. Endocr. Rev. 1990, 11, 177–199. [Google Scholar] [CrossRef] [PubMed]
- De Roux, N.; Genin, E.; Carel, J.C.; Matsuda, F.; Chaussain, J.L.; Milgrom, E. Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc. Natl. Acad. Sci. USA 2003, 100, 10972–10976. [Google Scholar] [CrossRef] [PubMed]
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S., Jr.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G.; et al. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef] [PubMed]
- Beltramo, M.; Dardente, H.; Cayla, X.; Caraty, A. Cellular mechanisms and integrative timing of neuroendocrine control of GnRH secretion by kisspeptin. Mol. Cell. Endocrinol. 2014, 382, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Maggi, R.; Cariboni, A.M.; Marelli, M.M.; Moretti, R.M.; Andre, V.; Marzagalli, M.; Limonta, P. GnRH and GnRH receptors in the pathophysiology of the human female reproductive system. Hum. Reprod. Update 2016, 22. [Google Scholar] [CrossRef] [PubMed]
- Crowley, W.F., Jr.; Filicori, M.; Spratt, D.I.; Santoro, N.F. The physiology of gonadotropin-releasing hormone (GnRH) secretion in men and women. Recent Prog. Horm. Res. 1985, 41, 473–531. [Google Scholar] [PubMed]
- Grattan, D.R.; Kokay, I.C. Prolactin: A pleiotropic neuroendocrine hormone. J. Neuroendocrinol. 2008, 20, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Hnasko, R. Dopamine as a prolactin (PRL) inhibitor. Endocr. Rev. 2001, 22, 724–763. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, H.; Oride, A.; Kyo, S. Role of pituitary adenylate cyclase-activating polypeptide in modulating hypothalamus-pituitary neuroendocrine functions in mouse cell models. J. Neuroendocrinol. 2015, 27, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Sherwood, N.M.; Krueckl, S.L.; McRory, J.E. The origin and function of the pituitary adenylate cyclase-activating polypeptide (PACAP)/glucagon superfamily. Endocr. Rev. 2000, 21, 619–670. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, J.; Mikkelsen, J.D.; Clausen, H.; Holst, J.J.; Wulff, B.S.; Fahrenkrug, J. Gene-expression of pituitary adenylate-cyclase activating polypeptide (PACAP) in the rat hypothalamus. Regul. Pept. 1995, 55, 133–148. [Google Scholar] [CrossRef]
- Kimura, S.; Ohshige, Y.; Lin, L.; Okumura, T.; Yanaihara, C.; Yanaihara, N.; Shiotani, Y. Localization of pituitary adenylate cyclase-activating polypeptide (PACAP) in the hypothalamus-pituitary system in rats: Light and electron microscopic immunocytochemical studies. J. Neuroendocrinol. 1994, 6, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Piggins, H.D.; Stamp, J.A.; Burns, J.; Rusak, B.; Semba, K. Distribution of pituitary adenylate cyclase activating polypeptide (PACAP) immunoreactivity in the hypothalamus and extended amygdala of the rat. J. Comp. Neurol. 1996, 376, 278–294. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Nogi, H.; Mori, K.; Ohishi, H.; Shigemoto, R.; Yamamoto, K.; Matsuda, T.; Mizuno, N.; Nagata, S.; Baba, A. Distribution of the mRNA for a pituitary adenylate cyclase-activating polypeptide receptor in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1996, 371, 567–577. [Google Scholar] [CrossRef]
- Vigh, S.; Arimura, A.; Gottschall, P.E.; Kitada, C.; Somogyvari-Vigh, A.; Childs, G.V. Cytochemical characterization of anterior pituitary target cells for the neuropeptide, pituitary adenylate cyclase activating polypeptide (PACAP), using biotinylated ligands. Peptides 1993, 14, 59–65. [Google Scholar] [CrossRef]
- Piet, R.; de Croft, S.; Liu, X.; Herbison, A.E. Electrical properties of kisspeptin neurons and their regulation of GnRH neurons. Front. Neuroendocrinol. 2015, 36, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Han, S.K.; Gottsch, M.L.; Lee, K.J.; Popa, S.M.; Smith, J.T.; Jakawich, S.K.; Clifton, D.K.; Steiner, R.A.; Herbison, A.E. Activation of gonadotropin-releasing hormone neurons by kisspeptin as a neuroendocrine switch for the onset of puberty. J. Neurosci. 2005, 25, 11349–11356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Roepke, T.A.; Kelly, M.J.; Ronnekleiv, O.K. Kisspeptin depolarizes gonadotropin-releasing hormone neurons through activation of TRPC-like cationic channels. J. Neurosci. 2008, 28, 4423–4434. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.L.; Patterson, M.; Murphy, K.G.; Smith, K.L.; Dhillo, W.S.; Todd, J.F.; Ghatei, M.A.; Bloom, S.R. Central and peripheral administration of kisspeptin-10 stimulates the hypothalamic-pituitary-gonadal axis. J. Neuroendocrinol. 2004, 16, 850–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novaira, H.J.; Ng, Y.; Wolfe, A.; Radovick, S. Kisspeptin increases GnRH mRNA expression and secretion in GnRH secreting neuronal cell lines. Mol. Cell. Endocrinol. 2009, 311, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Skorupskaite, K.; George, J.T.; Anderson, R.A. The kisspeptin-GnRH pathway in human reproductive health and disease. Hum. Reprod. Update 2014, 20, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Messager, S.; Chatzidaki, E.E.; Ma, D.; Hendrick, A.G.; Zahn, D.; Dixon, J.; Thresher, R.R.; Malinge, I.; Lomet, D.; Carlton, M.B.; et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc. Natl. Acad. Sci. USA 2005, 102, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.M.; le Poul, E.; Brezillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [PubMed]
- Ohtaki, T.; Shintani, Y.; Honda, S.; Matsumoto, H.; Hori, A.; Kanehashi, K.; Terao, Y.; Kumano, S.; Takatsu, Y.; Masuda, Y.; et al. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001, 411, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Stafford, L.J.; Xia, C.; Ma, W.; Cai, Y.; Liu, M. Identification and characterization of mouse metastasis-suppressor KiSS1 and its G-protein-coupled receptor. Cancer Res. 2002, 62, 5399–5404. [Google Scholar] [PubMed]
- Sukhbaatar, U.; Kanasaki, H.; Mijiddorj, T.; Oride, A.; Miyazaki, K. Kisspeptin induces expression of gonadotropin-releasing hormone receptor in GnRH-producing GT1–7 cells overexpressing G protein-coupled receptor 54. Gen. Comp. Endocrinol. 2013, 194, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Castellano, J.M.; Navarro, V.M.; Fernandez-Fernandez, R.; Castano, J.P.; Malagon, M.M.; Aguilar, E.; Dieguez, C.; Magni, P.; Pinilla, L.; Tena-Sempere, M. Ontogeny and mechanisms of action for the stimulatory effect of kisspeptin on gonadotropin-releasing hormone system of the rat. Mol. Cell. Endocrinol. 2006, 257–258, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, M.; Alcin, E.; Ayar, A.; Yilmaz, B.; Sandal, S.; Kelestimur, H. Kisspeptin-10 elicits triphasic cytosolic calcium responses in immortalized GT1–7 GnRH neurones. Neurosci. Lett. 2011, 492, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.T.; Sawangjaroen, K.; Curlewis, J.D. Pituitary adenylate cyclase-activating polypeptide acts within the medial basal hypothalamus to inhibit prolactin and luteinizing hormone secretion. Endocrinology 1996, 137, 3424–3429. [Google Scholar] [PubMed]
- Szabo, F.; Horvath, J.; Heinzlmann, A.; Arimura, A.; Koves, K. Neonatal PACAP administration in rats delays puberty through the influence of the LHRH neuronal system. Regul. Pept. 2002, 109, 49–55. [Google Scholar] [CrossRef]
- Li, S.; Grinevich, V.; Fournier, A.; Pelletier, G. Effects of pituitary adenylate cyclase-activating polypeptide (PACAP) on gonadotropin-releasing hormone and somatostatin gene expression in the rat brain. Brain Res. Mol. Brain Res. 1996, 41, 157–162. [Google Scholar] [CrossRef]
- Kanasaki, H.; Mijiddorj, T.; Sukhbaatar, U.; Oride, A.; Miyazaki, K. Pituitary adenylate cyclase-activating polypeptide (PACAP) increases expression of the gonadotropin-releasing hormone (GnRH) receptor in GnRH-producing GT1–7 cells overexpressing PACAP type I receptor. Gen. Comp. Endocrinol. 2013, 193, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Wildt, L.; Hausler, A.; Marshall, G.; Hutchison, J.S.; Plant, T.M.; Belchetz, P.E.; Knobil, E. Frequency and amplitude of gonadotropin-releasing hormone stimulation and gonadotropin secretion in the rhesus monkey. Endocrinology 1981, 109, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Conn, P.M.; Huckle, W.R.; Andrews, W.V.; McArdle, C.A. The molecular mechanism of action of gonadotropin releasing hormone (GnRH) in the pituitary. Recent Prog. Horm. Res. 1987, 43, 29–68. [Google Scholar] [PubMed]
- Sundaresan, S.; Colin, I.M.; Pestell, R.G.; Jameson, J.L. Stimulation of mitogen-activated protein kinase by gonadotropin-releasing hormone: Evidence for the involvement of protein kinase C. Endocrinology 1996, 137, 304–311. [Google Scholar] [PubMed]
- Kraus, S.; Naor, Z.; Seger, R. Intracellular signaling pathways mediated by the gonadotropin-releasing hormone (GnRH) receptor. Arch. Med. Res. 2001, 32, 499–509. [Google Scholar] [CrossRef]
- Liu, F.; Usui, I.; Evans, L.G.; Austin, D.A.; Mellon, P.L.; Olefsky, J.M.; Webster, N.J. Involvement of both G(q/11) and G(s) proteins in gonadotropin-releasing hormone receptor-mediated signaling in LβT2 cells. J. Biol. Chem. 2002, 277, 32099–32108. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.; Chuderland, D.; Bonfil, D.; Kraus, S.; Seger, R.; Naor, Z. Extracellular signal-regulated kinase and c-Src, but not Jun N-terminal kinase, are involved in basal and gonadotropin-releasing hormone-stimulated activity of the glycoprotein hormone α-subunit promoter. Endocrinology 2003, 144, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.; Bonfil, D.; Chuderland, D.; Kraus, S.; Seger, R.; Naor, Z. Activation of MAPK cascades by GnRH: ERK and Jun N-terminal kinase are involved in basal and GnRH-stimulated activity of the glycoprotein hormone LHβ-subunit promoter. Endocrinology 2002, 143, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Vasilyev, V.V.; Pernasetti, F.; Rosenberg, S.B.; Barsoum, M.J.; Austin, D.A.; Webster, N.J.; Mellon, P.L. Transcriptional activation of the ovine follicle-stimulating hormone-β gene by gonadotropin-releasing hormone involves multiple signal transduction pathways. Endocrinology 2002, 143, 1651–1659. [Google Scholar] [PubMed]
- Kanasaki, H.; Bedecarrats, G.Y.; Kam, K.Y.; Xu, S.; Kaiser, U.B. Gonadotropin-releasing hormone pulse frequency-dependent activation of extracellular signal-regulated kinase pathways in perifused LβT2 cells. Endocrinology 2005, 146, 5503–5513. [Google Scholar] [CrossRef] [PubMed]
- Keyse, S.M. Protein phosphatases and the regulation of MAP kinase activity. Semin. Cell Dev. Biol. 1998, 9, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Purwana, I.N.; Kanasaki, H.; Mijiddorj, T.; Oride, A.; Miyazaki, K. Induction of dual-specificity phosphatase 1 (DUSP1) by pulsatile gonadotropin-releasing hormone stimulation: Role for gonadotropin subunit expression in mouse pituitary LβT2 cells. Biol. Reprod. 2011, 84, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Koves, K.; Kantor, O.; Scammell, J.G.; Arimura, A. PACAP colocalizes with luteinizing and follicle-stimulating hormone immunoreactivities in the anterior lobe of the pituitary gland. Peptides 1998, 19, 1069–1072. [Google Scholar] [CrossRef]
- Jin, L.; Tsumanuma, I.; Ruebel, K.H.; Bayliss, J.M.; Lloyd, R.V. Analysis of homogeneous populations of anterior pituitary folliculostellate cells by laser capture microdissection and reverse transcription-polymerase chain reaction. Endocrinology 2001, 142, 1703–1709. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Kanasaki, H.; Mutiara, S.; Oride, A.; Miyazaki, K. Cyclic adenosine 3′,5′-monophosphate/protein kinase A and mitogen-activated protein kinase 3/1 pathways are involved in adenylate cyclase-activating polypeptide 1-induced common α-glycoprotein subunit gene (Cga) expression in mouse pituitary gonadotroph LβT2 cells. Biol. Reprod. 2007, 77, 707–716. [Google Scholar] [PubMed]
- Kanasaki, H.; Mutiara, S.; Oride, A.; Purwana, I.N.; Miyazaki, K. Pulse frequency-dependent gonadotropin gene expression by adenylate cyclase-activating polypeptide 1 in perifused mouse pituitary gonadotroph LβT2 cells. Biol. Reprod. 2009, 81, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Grafer, C.M.; Thomas, R.; Lambrakos, L.; Montoya, I.; White, S.; Halvorson, L.M. GnRH stimulates expression of PACAP in the pituitary gonadotropes via both the PKA and PKC signaling systems. Mol. Endocrinol. 2009, 23, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Purwana, I.N.; Kanasaki, H.; Oride, A.; Mijiddorj, T.; Shintani, N.; Hashimoto, H.; Baba, A.; Miyazaki, K. GnRH-induced PACAP and PAC1 receptor expression in pituitary gonadotrophs: A possible role in the regulation of gonadotropin subunit gene expression. Peptides 2010, 31, 1748–1755. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, H.; Purwana, I.N.; Mijiddorj, T.; Oride, A.; Miyazaki, K. Possible involvement of PACAP and PACAP type 1 receptor in GnRH-induced FSH β-subunit gene expression. Regul. Pept. 2011, 167, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, U.B.; Sabbagh, E.; Katzenellenbogen, R.A.; Conn, P.M.; Chin, W.W. A mechanism for the differential regulation of gonadotropin subunit gene expression by gonadotropin-releasing hormone. Proc. Natl. Acad. Sci. USA 1995, 92, 12280–12284. [Google Scholar] [CrossRef] [PubMed]
- Purwana, I.N.; Kanasaki, H.; Oride, A.; Mijiddorj, T.; Miyazaki, K. Expression of the pituitary adenylate cyclase-activating polypeptide (PACAP) type 1 receptor (PAC1R) potentiates the effects of GnRH on gonadotropin subunit gene expression. Biochem. Biophys. Res. Commun. 2011, 410, 295–300. [Google Scholar] [CrossRef] [PubMed]
- McArdle, C.A.; Counis, R. GnRH and PACAP action in gonadotropes: Cross-talk between phosphoinositidase C and adenylyl cyclase mediated signaling pathways. Trends Endocrinol. Metab. 1996, 7, 168–175. [Google Scholar] [CrossRef]
- Lariviere, S.; Garrel-Lazayres, G.; Simon, V.; Shintani, N.; Baba, A.; Counis, R.; Cohen-Tannoudji, J. Gonadotropin-releasing hormone inhibits pituitary adenylyl cyclase-activating polypeptide coupling to 3′,5′-cyclic adenosine-5′-monophosphate pathway in LβT2 gonadotrope cells through novel protein kinase C isoforms and phosphorylation of pituitary adenylyl cyclase-activating polypeptide type I receptor. Endocrinology 2008, 149, 6389–6398. [Google Scholar] [PubMed]
- Kaiser, U.B.; Conn, P.M.; Chin, W.W. Studies of gonadotropin-releasing hormone (GnRH) action using GnRH receptor-expressing pituitary cell lines. Endocr. Rev. 1997, 18, 46–70. [Google Scholar] [CrossRef] [PubMed]
- Hennighausen, L.; Robinson, G.W. Signaling pathways in mammary gland development. Dev. Cell 2001, 1, 467–475. [Google Scholar] [CrossRef]
- Ormandy, C.J.; Camus, A.; Barra, J.; Damotte, D.; Lucas, B.; Buteau, H.; Edery, M.; Brousse, N.; Babinet, C.; Binart, N.; et al. Null mutation of the prolactin receptor gene produces multiple reproductive defects in the mouse. Genes Dev. 1997, 11, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Kauppila, A.; Martikainen, H.; Puistola, U.; Reinila, M.; Ronnberg, L. Hypoprolactinemia and ovarian function. Fertil. Steril. 1988, 49, 437–441. [Google Scholar] [CrossRef]
- Melgar, V.; Espinosa, E.; Sosa, E.; Rangel, M.J.; Cuenca, D.; Ramirez, C.; Mercado, M. Current diagnosis and treatment of hyperprolactinemia. Rev. Med. Inst. Mex. Seguro Soc. 2016, 54, 111–121. [Google Scholar] [PubMed]
- Tashjian, A.H., Jr.; Barowsky, N.J.; Jensen, D.K. Thyrotropin releasing hormone: Direct evidence for stimulation of prolactin production by pituitary cells in culture. Biochem. Biophys. Res. Commun. 1971, 43, 516–523. [Google Scholar] [CrossRef]
- Jacobs, L.S.; Snyder, P.J.; Wilber, J.F.; Utiger, R.D.; Daughaday, W.H. Increased serum prolactin after administration of synthetic thyrotropin releasing hormone (TRH) in man. J. Clin. Endocrinol. Metab. 1971, 33, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Gershengorn, M.C. Mechanism of thyrotropin releasing hormone stimulation of pituitary hormone secretion. Annu. Rev. Physiol. 1986, 48, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Ohmichi, M.; Sawada, T.; Kanda, Y.; Koike, K.; Hirota, K.; Miyake, A.; Saltiel, A.R. Thyrotropin-releasing hormone stimulates MAP kinase activity in GH3 cells by divergent pathways. Evidence of a role for early tyrosine phosphorylation. J. Biol. Chem. 1994, 269, 3783–3788. [Google Scholar] [PubMed]
- Ryser, S.; Tortola, S.; van Haasteren, G.; Muda, M.; Li, S.; Schlegel, W. MAP kinase phosphatase-1 gene transcription in rat neuroendocrine cells is modulated by a calcium-sensitive block to elongation in the first exon. J. Biol. Chem. 2001, 276, 33319–33327. [Google Scholar] [CrossRef] [PubMed]
- Oride, A.; Kanasaki, H.; Mutiara, S.; Purwana, I.N.; Miyazaki, K. Activation of extracellular signal-regulated kinase (ERK) and induction of mitogen-activated protein kinase phosphatase 1 (MKP-1) by perifused thyrotropin-releasing hormone (TRH) stimulation in rat pituitary GH3 cells. Mol. Cell. Endocrinol. 2008, 296, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, H.; Fukunaga, K.; Takahashi, K.; Miyazaki, K.; Miyamoto, E. Mitogen-activated protein kinase activation by stimulation with thyrotropin-releasing hormone in rat pituitary GH3 cells. Biol. Reprod. 1999, 61, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, H.; Yonehara, T.; Yamamoto, H.; Takeuchi, Y.; Fukunaga, K.; Takahashi, K.; Miyazaki, K.; Miyamoto, E. Differential regulation of pituitary hormone secretion and gene expression by thyrotropin-releasing hormone: A role for mitogen-activated protein kinase signaling cascade in rat pituitary GH3 cells. Biol. Reprod. 2002, 67, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Sawangjaroen, K.; Anderson, S.T.; Curlewis, J.D. Effects of pituitary adenylate cyclase-activating polypeptide (PACAP) and vasoactive intestinal polypeptide (VIP) on hormone secretion from sheep pituitary cells in vitro. J. Neuroendocrinol. 1997, 9, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, T.; Soliman, E.B.; Kanematsu, S. Effects of pituitary adenylate cyclase-activating polypeptide (PACAP), prostaglandin E2 (PGE2) and growth hormone releasing factor (GRF) on the release of growth hormone from cultured bovine anterior pituitary cells in vitro. Domest. Anim. Endocrinol. 1994, 11, 331–337. [Google Scholar] [CrossRef]
- Arbogast, L.A.; Voogt, J.L. Pituitary adenylate cyclase-activating polypeptide (PACAP) increases prolactin release and tuberoinfundibular dopaminergic neuronal activity. Brain Res. 1994, 655, 17–24. [Google Scholar] [CrossRef]
- Benter, S.; Leonhardt, S.; Wuttke, W.; Jarry, H. Paracrine cell to cell interactions determine the effects of pituitary adenylate cyclase activating polypeptide (PACAP) on in vitro prolactin release from rat pituitary cells. Exp. Clin. Endocrinol. Diabetes 1995, 103, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Yonehara, T.; Kanasaki, H.; Yamamoto, H.; Fukunaga, K.; Miyazaki, K.; Miyamoto, E. Involvement of mitogen-activated protein kinase in cyclic adenosine 3′,5′-monophosphate-induced hormone gene expression in rat pituitary GH(3) cells. Endocrinology 2001, 142, 2811–2819. [Google Scholar] [PubMed]
- Mijiddorj, T.; Kanasaki, H.; Purwana, I.N.; Oride, A.; Miyazaki, K. Stimulatory effect of pituitary adenylate-cyclase activating polypeptide (PACAP) and its PACAP type I receptor (PAC1R) on prolactin synthesis in rat pituitary somatolactotroph GH3 cells. Mol. Cell. Endocrinol. 2011, 339, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Knobil, E. Patterns of hypophysiotropic signals and gonadotropin secretion in the rhesus monkey. Biol. Reprod. 1981, 24, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Hinkle, P.M.; Tashjian, A.H., Jr. Thyrotropin-releasing hormone regulates the number of its own receptors in the GH3 strain of pituitary cells in culture. Biochemistry 1975, 14, 3845–3851. [Google Scholar] [CrossRef] [PubMed]
- Mijiddorj, T.; Kanasaki, H.; Unurjargal, S.; Oride, A.; Purwana, I.; Miyazaki, K. Prolonged stimulation with thyrotropin-releasing hormone and pituitary adenylate cyclase-activating polypeptide desensitize their receptor functions in prolactin-producing GH3 cells. Mol. Cell. Endocrinol. 2013, 365, 139–145. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanasaki, H.; Oride, A.; Hara, T.; Mijiddorj, T.; Sukhbaatar, U.; Kyo, S. Interactions between Two Different G Protein-Coupled Receptors in Reproductive Hormone-Producing Cells: The Role of PACAP and Its Receptor PAC1R. Int. J. Mol. Sci. 2016, 17, 1635. https://doi.org/10.3390/ijms17101635
Kanasaki H, Oride A, Hara T, Mijiddorj T, Sukhbaatar U, Kyo S. Interactions between Two Different G Protein-Coupled Receptors in Reproductive Hormone-Producing Cells: The Role of PACAP and Its Receptor PAC1R. International Journal of Molecular Sciences. 2016; 17(10):1635. https://doi.org/10.3390/ijms17101635
Chicago/Turabian StyleKanasaki, Haruhiko, Aki Oride, Tomomi Hara, Tselmeg Mijiddorj, Unurjargal Sukhbaatar, and Satoru Kyo. 2016. "Interactions between Two Different G Protein-Coupled Receptors in Reproductive Hormone-Producing Cells: The Role of PACAP and Its Receptor PAC1R" International Journal of Molecular Sciences 17, no. 10: 1635. https://doi.org/10.3390/ijms17101635
APA StyleKanasaki, H., Oride, A., Hara, T., Mijiddorj, T., Sukhbaatar, U., & Kyo, S. (2016). Interactions between Two Different G Protein-Coupled Receptors in Reproductive Hormone-Producing Cells: The Role of PACAP and Its Receptor PAC1R. International Journal of Molecular Sciences, 17(10), 1635. https://doi.org/10.3390/ijms17101635